
Sesamin Promotes Osteoporotic Fracture Healing by Activating Chondrogenesis and Angiogenesis Pathways

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Molecular Docking Study
2.2. Cell Transient Transfection and Dual-Luciferase Reporter Assay
2.3. Cell Viability Assay
2.4. Capillary Tube Formation Assay
2.5. Chondrogenic Induction
2.6. Real-Time PCR
2.7. Western Blot Analysis
2.8. Osteoporotic Bone Fracture Animal Model
2.9. Mechanical Testing and Micro-Computed Tomography Analysis
2.10. Histological Studies
2.11. Immunohistochemistry and Immunofluorescence Staining
2.12. Statistical Analysis
3. Result
3.1. Sesamin Promoted Tube Formation of HUVECs through ERα
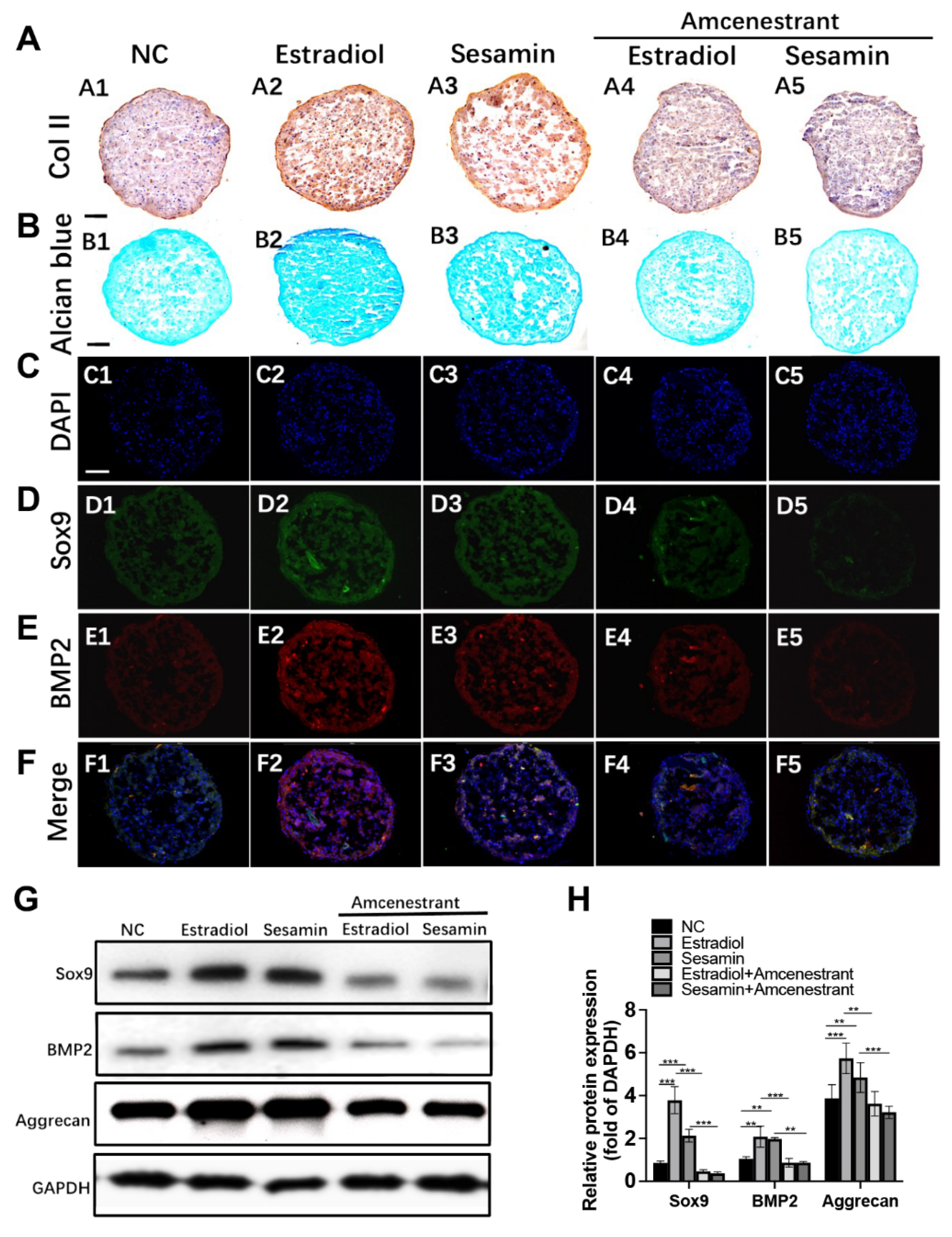
3.2. Sesamin Indued Chondrogenesis by Activating ERα and BMP2 Signaling
3.3. Sesamin Oral Administration of Improved Osteoporosis Fracture Healing In Vivo
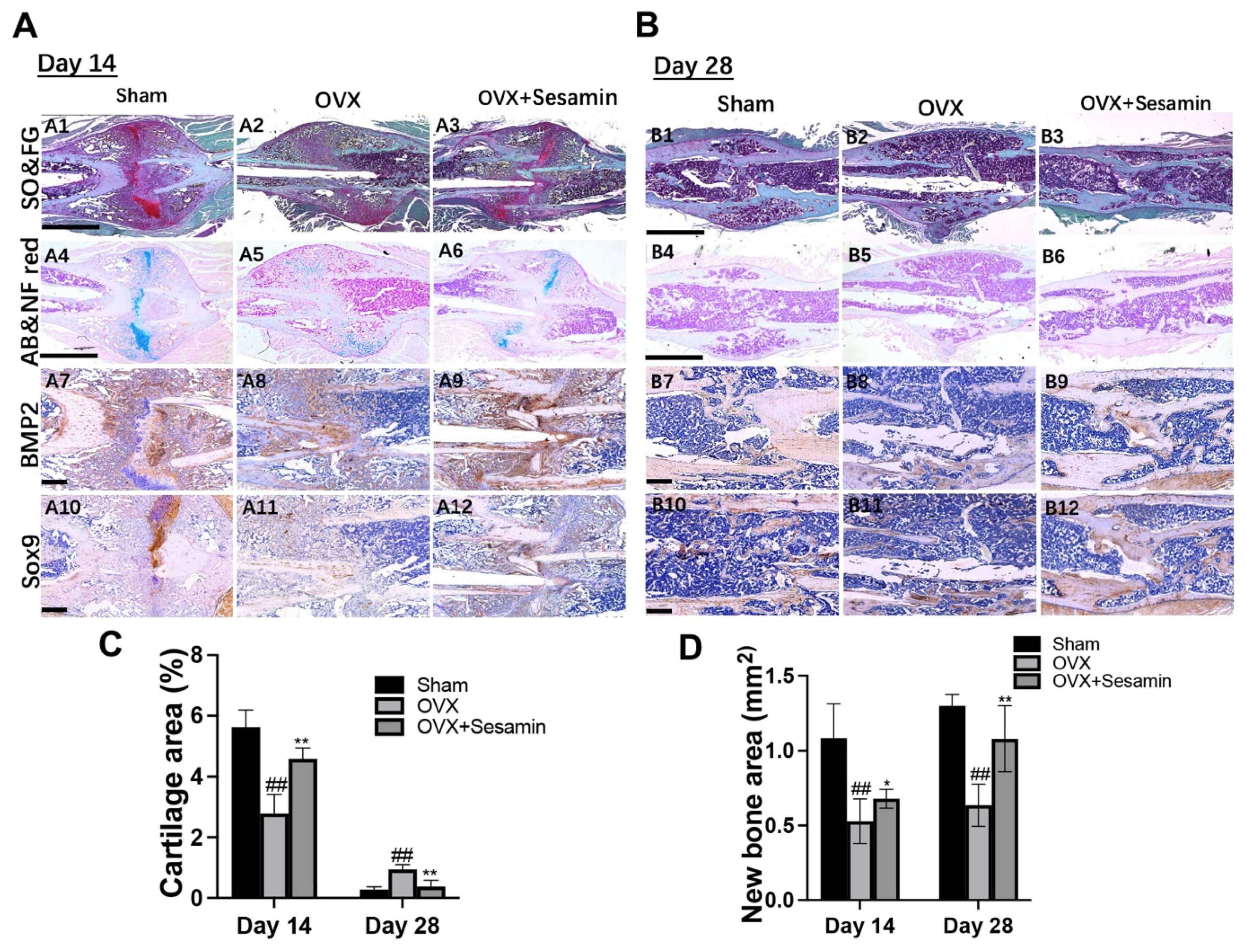
3.4. Sesamin Facilitated Osteoporotic Fracture Healing by Promoting Chondrogenesis at Early Stage
3.5. Sesamin Ameliorates Angiogenesis within Fracture Healing of OVX Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jahn-Rickert, K.; Zimmermann, E.A. Potential Role of Perilacunar Remodeling in the Progression of Osteoporosis and Implications on Age-Related Decline in Fracture Resistance of Bone. Curr. Osteoporos. Rep. 2021, 19, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Riggs, B.L.; Melton, L.J., 3rd. The worldwide problem of osteoporosis: Insights afforded by epidemiology. Bone 1995, 17, 505S–511S. [Google Scholar] [CrossRef]
- Lorentzon, M.; Johansson, H.; Harvey, N.C.; Liu, E.; Vandenput, L.; McCloskey, E.V.; Kanis, J.A. Osteoporosis and fractures in women: The burden of disease. Climacteric 2022, 25, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Beil, F.T.; Barvencik, F.; Gebauer, M.; Seitz, S.; Rueger, J.M.; Ignatius, A.; Pogoda, P.; Schinke, T.; Amling, M. Effects of estrogen on fracture healing in mice. J. Trauma 2010, 69, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; National Osteoporosis Foundation Guide Committee. A revised clinician’s guide to the prevention and treatment of osteoporosis. J. Clin. Endocrinol. Metab. 2008, 93, 2463–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compston, J.; Cooper, A.; Cooper, C.; Francis, R.; Kanis, J.A.; Marsh, D.; McCloskey, E.V.; Reid, D.M.; Selby, P.; Wilkins, M.; et al. Guidelines for the diagnosis and management of osteoporosis in postmenopausal women and men from the age of 50 years in the UK. Maturitas 2009, 62, 105–108. [Google Scholar] [CrossRef]
- Lindsay, R.; Pack, S.; Li, Z. Longitudinal progression of fracture prevalence through a population of postmenopausal women with osteoporosis. Osteoporos. Int. 2005, 16, 306–312. [Google Scholar] [CrossRef]
- Xu, D.; Xu, L.; Zhou, C.; Lee, W.Y.; Wu, T.; Cui, L.; Li, G. Salvianolic acid B promotes osteogenesis of human mesenchymal stem cells through activating ERK signaling pathway. Int. J. Biochem. Cell Biol. 2014, 51, 1–9. [Google Scholar] [CrossRef]
- Mok, S.K.; Chen, W.F.; Lai, W.P.; Leung, P.C.; Wang, X.L.; Yao, X.S.; Wong, M.S. Icariin protects against bone loss induced by oestrogen deficiency and activates oestrogen receptor-dependent osteoblastic functions in UMR 106 cells. Br. J. Pharmacol. 2010, 159, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Gao, Y.; Wang, Y.; Zhou, J.; Wei, Z.; Ma, X.; Ma, H.; Xian, C.J.; Wang, J.; Chen, K. The flavonol glycoside icariin promotes bone formation in growing rats by activating the cAMP signaling pathway in primary cilia of osteoblasts. J. Biol. Chem. 2017, 292, 20883–20896. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Feng, L.; Wang, H.; Li, Y.; Lo, J.H.T.; Zhang, X.; Lu, X.; Wang, Y.; Lin, S.; Tortorella, M.D.; et al. DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis. Nutrients 2021, 13, 4455. [Google Scholar] [CrossRef] [PubMed]
- Farr, J.N.; Rowsey, J.L.; Eckhardt, B.A.; Thicke, B.S.; Fraser, D.G.; Tchkonia, T.; Kirkland, J.L.; Monroe, D.G.; Khosla, S. Independent roles of estrogen deficiency and cellular senescence in the pathogenesis of osteoporosis: Evidence in young adult mice and older humans. J. Bone Miner. Res. 2019, 34, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Klein-Nulend, J.; van Oers, R.F.; Bakker, A.D.; Bacabac, R.G. Bone cell mechanosensitivity, estrogen deficiency, and osteoporosis. J. Biomech. 2015, 48, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Cheng, S.; Sun, K.; Wang, J.; Liu, X.; Zhao, Y.; Yang, J.; Zhao, D.; Xue, C.; Tao, Y.; et al. Changes in macrophage and inflammatory cytokine expressions during fracture healing in an ovariectomized mice model. BMC Musculoskelet. Disord. 2021, 22, 494. [Google Scholar] [CrossRef] [PubMed]
- Haffner-Luntzer, M.; Fischer, V.; Prystaz, K.; Liedert, A.; Ignatius, A. The inflammatory phase of fracture healing is influenced by oestrogen status in mice. Eur. J. Med. Res. 2017, 22, 23. [Google Scholar] [CrossRef] [Green Version]
- Chow, S.K.; Leung, K.S.; Qin, L.; Wei, F.; Cheung, W.H. Callus formation is related to the expression ratios of estrogen receptors-alpha and -beta in ovariectomy-induced osteoporotic fracture healing. Arch. Orthop. Trauma Surg. 2014, 134, 1405–1416. [Google Scholar] [CrossRef]
- Wu, G.J.; Chen, J.T.; Cherng, Y.G.; Chang, C.C.; Liu, S.H.; Chen, R.M. Genistein Improves Bone Healing via Triggering Estrogen Receptor Alpha-Mediated Expressions of Osteogenesis-Associated Genes and Consequent Maturation of Osteoblasts. J. Agric. Food Chem. 2020, 68, 10639–10650. [Google Scholar] [CrossRef]
- Huang, K.; Sun, Y.Q.; Chen, X.F.; Tian, F.; Cheng, F.; Gong, Q.L.; Liu, K.B. Psoralen, a natural phytoestrogen, improves diaphyseal fracture healing in ovariectomized mice: A preliminary study. Exp. Ther. Med. 2021, 21, 368. [Google Scholar] [CrossRef]
- Kolios, L.; Sehmisch, S.; Daub, F.; Rack, T.; Tezval, M.; Stuermer, K.M.; Stuermer, E.K. Equol but not genistein improves early metaphyseal fracture healing in osteoporotic rats. Planta Med. 2009, 75, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Shi, L.; Lu, Y.F.; Wang, B.; Tang, T.; Fu, W.M.; He, W.; Li, G.; Zhang, J.F. Linc-ROR Promotes Osteogenic Differentiation of Mesenchymal Stem Cells by Functioning as a Competing Endogenous RNA for miR-138 and miR-145. Mol. Ther. Nucleic Acids 2018, 11, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Feng, L.; Huang, J.; Zhang, X.; Lin, W.; Wang, B.; Cui, L.; Lin, S.; Li, G. Asiatic acid protects articular cartilage through promoting chondrogenesis and inhibiting inflammation and hypertrophy in osteoarthritis. Eur. J. Pharmacol. 2021, 907, 174265. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Yang, Z.M.; Li, Y.C.; Wang, H.X.; Lo, J.H.T.; Zhang, X.T.; Li, G. Linc-ROR promotes mesenchymal stem cells chondrogenesis and cartilage formation via regulating SOX9 expression. Osteoarthr. Cartil. 2021, 29, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Beil, F.T.; Barvencik, F.; Gebauer, M.; Beil, B.; Pogoda, P.; Rueger, J.M.; Ignatius, A.; Schinke, T.; Amling, M. Effects of increased bone formation on fracture healing in mice. J. Trauma 2011, 70, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhang, J.F.; Shi, L.; Yang, Z.M.; Wu, T.Y.; Wang, H.X.; Lin, W.P.; Lu, Y.F.; Lo, J.H.T.; Zhu, D.H.; et al. MicroRNA-378 Suppressed Osteogenesis of MSCs and Impaired Bone Formation via Inactivating Wnt/beta-Catenin Signaling. Mol. Ther. Nucleic Acids 2020, 21, 1017–1028. [Google Scholar] [CrossRef]
- Patoine, A.; Husseini, A.; Kasaai, B.; Gaumond, M.H.; Moffatt, P. The osteogenic cell surface marker BRIL/IFITM5 is dispensable for bone development and homeostasis in mice. PLoS ONE 2017, 12, e0184568. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Liu, Y.; Yang, Z.; Wu, T.; Lo, H.T.; Xu, J.; Zhang, J.; Lin, W.; Zhang, J.; Feng, L.; et al. Vasoactive Intestinal Peptide Promotes Fracture Healing in Sympathectomized Mice. Calcif. Tissue Int. 2021, 109, 55–65. [Google Scholar] [CrossRef]
- Kong, L.; Wang, Y.; Wang, H.; Pan, Q.; Zuo, R.; Bai, S.; Zhang, X.; Lee, W.Y.; Kang, Q.; Li, G. Conditioned media from endothelial progenitor cells cultured in simulated microgravity promote angiogenesis and bone fracture healing. Stem Cell Res. Ther. 2021, 12, 47. [Google Scholar] [CrossRef]
- Prince, R.L.; Smith, M.; Dick, I.M.; Price, R.I.; Webb, P.G.; Henderson, N.K.; Harris, M.M. Prevention of postmenopausal osteoporosis. A comparative study of exercise, calcium supplementation, and hormone-replacement therapy. N. Engl. J. Med. 1991, 325, 1189–1195. [Google Scholar] [CrossRef]
- Lo, K.W.; Ashe, K.M.; Kan, H.M.; Laurencin, C.T. The role of small molecules in musculoskeletal regeneration. Regen. Med. 2012, 7, 535–549. [Google Scholar] [CrossRef] [Green Version]
- Vaananen, H.K.; Harkonen, P.L. Estrogen and bone metabolism. Maturitas 1996, 23, S65–S69. [Google Scholar] [CrossRef]
- Nelson, H.D.; Humphrey, L.L.; Nygren, P.; Teutsch, S.M.; Allan, J.D. Postmenopausal hormone replacement therapy: Scientific review. JAMA 2002, 288, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Kongtawelert, P.; Wudtiwai, B.; Shwe, T.H.; Pothacharoen, P.; Phitak, T. Inhibition of programmed death ligand 1 (PD-L1) expression in breast cancer cells by sesamin. Int. Immunopharmacol. 2020, 86, 106759. [Google Scholar] [CrossRef] [PubMed]
- Scranton, P.E., Jr.; McMaster, J.H.; Diamond, P.E. Hormone suppression of DNA synthesis in cultured chondrocyte and osteosarcoma cell line. Clin. Orthop. Relat. Res. 1975, 112, 340–348. [Google Scholar] [CrossRef]
- Maneix, L.; Beauchef, G.; Servent, A.; Wegrowski, Y.; Maquart, F.X.; Boujrad, N.; Flouriot, G.; Pujol, J.P.; Boumediene, K.; Galera, P.; et al. 17Beta-oestradiol up-regulates the expression of a functional UDP-glucose dehydrogenase in articular chondrocytes: Comparison with effects of cytokines and growth factors. Rheumatology 2008, 47, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Srisuthtayanont, W.; Pruksakorn, D.; Kongtawelert, P.; Pothacharoen, P. Effects of sesamin on chondroitin sulfate proteoglycan synthesis induced by interleukin-1beta in human chondrocytes. BMC Complement. Altern. Med. 2017, 17, 286. [Google Scholar] [CrossRef] [Green Version]
- Bord, S.; Horner, A.; Beavan, S.; Compston, J. Estrogen receptors alpha and beta are differentially expressed in developing human bone. J. Clin. Endocrinol. Metab. 2001, 86, 2309–2314. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, M.K.; Erlandsson, M.; Alatalo, S.L.; Windahl, S.; Andersson, G.; Halleen, J.M.; Carlsten, H.; Gustafsson, J.A.; Ohlsson, C. Estrogen receptor alpha, but not estrogen receptor beta, is involved in the regulation of the OPG/RANKL (osteoprotegerin/receptor activator of NF-kappa B ligand) ratio and serum interleukin-6 in male mice. J. Endocrinol. 2001, 171, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Jessop, H.; Suswillo, R.; Zaman, G.; Lanyon, L. Endocrinology: Bone adaptation requires oestrogen receptor-alpha. Nature 2003, 424, 389. [Google Scholar] [CrossRef]
- Ikedo, A.; Imai, Y. Estrogen receptor alpha in mature osteoblasts regulates the late stage of bone regeneration. Biochem. Biophys Res. Commun. 2021, 559, 238–244. [Google Scholar] [CrossRef]
- Zhou, S.; Turgeman, G.; Harris, S.E.; Leitman, D.C.; Komm, B.S.; Bodine, P.V.; Gazit, D. Estrogens activate bone morphogenetic protein-2 gene transcription in mouse mesenchymal stem cells. Mol. Endocrinol. 2003, 17, 56–66. [Google Scholar] [CrossRef]
- Rosen, V. BMP2 signaling in bone development and repair. Cytokine Growth Factor Rev. 2009, 20, 475–480. [Google Scholar] [CrossRef]
- Kanakaris, N.K.; Petsatodis, G.; Tagil, M.; Giannoudis, P.V. Is there a role for bone morphogenetic proteins in osteoporotic fractures? Injury 2009, 40 (Suppl. S3), S21–S26. [Google Scholar] [CrossRef]
- Tsuji, K.; Bandyopadhyay, A.; Harfe, B.D.; Cox, K.; Kakar, S.; Gerstenfeld, L.; Einhorn, T.; Tabin, C.J.; Rosen, V. BMP2 activity, although dispensable for bone formation, is required for the initiation of fracture healing. Nat. Genet. 2006, 38, 1424–1429. [Google Scholar] [CrossRef] [PubMed]
- Mi, M.; Jin, H.; Wang, B.; Yukata, K.; Sheu, T.J.; Ke, Q.H.; Tong, P.; Im, H.J.; Xiao, G.; Chen, D. Chondrocyte BMP2 signaling plays an essential role in bone fracture healing. Gene 2013, 512, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, E.J.; Lee, S.Y.; Choi, Y.A.; Jung, J.C.; Bang, O.S.; Kang, S.S. BMP-2-enhanced chondrogenesis involves p38 MAPK-mediated down-regulation of Wnt-7a pathway. Mol. Cells 2006, 22, 353–359. [Google Scholar]
- Bai, Y.; Leng, Y.; Yin, G.; Pu, X.; Huang, Z.; Liao, X.; Chen, X.; Yao, Y. Effects of combinations of BMP-2 with FGF-2 and/or VEGF on HUVECs angiogenesis in vitro and CAM angiogenesis in vivo. Cell Tissue Res. 2014, 356, 109–121. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Feng, L.; Wang, M.; Li, Y.; Bai, S.; Lu, X.; Wang, H.; Zhang, X.; Wang, Y.; Lin, S.; et al. Sesamin Promotes Osteoporotic Fracture Healing by Activating Chondrogenesis and Angiogenesis Pathways. Nutrients 2022, 14, 2106. https://doi.org/10.3390/nu14102106
Yang Z, Feng L, Wang M, Li Y, Bai S, Lu X, Wang H, Zhang X, Wang Y, Lin S, et al. Sesamin Promotes Osteoporotic Fracture Healing by Activating Chondrogenesis and Angiogenesis Pathways. Nutrients. 2022; 14(10):2106. https://doi.org/10.3390/nu14102106
Chicago/Turabian StyleYang, Zhengmeng, Lu Feng, Ming Wang, Yucong Li, Shanshan Bai, Xuan Lu, Haixing Wang, Xiaoting Zhang, Yaofeng Wang, Sien Lin, and et al. 2022. "Sesamin Promotes Osteoporotic Fracture Healing by Activating Chondrogenesis and Angiogenesis Pathways" Nutrients 14, no. 10: 2106. https://doi.org/10.3390/nu14102106