
Habitual Diet Pattern Associations with Gut Microbiome Diversity and Composition: Results from a Chinese Adult Cohort


Abstract
:1. Introduction
2. Methods
2.1. Study Participants
2.2. Stool Sample Collection
2.3. DNA Extraction and 16S rRNA Gene Sequencing
2.4. Dietary Data Collection
2.5. Dietary Pattern Analysis
2.6. Statistical Analysis
3. Results
3.1. Study Sample Characteristics
3.2. Data-Driven Posteriori Dietary Patterns
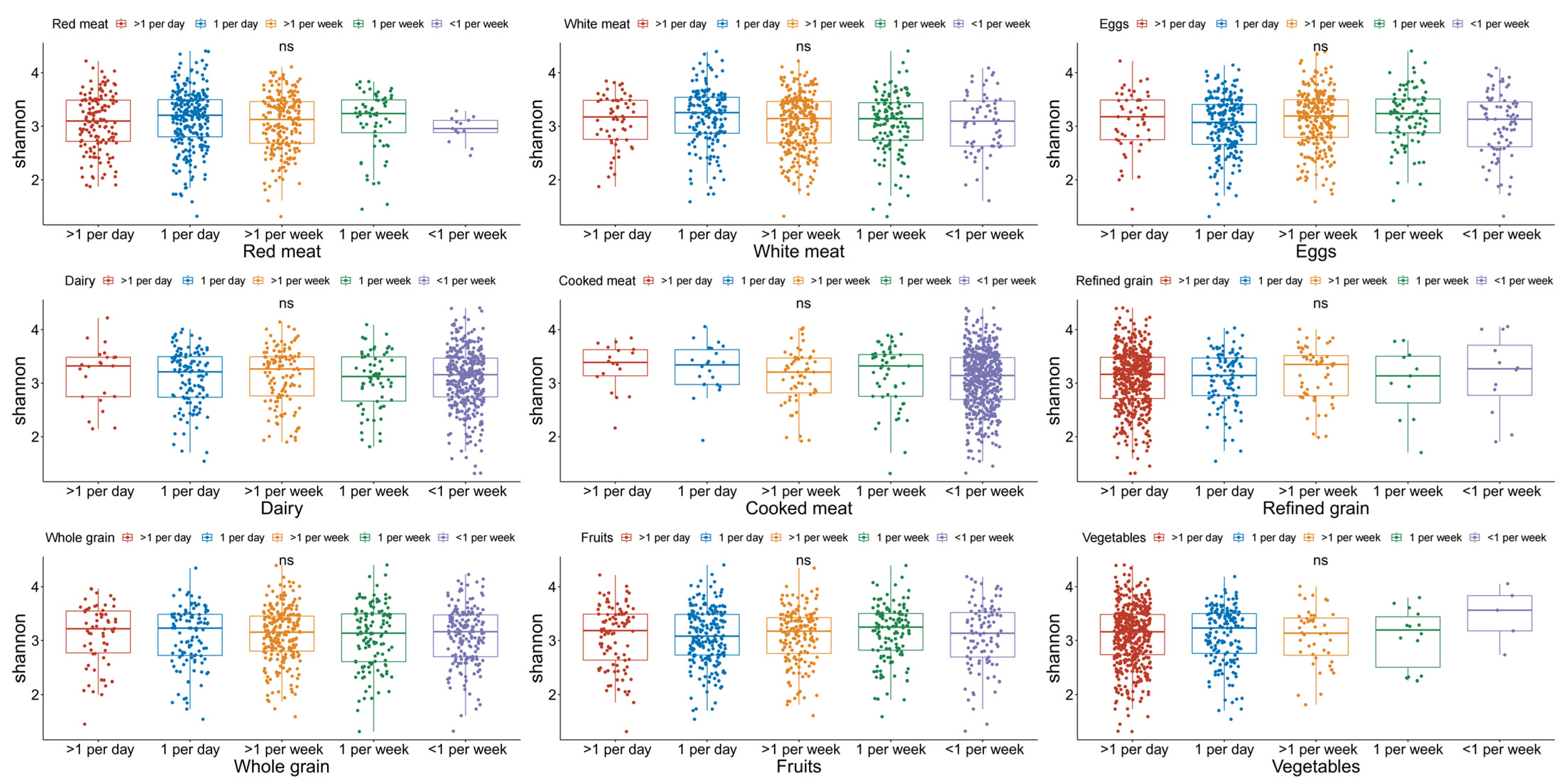
3.3. α-Diversity Indexes Distributed by Food Intake Frequencies
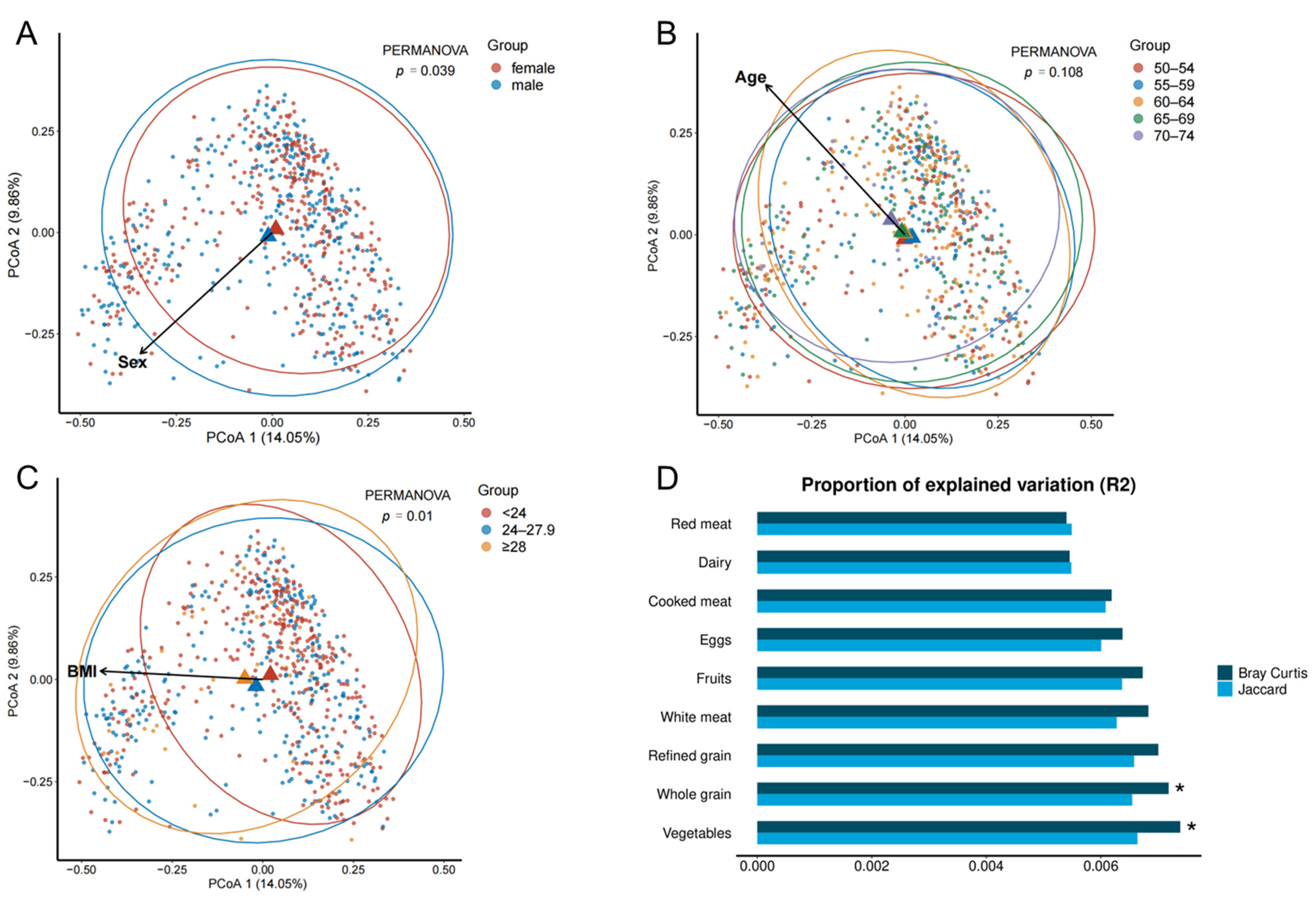
3.4. Associations between Dietary Variables and β-Diversity
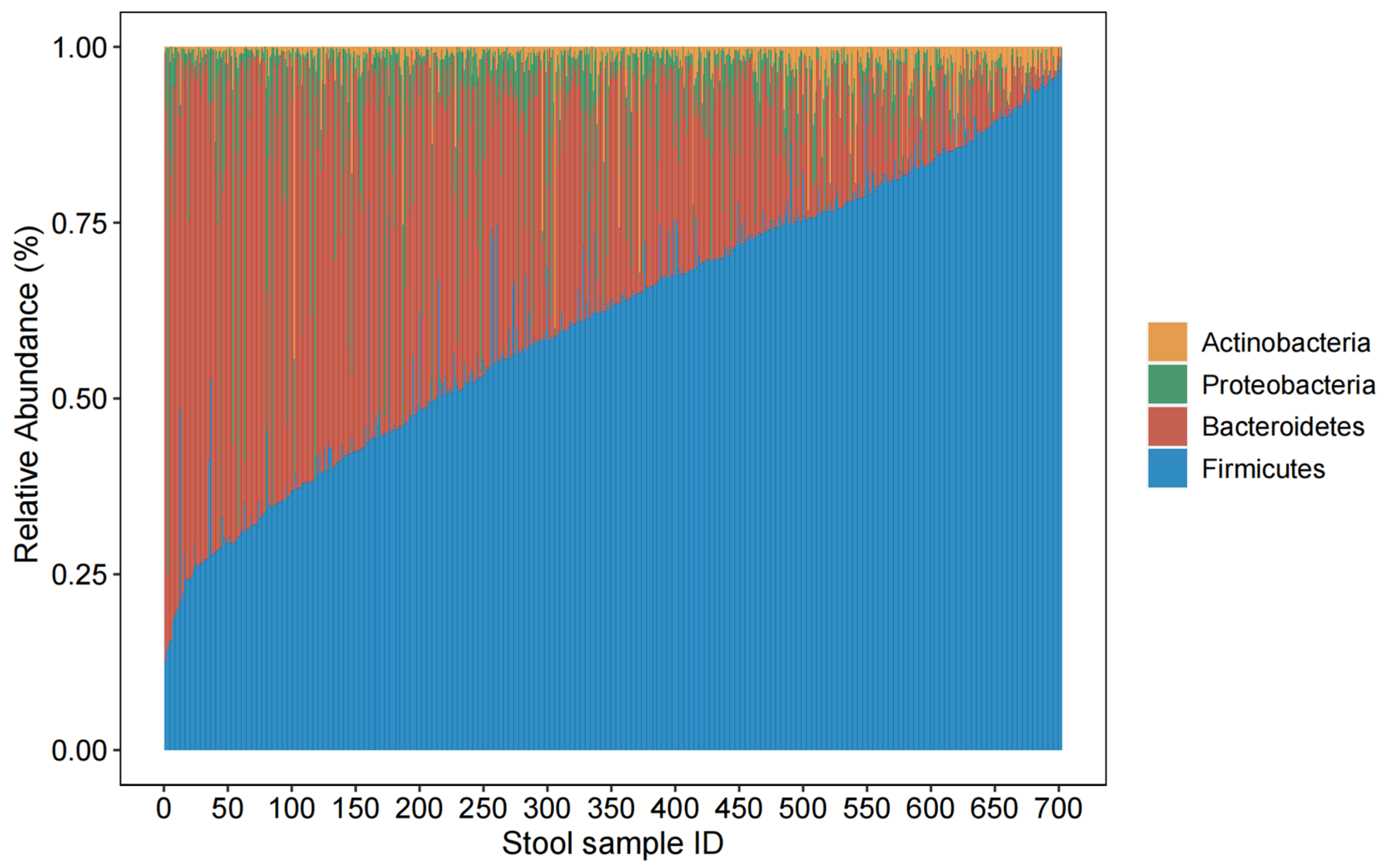
3.5. Associations between Dietary Variables and Relative Abundances of Taxa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schroeder, B.O.; Bäckhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Kundu, P.; Blacher, E.; Elinav, E.; Pettersson, S. Our Gut Microbiome: The Evolving Inner Self. Cell 2017, 171, 1481–1493. [Google Scholar] [CrossRef] [Green Version]
- Muller, P.A.; Matheis, F.; Schneeberger, M.; Kerner, Z.; Jové, V.; Mucida, D. Microbiota-modulated CART(+) enteric neurons autonomously regulate blood glucose. Science 2020, 370, 314–321. [Google Scholar] [CrossRef]
- Janney, A.; Powrie, F.; Mann, E.H. Host-microbiota maladaptation in colorectal cancer. Nature 2020, 585, 509–517. [Google Scholar] [CrossRef]
- Ruff, W.E.; Greiling, T.M.; Kriegel, M.A. Host-microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 2020, 18, 521–538. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.t.; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Clément, K. The gut microbiome, diet, and links to cardiometabolic and chronic disorders. Nat. Rev. Nephrol. 2016, 12, 169–181. [Google Scholar] [CrossRef]
- Wolter, M.; Grant, E.T.; Boudaud, M.; Steimle, A.; Pereira, G.V.; Martens, E.C.; Desai, M.S. Leveraging diet to engineer the gut microbiome. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 885–902. [Google Scholar] [CrossRef]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet-microbiota interactions and personalized nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Sklar, J.; Jiang, L.; Natarajan, L.; Knight, R.; Belkaid, Y. Host variables confound gut microbiota studies of human disease. Nature 2020, 587, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, R.C.E.; Jackson, M.A.; Pallister, T.; Skinner, J.; Spector, T.D.; Welch, A.A.; Steves, C.J. Use of dietary indices to control for diet in human gut microbiota studies. Microbiome 2018, 6, 77. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Fragiadakis, G.K.; Wastyk, H.C.; Robinson, J.L.; Sonnenburg, E.D.; Sonnenburg, J.L.; Gardner, C.D. Long-term dietary intervention reveals resilience of the gut microbiota despite changes in diet and weight. Am. J. Clin. Nutr. 2020, 111, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Koponen, K.K.; Salosensaari, A.; Ruuskanen, M.O.; Havulinna, A.S.; Männistö, S.; Jousilahti, P.; Palmu, J.; Salido, R.; Sanders, K.; Brennan, C.; et al. Associations of healthy food choices with gut microbiota profiles. Am. J. Clin. Nutr. 2021, 114, 605–616. [Google Scholar] [CrossRef]
- Zhernakova, A.; Kurilshikov, A.; Bonder, M.J.; Tigchelaar, E.F.; Schirmer, M.; Vatanen, T.; Mujagic, Z.; Vila, A.V.; Falony, G.; Vieira-Silva, S.; et al. Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 2016, 352, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Partula, V.; Mondot, S.; Torres, M.J.; Kesse-Guyot, E.; Deschasaux, M.; Assmann, K.; Latino-Martel, P.; Buscail, C.; Julia, C.; Galan, P.; et al. Associations between usual diet and gut microbiota composition: Results from the Milieu Intérieur cross-sectional study. Am. J. Clin. Nutr. 2019, 109, 1472–1483. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-level analysis of gut microbiome variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef]
- Taylor, B.C.; Lejzerowicz, F.; Poirel, M.; Shaffer, J.P.; Jiang, L.; Aksenov, A.; Litwin, N.; Humphrey, G.; Martino, C.; Miller-Montgomery, S.; et al. Consumption of Fermented Foods Is Associated with Systematic Differences in the Gut Microbiome and Metabolome. mSystems 2020, 5, e00901-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, R.C.; Wang, Z.; Usyk, M.; Sotres-Alvarez, D.; Daviglus, M.L.; Schneiderman, N.; Talavera, G.A.; Gellman, M.D.; Thyagarajan, B.; Moon, J.Y.; et al. Gut microbiome composition in the Hispanic Community Health Study/Study of Latinos is shaped by geographic relocation, environmental factors, and obesity. Genome Biol. 2019, 20, 219. [Google Scholar] [CrossRef] [PubMed]
- Maskarinec, G.; Hullar, M.A.J.; Monroe, K.R.; Shepherd, J.A.; Hunt, J.; Randolph, T.W.; Wilkens, L.R.; Boushey, C.J.; Le Marchand, L.; Lim, U.; et al. Fecal Microbial Diversity and Structure Are Associated with Diet Quality in the Multiethnic Cohort Adiposity Phenotype Study. J. Nutr. 2019, 149, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Gallè, F.; Valeriani, F.; Cattaruzza, M.S.; Gianfranceschi, G.; Liguori, R.; Antinozzi, M.; Mederer, B.; Liguori, G.; Romano Spica, V. Mediterranean Diet, Physical Activity and Gut Microbiome Composition: A Cross-Sectional Study among Healthy Young Italian Adults. Nutrients 2020, 12, 2164. [Google Scholar] [CrossRef]
- Yu, D.; Nguyen, S.M.; Yang, Y.; Xu, W.; Cai, H.; Wu, J.; Cai, Q.; Long, J.; Zheng, W.; Shu, X.O. Long-term diet quality is associated with gut microbiome diversity and composition among urban Chinese adults. Am. J. Clin. Nutr. 2021, 113, 684–694. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, L.; Zhai, Q.; Zhao, J.; Zhang, H.; Lee, Y.K.; Lu, W.; Li, M.; Chen, W. Chinese gut microbiota and its associations with staple food type, ethnicity, and urbanization. NPJ Biofilm. Microbiomes 2021, 7, 71. [Google Scholar] [CrossRef]
- Chen, H.; Li, N.; Shi, J.; Ren, J.; Liu, C.; Zhang, Y.; Jiang, Z.; Zhang, Z.; Dai, M. Comparative evaluation of novel screening strategies for colorectal cancer screening in China (TARGET-C): A study protocol for a multicentre randomised controlled trial. BMJ Open 2019, 9, e025935. [Google Scholar] [CrossRef]
- Chen, H.; Lu, M.; Liu, C.; Zou, S.; Du, L.; Liao, X.; Dong, D.; Wei, D.; Gao, Y.; Zhu, C.; et al. Comparative Evaluation of Participation and Diagnostic Yield of Colonoscopy vs Fecal Immunochemical Test vs Risk-Adapted Screening in Colorectal Cancer Screening: Interim Analysis of a Multicenter Randomized Controlled Trial (TARGET-C). Am. J. Gastroenterol. 2020, 115, 1264–1274. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, M.; Lu, B.; Liu, C.; Ma, Y.; Liu, L.; Miao, X.; Qin, J.; Chen, H.; Dai, M. Leveraging Fecal Microbial Markers to Improve the Diagnostic Accuracy of the Fecal Immunochemical Test for Advanced Colorectal Adenoma. Clin. Transl. Gastroenterol. 2021, 12, e00389. [Google Scholar] [CrossRef]
- Vogtmann, E.; Chen, J.; Kibriya, M.G.; Chen, Y.; Islam, T.; Eunes, M.; Ahmed, A.; Naher, J.; Rahman, A.; Amir, A.; et al. Comparison of Fecal Collection Methods for Microbiota Studies in Bangladesh. Appl. Environ. Microbiol. 2017, 83, e00361-17. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.K.; Paul, S.; Dutta, C. Geography, Ethnicity or Subsistence-Specific Variations in Human Microbiome Composition and Diversity. Front. Microbiol. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faits, T.; Walker, M.E.; Rodriguez-Morato, J.; Meng, H.; Gervis, J.E.; Galluccio, J.M.; Lichtenstein, A.H.; Johnson, W.E.; Matthan, N.R. Exploring changes in the human gut microbiota and microbial-derived metabolites in response to diets enriched in simple, refined, or unrefined carbohydrate-containing foods: A post hoc analysis of a randomized clinical trial. Am. J. Clin. Nutr. 2020, 112, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.N.; Mannerås-Holm, L.; Puschmann, R.; Wu, H.; Troise, A.D.; Nijsse, B.; Boeren, S.; Bäckhed, F.; Fiedler, D.; deVos, W.M. Conversion of dietary inositol into propionate and acetate by commensal Anaerostipes associates with host health. Nat. Commun. 2021, 12, 4798. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bhatia, R.; Singh, A.; Singh, P.; Kaur, R.; Khare, P.; Purama, R.K.; Boparai, R.K.; Rishi, P.; Ambalam, P.; et al. Probiotic attributes and prevention of LPS-induced pro-inflammatory stress in RAW264.7 macrophages and human intestinal epithelial cell line (Caco-2) by newly isolated Weissella cibaria strains. Food Funct. 2018, 9, 1254–1264. [Google Scholar] [CrossRef]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Choi Hong, S.M.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The human gut bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef]
- Wei, W.; Jiang, W.; Tian, Z.; Wu, H.; Ning, H.; Yan, G.; Zhang, Z.; Li, Z.; Dong, F.; Sun, Y.; et al. Fecal g. Streptococcus and g. Eubacterium_coprostanoligenes_group combined with sphingosine to modulate the serum dyslipidemia in high-fat diet mice. Clin. Nutr. 2021, 40, 4234–4245. [Google Scholar] [CrossRef]
- Freier, T.A.; Beitz, D.C.; Li, L.; Hartman, P.A. Characterization of Eubacterium coprostanoligenes sp. nov., a cholesterol-reducing anaerobe. Int. J. Syst. Bacteriol. 1994, 44, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Cotillard, A.; Cartier-Meheust, A.; Litwin, N.S.; Chaumont, S.; Saccareau, M.; Lejzerowicz, F.; Tap, J.; Koutnikova, H.; Lopez, D.G.; McDonald, D.; et al. A posteriori dietary patterns better explain variations of the gut microbiome than individual markers in the American Gut Project. Am. J. Clin. Nutr. 2021, 115, 432–443. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N | Percentage | |
|---|---|---|
| Sex | ||
| Female | 369 | 52.56% |
| Male | 333 | 47.44% |
| Age, years | ||
| 50–54 | 188 | 26.78% |
| 55–59 | 153 | 21.79% |
| 60–64 | 181 | 25.78% |
| 65–69 | 141 | 20.09% |
| 70–74 | 39 | 5.56% |
| Smoking status | ||
| Current smoker | 515 | 73.36% |
| Past smoker | 45 | 6.41% |
| Nonsmoker | 142 | 20.23% |
| Alcohol consumption | ||
| No | 463 | 65.95% |
| Seldom | 102 | 14.53% |
| Regular | 137 | 19.52% |
| BMI, kg/m2 | ||
| <24.0 | 373 | 53.13% |
| 24.0–27.9 | 282 | 40.17% |
| ≥28.0 | 47 | 6.7% |
| Physical activity (MET, h/week) | ||
| <33.60 | 175 | 24.93% |
| 33.60–82.05 | 176 | 25.07% |
| 82.05–147.80 | 175 | 24.93% |
| ≥147.80 | 176 | 25.07% |
| Region | ||
| Changsha, Hunan | 190 | 27.07% |
| Hefei, Anhui | 92 | 13.11% |
| Kunming, Yunnan | 14 | 1.99% |
| Lanxi, Zhejiang | 154 | 21.94% |
| Taizhou, Zhejiang | 164 | 23.36% |
| Xuzhou, Jiangsu | 88 | 12.54% |
| Food Group | >1 per Day | 1 per Day | >1 per Week | 1 per Week | <1 per Week |
|---|---|---|---|---|---|
| Red meat (pork, beef, lamb, etc.) | 142 (20.23%) | 272 (38.75%) | 211 (30.06%) | 64 (9.12%) | 13 (1.85%) |
| White meat (fish and poultry) | 57 (8.12%) | 170 (24.22%) | 280 (39.89%) | 131 (18.66%) | 64 (9.12%) |
| Eggs | 51 (7.26%) | 218 (31.05%) | 263 (37.46%) | 88 (12.54%) | 82 (11.68%) |
| Dairy products (milk, yoghurt, etc.) | 23 (3.28%) | 123 (17.52%) | 121 (17.24%) | 66 (9.40%) | 369 (52.56%) |
| Cooked and cured meats (e.g., sausages) | 18 (2.56%) | 22 (3.13%) | 57 (8.12%) | 53 (7.55%) | 552 (78.63%) |
| Refined grains (rice, flour, etc.) | 521 (74.22%) | 103 (14.67%) | 55 (7.83%) | 11 (1.57%) | 12 (1.71%) |
| Whole grains (millet, corn, sorghum, etc.) | 62 (8.83%) | 107 (15.24%) | 234 (33.33%) | 123 (17.52%) | 176 (25.07%) |
| Fruits | 98 (13.96%) | 212 (30.20%) | 167 (23.79%) | 127 (18.09%) | 98 (13.96%) |
| Vegetables | 480 (68.38%) | 159 (22.65%) | 44 (6.27%) | 14 (1.99%) | 5 (0.71%) |
| Food Group | Phylum | Class | Order | Family | Genus | Species | Value | Coef 1 | Coverage (%) 2 | Pval 3 | Qval 4 |
|---|---|---|---|---|---|---|---|---|---|
| Red meat | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Weissella | Uncultured organism | pd | −0.0379 | 28.35% | <0.0001 | 0.0300 |
| Red meat | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Weissella | pd | −0.0379 | 29.91% | <0.0001 | 0.0308 | |
| Dairy | Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Anaerostipes | uncultured organism | pd | 0.0146 | 67.95% | <0.0001 | 0.0261 |
| Dairy | Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Anaerostipes | pd | 0.0146 | 71.37% | <0.0001 | 0.0261 | |
| Cooked meat | Bacteroidetes | Bacteroidia | Bacteroidales | Barnesiellaceae | Coprobacter | pd | 0.0118 | 11.97% | <0.0001 | 0.0044 | |
| Whole grains | Firmicutes | Negativicutes | Veillonellales | Veillonellaceae | Megasphaera | uncultured organism | mul_pd | 0.0420 | 14.25% | <0.0001 | 0.0183 |
| Refined grains | Firmicutes | Bacilli | Lactobacillales | Lactobacillaceae | Lactobacillus | uncultured organism | pw | 0.0602 | 13.82% | 0.0001 | 0.0763 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Eubacterium coprostanoligenes group | uncultured organism | pd | −0.0767 | 23.50% | <0.0001 | 0.0123 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Eubacterium coprostanoligenes group | uncultured organism | mul_pd | −0.0737 | 23.50% | <0.0001 | 0.0140 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | uncultured | pd | −0.0389 | 43.87% | <0.0001 | 0.0156 | |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Eubacterium coprostanoligenes group | uncultured organism | mul_pw | −0.0746 | 23.50% | <0.0001 | 0.0173 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | uncultured | mul_pw | −0.0394 | 43.87% | <0.0001 | 0.0173 | |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Christensenellaceae | Christensenellaceae R7 group | uncultured organism | pd | −0.0573 | 27.78% | <0.0001 | 0.0226 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Christensenellaceae | Christensenellaceae R7 group | uncultured organism | mul_pd | −0.0561 | 27.78% | <0.0001 | 0.0256 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Eubacterium coprostanoligenes group | uncultured organism | pw | −0.0754 | 23.50% | 0.0001 | 0.0460 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Ruminococcaceae UCG 005 | uncultured organism | mul_pw | −0.0243 | 12.68% | 0.0002 | 0.0588 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | uncultured | mul_pd | −0.0339 | 43.87% | 0.0001 | 0.0588 | |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | uncultured | pw | −0.0388 | 43.87% | 0.0002 | 0.0588 | |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | uncultured | uncultured organism | mul_pw | −0.0284 | 13.96% | 0.0002 | 0.0595 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Ruminococcaceae UCG 005 | uncultured organism | pd | −0.0230 | 12.68% | 0.0002 | 0.0629 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | uncultured | uncultured organism | pd | −0.0267 | 13.96% | 0.0003 | 0.0733 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Christensenellaceae | Christensenellaceae R7 group | uncultured organism | mul_pw | −0.0522 | 27.78% | 0.0003 | 0.0743 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Christensenellaceae | Christensenellaceae R7 group | uncultured organism | pw | −0.0572 | 27.78% | 0.0003 | 0.0743 |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Ruminococcaceae UCG 005 | uncultured organism | mul_pd | −0.0222 | 12.68% | 0.0003 | 0.0743 |
| Vegetables | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Leuconostoc | uncultured organism | mul_pw | −0.0246 | 17.81% | 0.0003 | 0.0745 |
| Vegetables | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Leuconostoc | mul_pw | −0.0246 | 17.81% | 0.0003 | 0.0745 | |
| Vegetables | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Leuconostoc | uncultured organism | pd | −0.0231 | 17.81% | 0.0005 | 0.0867 |
| Vegetables | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Leuconostoc | pd | −0.0231 | 17.81% | 0.0005 | 0.0867 | |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Ruminococcaceae UCG 005 | uncultured organism | pw | −0.0246 | 12.68% | 0.0005 | 0.0900 |
| Vegetables | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Leuconostoc | uncultured organism | mul_pd | −0.0225 | 17.81% | 0.0006 | 0.0958 |
| Vegetables | Firmicutes | Bacilli | Lactobacillales | Leuconostocaceae | Leuconostoc | mul_pd | −0.0225 | 17.81% | 0.0006 | 0.0958 | |
| Vegetables | Firmicutes | Clostridia | Clostridiales | Ruminococcaceae | Eubacterium coprostanoligenes group | pd | −0.0804 | 71.23% | 0.0006 | 0.0958 | |
| Cluster | Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Anaerostipes | uncultured organism | C | 0.0119 | 67.95% | 0.0001 | 0.0749 |
| Cluster | Firmicutes | Clostridia | Clostridiales | Lachnospiraceae | Anaerostipes | C | 0.0115 | 71.37% | 0.0001 | 0.0858 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, H.; Lu, M.; Cai, J.; Lu, B.; Luo, C.; Dai, M. Habitual Diet Pattern Associations with Gut Microbiome Diversity and Composition: Results from a Chinese Adult Cohort. Nutrients 2022, 14, 2639. https://doi.org/10.3390/nu14132639
Zhang Y, Chen H, Lu M, Cai J, Lu B, Luo C, Dai M. Habitual Diet Pattern Associations with Gut Microbiome Diversity and Composition: Results from a Chinese Adult Cohort. Nutrients. 2022; 14(13):2639. https://doi.org/10.3390/nu14132639
Chicago/Turabian StyleZhang, Yuhan, Hongda Chen, Ming Lu, Jie Cai, Bin Lu, Chenyu Luo, and Min Dai. 2022. "Habitual Diet Pattern Associations with Gut Microbiome Diversity and Composition: Results from a Chinese Adult Cohort" Nutrients 14, no. 13: 2639. https://doi.org/10.3390/nu14132639