1. Introduction
With the rapid development of the modern economy, the demand for high-protein food sources is increasing year by year, and beef, mutton, and dairy products (especially ruminant products) seem to be the first choice for most consumers [
1,
2]. However, ruminant production is one of the main drivers of global environmental degradation, and the contribution to environmental pollution is much greater compared with non-ruminants [
3,
4]. Ruminants account for over 70% of global livestock ammonia (NH
3) emissions and around 30% of total anthropogenic NH
3 emissions, resulting in significant economic losses and negative human health impacts [
5,
6]. In China, 55–59% of total nitrogen (N) emissions in the past decade (2004–2014) have been emitted into the air in gaseous form, with agricultural and livestock production accounting for the largest amount (62–69%) [
7]. Hence, reducing the environmental impact of N emissions while meeting the growing demand for ruminant products is one of the main goals of current sustainable food production systems.
Feed nutrient composition is considered an important reference strategy to improve N retention and reduce N emission in ruminant production, e.g., using low-protein diets [
8] or adjusting the rumen degradable to non-degradable protein ratio with a non-soluble to soluble carbohydrates ratio in diets [
9]. Our previous study on
Hu sheep indicated that when reducing crude protein (CP) content as well as adjusting soluble protein (SP) to 25–30%, N efficiency was increased [
10]. Stefanski et al., isolated different protein fractions from rapeseed meal and demonstrated a different efficiency of ruminal N metabolism of these fractions in vivo [
11]. Undeniably, feed protein fractions play an important role in nutrient absorption of ruminants, specifically the efficiency of N utilization.
Current research on SP in ruminant production has been mainly focused on establishing models of rumen metabolism, typically fractions PA1 (non-protein nitrogen, NPN) and PA2 (true protein) described by the latest CNCPS version 6.5 [
12]. However, diet-microbiome interactions in the context of SP are rarely reported. Our previous research indicated that altering dietary SP levels to ~25–30% with decreasing CP increased the relative abundance of Prevotellaceae and
Prevotella_1 in the rumen [
10]. Ciliate protozoa accounting for an estimated 25% to 50% of microbial biomass in the rumen are also involved in the breakdown of nitrogenous substances, especially insoluble proteins [
13]. Despite the antagonism of predation in protozoa–bacteria interactions, positive interactions (e.g., metabolic exchange) have also been demonstrated [
14,
15]. For instance, ciliate protozoa convert a small amount of insoluble protein that cannot be used in the rumen into SP for bacterial growth and utilization, thus contributing to dietary protein degradation [
16]. Solomon et al., reported the existence of protozoa-mediated enrichment of the Gammaproteobacteria family and genera and increased the overall diversity of the genera
Prevotella and
Treponema, which resulted in significant individual variation in end-product metabolites including methane [
17].
Rumen bacteria–bacteria interactions have attracted considerable attention in recent years, and our previous work has investigated the relationship between rumen bacteria, metabolites and phenotype when feeding Hu sheep low-protein diets with different SP. However, there is a paucity of information on how bacteria–protozoa interactions play a role in the rumen degradation of SP. In this study, we aimed to evaluate rumen bacterial communities, protozoal communities, and bacteria–protozoa interactions, as well as their associations with N metabolism in response to different SP levels in vitro. We hypothesized that altering SP levels could regulate rumen bacterial and protozoal communities, which result in different efficiency of N metabolism in vitro.
4. Discussion
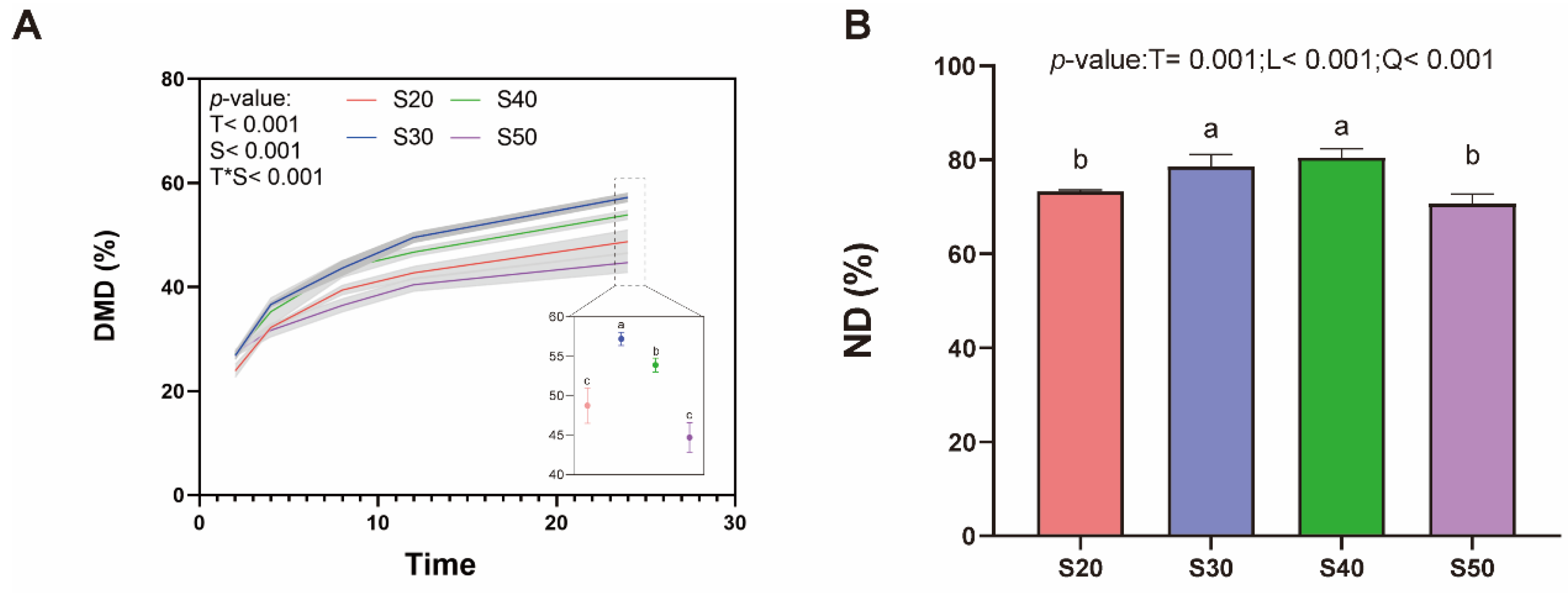
Regulation of dietary protein fractions is one of the important nutritional intervention strategies to improve nutritional metabolism in ruminants. Our previous in vivo experiments have shown that SP levels (% of CP) of ∼25–30% have higher N efficiency with dietary CP content decreased by∼10% in
Hu sheep [
10]. In this study, as time progressed, the pH value of each treatment gradually decreased and fluctuated from 6.85 to 5.90, which was mainly due to the decomposition and utilization of substrate nutrients by rumen microorganisms and the accumulation of organic acids [
30]. It is worth noting that the urea addition level of the substrate gradually increased (0~1.6% DM). Tian et al. [
31] showed that the hydrolysis of urea is alkaline; thus, with the increase in urea addition, the pH value also increased. However, the change in pH was not obvious among treatments in this study, which may be related to the urea addition levels.
The fluctuation of NH
3-N concentration in the rumen reflects the degradation of dietary N and the utilization of NH
3-N by rumen microorganisms. Generally, the optimal NH
3-N concentration of rumen microorganisms is 6.3–27.5 mg/dL [
32]. In our study, the NH
3-N concentration of each treatment fluctuated from 9.6 to 24.5 mg/dL, which was within the normal range. However, SP treatments significantly changed the NH
3-N concentration, especially in the S50, in which it was at a high level from 4 h of in vitro fermentation, which may be mainly related to the higher SP level (50% of CP) in the substrate. A large amount of SP in the substrate was rapidly decomposed in the rumen to generate ammonia, polypeptides and amino acids, while excess ammonia could not be utilized by rumen microorganisms [
33]. The above results were also consistent with the in vivo study. Similarly, Wilson et al. [
34] reported that the NH
3-N concentration in the rumen was significantly up-regulated with increasing dietary SP (% of CP) from 34.4% to 44.9%. At 24 h, as the substrate SP (% of CP) was 30, the NH
3-N concentration was lower, which may reflect the balance of substrate carbon and N degradation, and the rumen microorganisms could utilize the generated NH
3-N to a greater extent.
Stern et al. [
35] found that substrate protein solubility (22.7 vs. 36% of CP) had no significant effect on MCP synthesis in vitro, and high SP substrates (36% of CP) seemed to reduce the DM digestibility, which was similar to our results of decreased DM digestibility in S50, the reason might be that the release of substrate energy and N in S50 was asynchronous, which led to the decline of fermentation efficiency [
36]. Moreover, Hume et al. [
37] reported that the maximum synthesis of MCP was related to the NH
3-N concentration (8.8 mg/dL), which is comparable to the concentration in the S30 at 8 h (10.93 mg/dL), and the MCP content at this time point was indeed the highest numerically. In addition, our previous in vivo study also found that with the increase in SP levels (21.2–29.4%), no difference was found in MCP content.
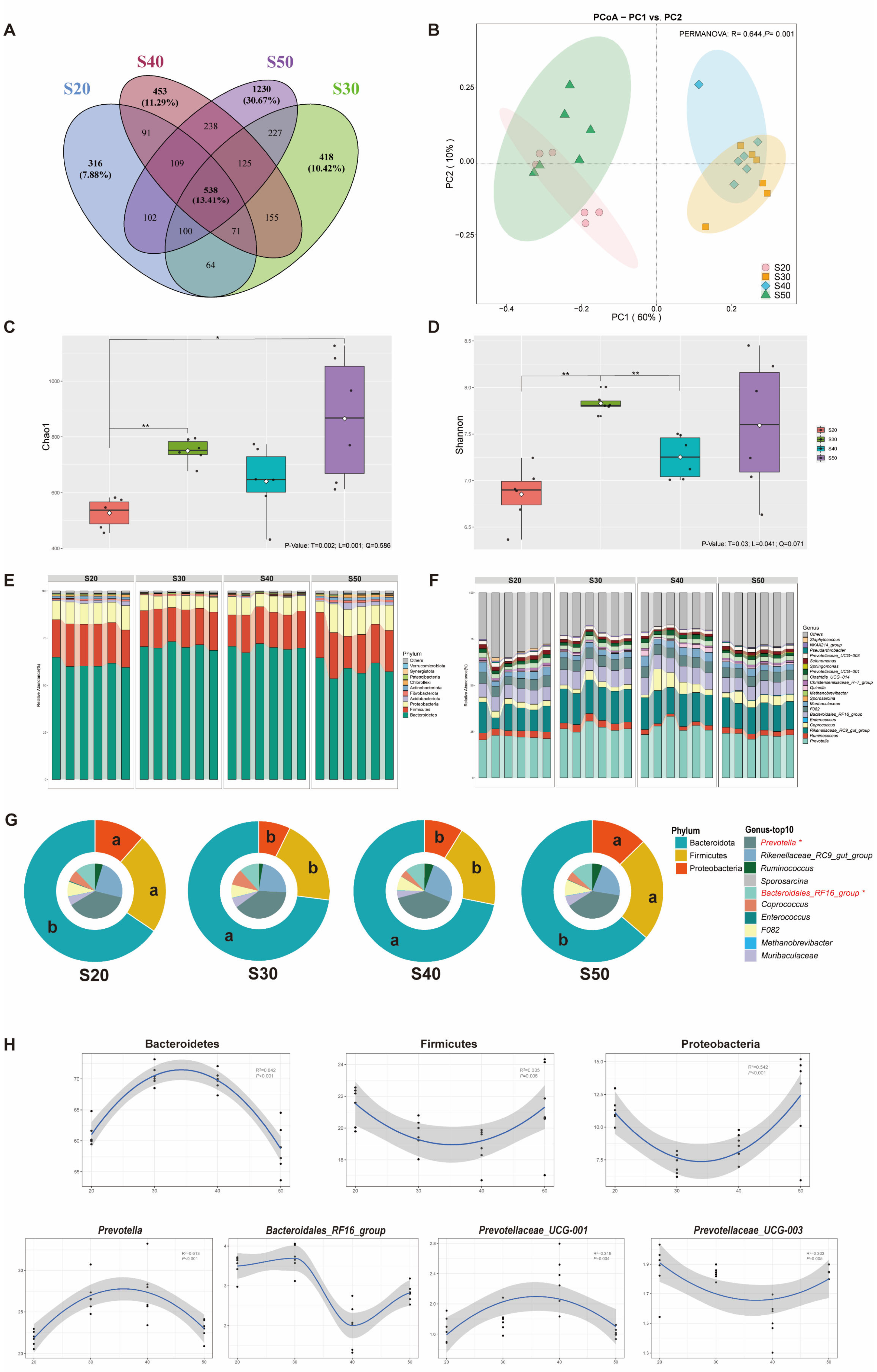
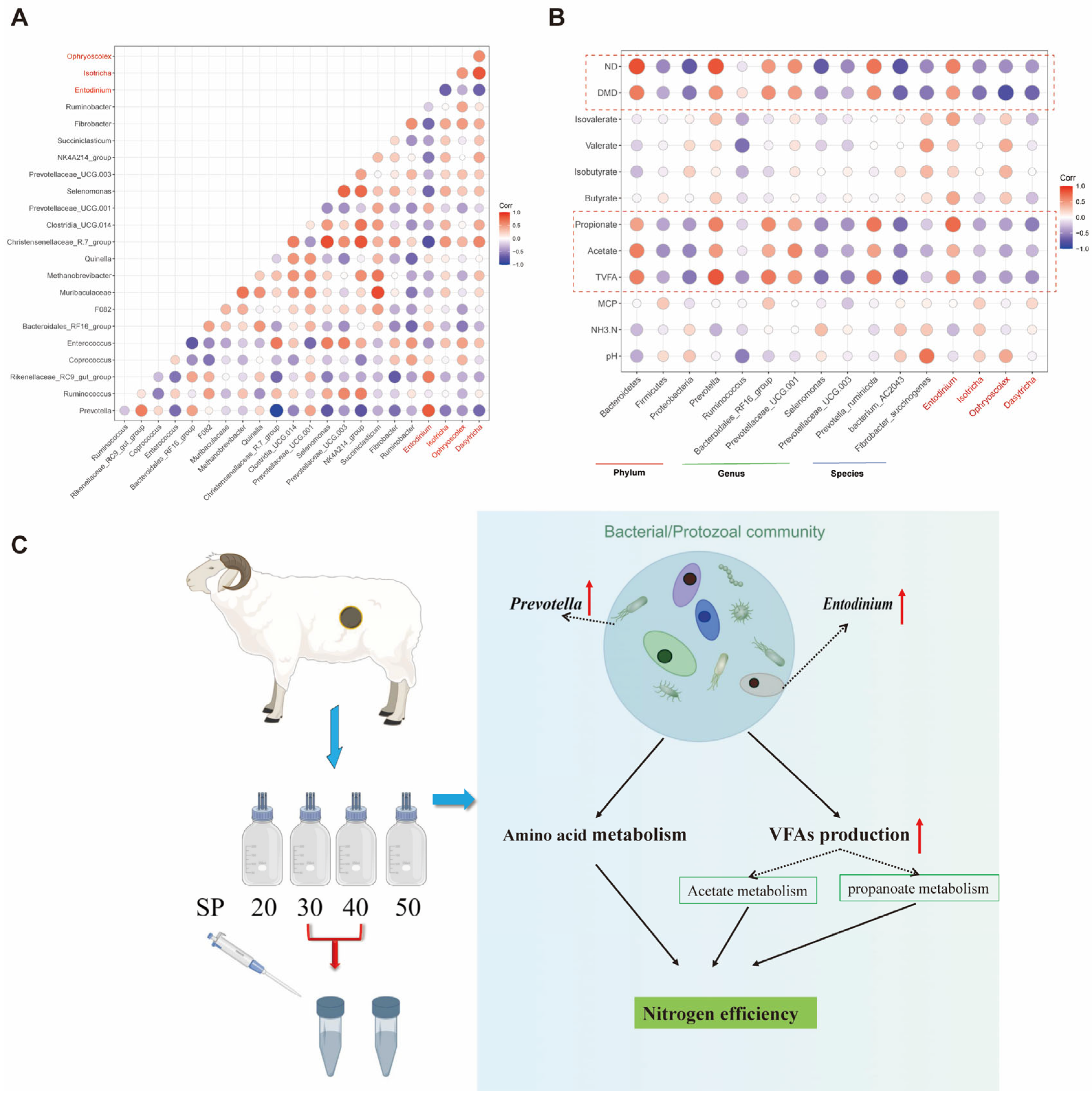
From the view of the rumen microbiome, the abundance of Bacteroidetes was improved, and Firmicutes and Proteobacteria were decreased as SP (% of CP) was 30 and 40. The total relative abundance of these three bacterial phyla in this study was over 80%. The host digests plant feed primarily through fermentation mediated by the microorganisms in these three phyla levels [
38]. In addition, VFA, acetate, propionate and valerate were also elevated with S30 and S40, which may be attributed to the enrichment of
Prevotella, which was the core genus of Bacteroidetes. The functions of this microbe include the fermentation of carbohydrates, the utilization of nitrogenous substances, and the biotransformation of bile acids and other steroids; the main by-products of anaerobic respiration are acetate, isovalerate, and succinate, which are closely related to VFA biosynthesis [
39,
40]. Moreover, propionate is a major substrate of gluconeogenesis, and a recent study also showed that it is closely related to ruminal N metabolism [
41]. Unsurprisingly, bacteria functional annotations were also enriched in fatty acid metabolism and propanoate metabolism (
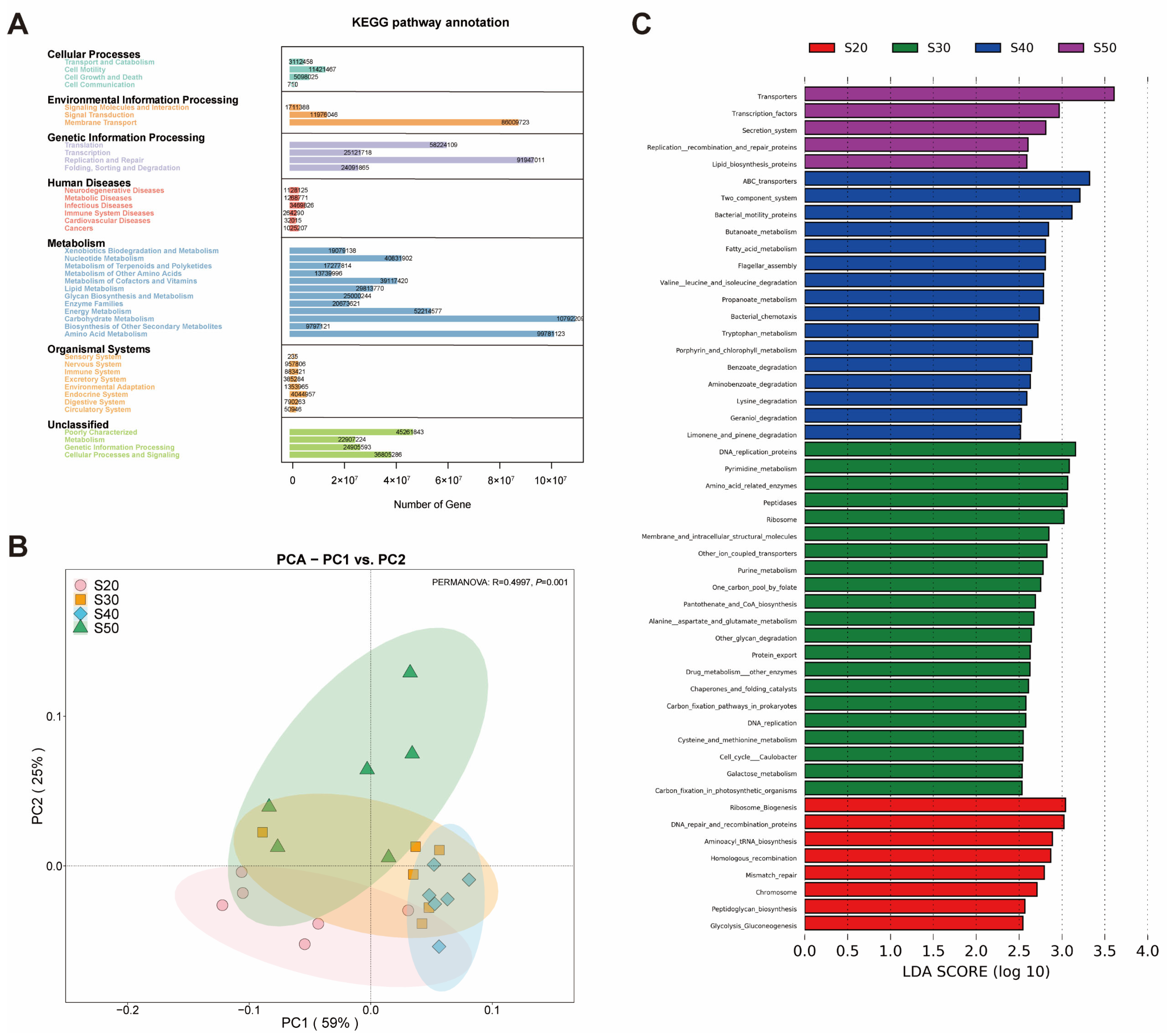
Figure 3C), which was consistent with our findings in vivo.
Prevotella_ruminicola (
P. ruminicola) plays an important role in the metabolism of proteins and peptides in the rumen [
42].
P. ruminicola has the greatest range and activity of dipeptidyl peptidases, which release dipeptides from the N-terminus of peptides [
43] and may be the main reason for the higher substrate N digestibility in S30 and S40 and also the strong positive correlation of this microbe with ND (
Figure 5B). A recent study [
44] showed that lower methane emissions and higher feed conversion efficiency were associated with a higher abundance of
P. ruminicola, which was also validated from the results that substrates DMD and ND were increased in S30 and S40, while methane production requires subsequent experiments to verify. Furthermore, it was observed that the abundance of
Fibrobacter_succinogenes (
F. succinogenes), which is considered to be a key cellulolytic bacterium that promotes the metabolism of other microbiota members by degrading cellulose into soluble sugars and succinate [
45], was up-regulated in S30 and S40. In spite of the weak peptidolytic activity, studies have reported that when
P. ruminicola was co-cultured with
F. succinogenes, the substrate nutrition digestibility was increased compared with culturing
F. succinogenes alone [
46]. In addition, the bacterial functions were enriched in related amino acid metabolism and carbohydrate metabolism at SP (% of CP) of 30 and 40, which also verified the higher substrate nutrient utilization obtained in S30 and S40 in our study.
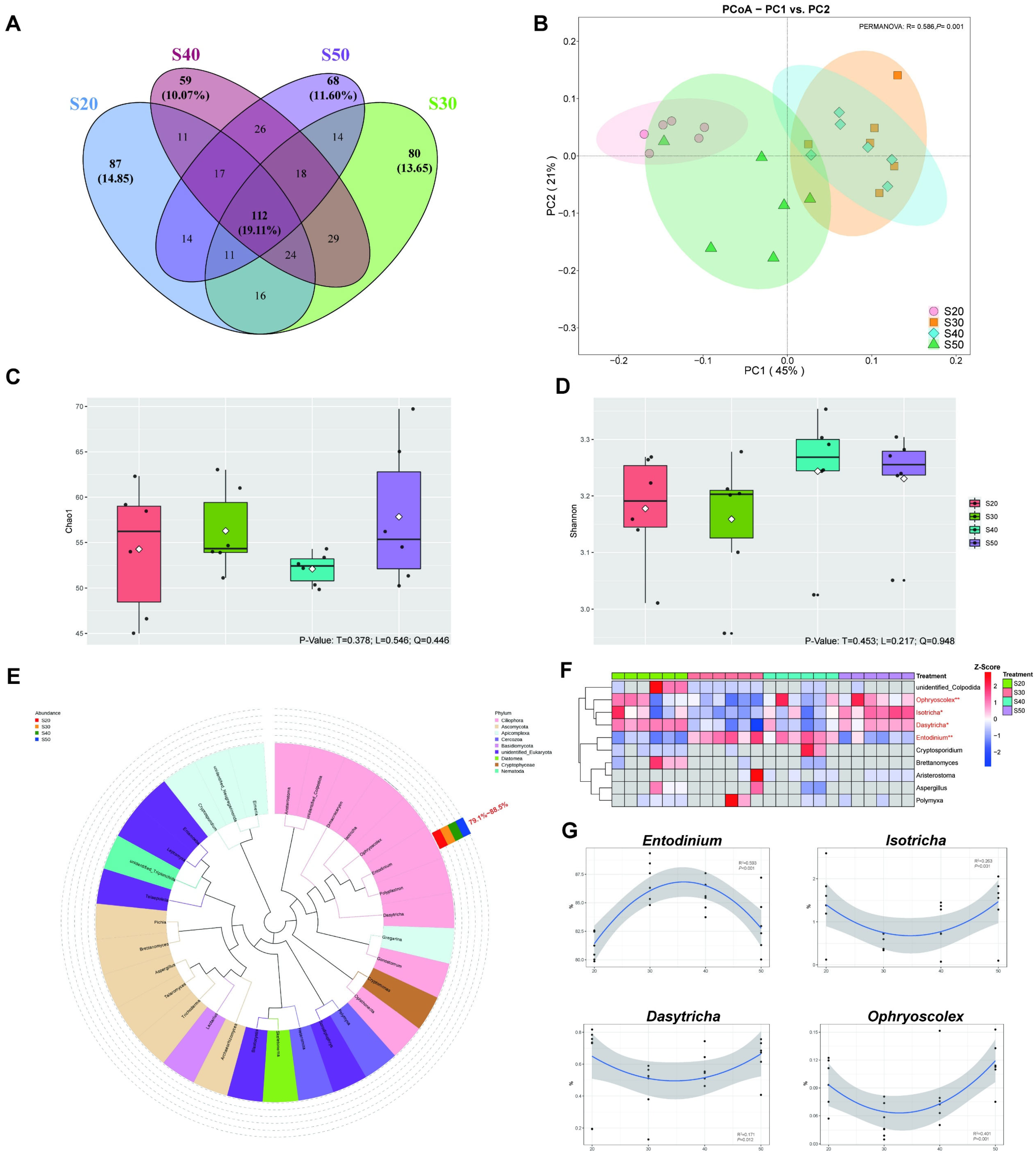
Similar to most studies on rumen protozoa [
47,
48],
Entodinium was the most abundant genus of ciliate protozoa, occupying an average of 83.8% in this study. Polyorach et al. [
49] and Chanjula et al. [
50] found that different dietary N sources had no effect on the number of ruminal protozoa, which may lead to different protozoa species composition. In our study, SP levels were modified by the addition of different proportions of N, while
Entodinium was up-regulated when SP (% of CP) was 30 and 40.
Entodinium could rapidly hydrolyze true protein (e.g., casein) and has high dipeptidase activity, with peptides and amino acids as main products [
51,
52]. However, it has been proposed that urea plays a minor role in the N metabolism of
Entodinium when used as a substrate [
53], which indicates that the change in the addition of urea as a substrate in our study might not be the reason for affecting the abundance of
Entodinium. On the other hand, the abundance of
Isotricha and
Ophryoscolex was reduced with S30 and S40, and the species members in this group of microbes exhibit lower proteolytic activity [
54]. Although the abundance of these protozoa genera was relatively lower, more experiments are required to reveal the association with SP levels in the future.
The interaction of bacteria and protozoa plays an important role in the degradation of substrates in this experiment, with
Prevotella and
Entodinium being strongly and positively correlated (
Figure 5A). Similarly, Elsayed Mickdam et al. [
55] reported that the relative abundance of
Prevotella and
Entodinium was elevated in the presence of feed-induced subacute ruminal acidosis in vitro, which might be related to lower ruminal pH, while
Prevotella could be used as a probiotic to prevent acidosis [
56]. Importantly, the synergistic effect of the two genera resulted in higher N efficiency of substrates, while Prevotella was negatively correlated with
Isotricha and
Ophryoscolex, which may be a result of phagocytosis of protozoa and bacteria or the autolysis of protozoa [
57].
Last but not least, a limitation of this study is that the diets were not configured with constant soluble carbohydrates content, which could potentially affect the N retention of the ruminal SP. Different soluble carbohydrate levels may exacerbate differences between treatments, such as NH3-N production and MCP synthesis. Therefore, the consistency of soluble carbohydrates needs to be refined in subsequent experiments by configuring the SP gradient.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}