
Investigating the Links between Lower Iron Status in Pregnancy and Respiratory Disease in Offspring Using Murine Models


 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Mice
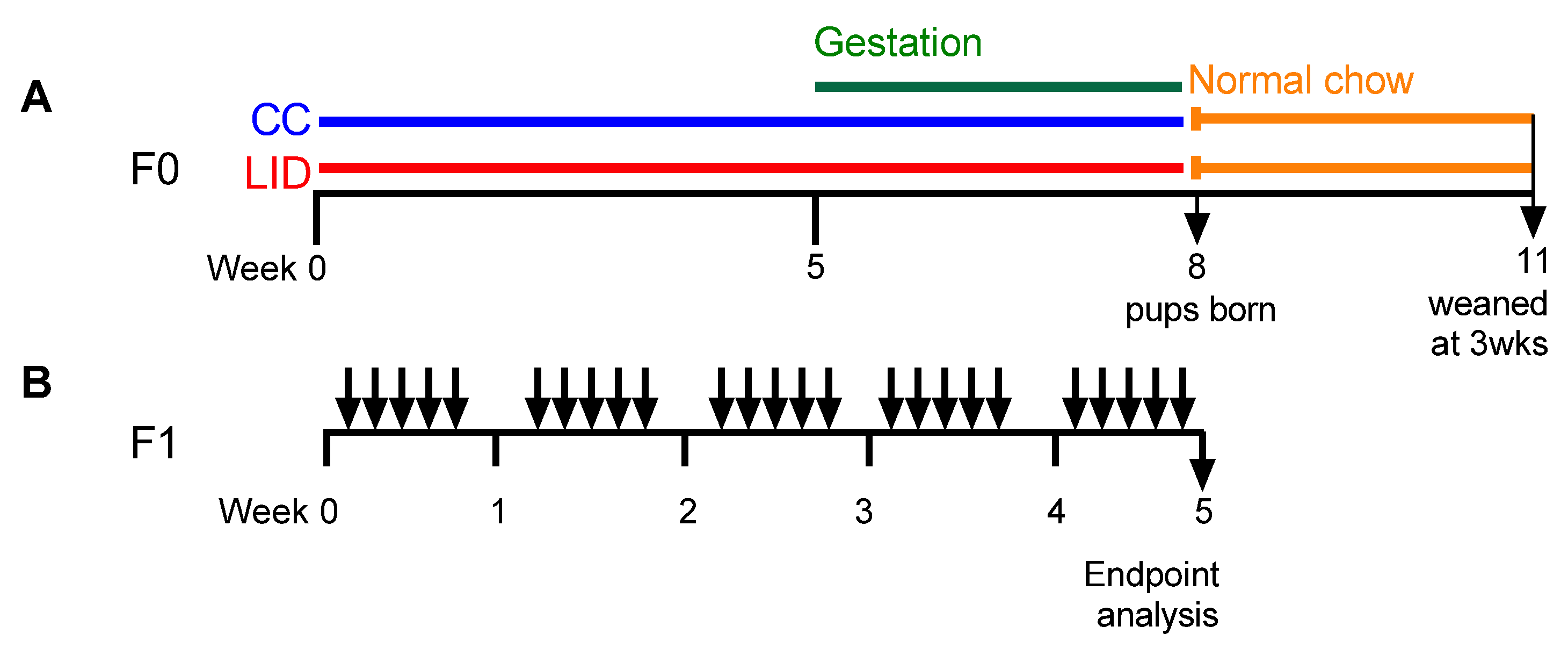
2.3. Low Systemic Iron Diet during Pregnancy and Time Mating Protocol (F0 Generation)
2.4. Establishment of HDM-Induced Experimental Asthma (F1 Generation)
2.5. Assessment of Airway Inflammation
2.6. Lung Function Analyses
2.7. Histological Analyses
2.8. Statistical Analyses
3. Results
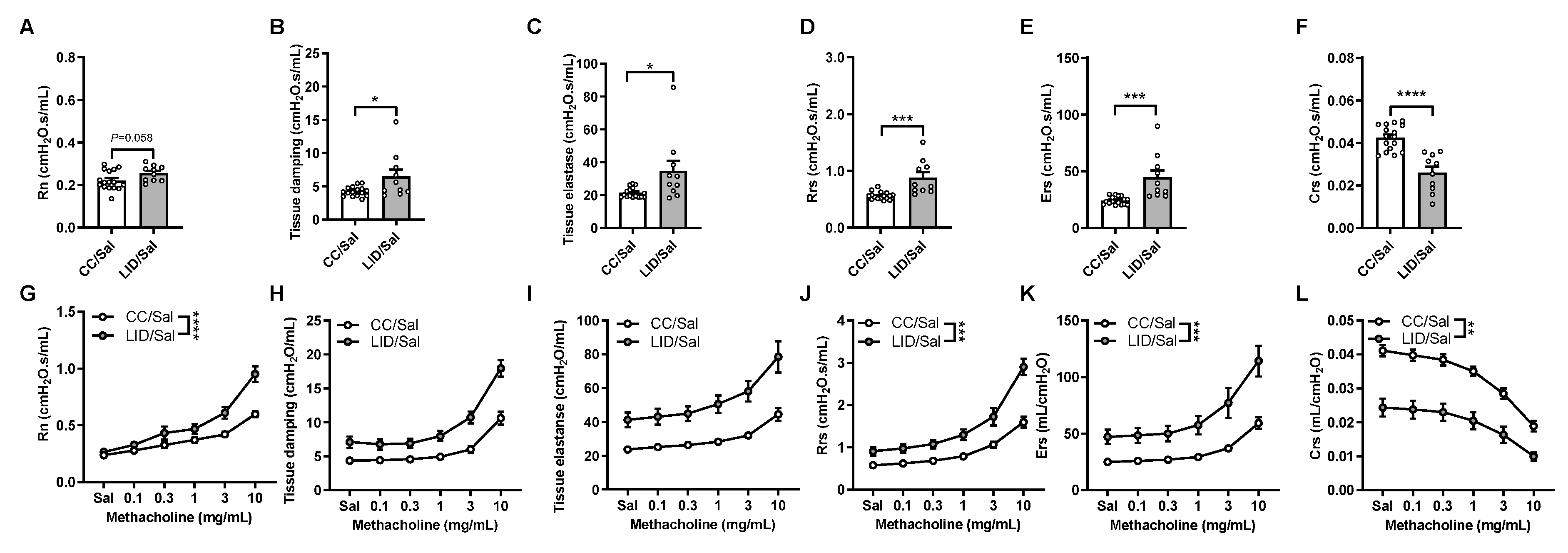
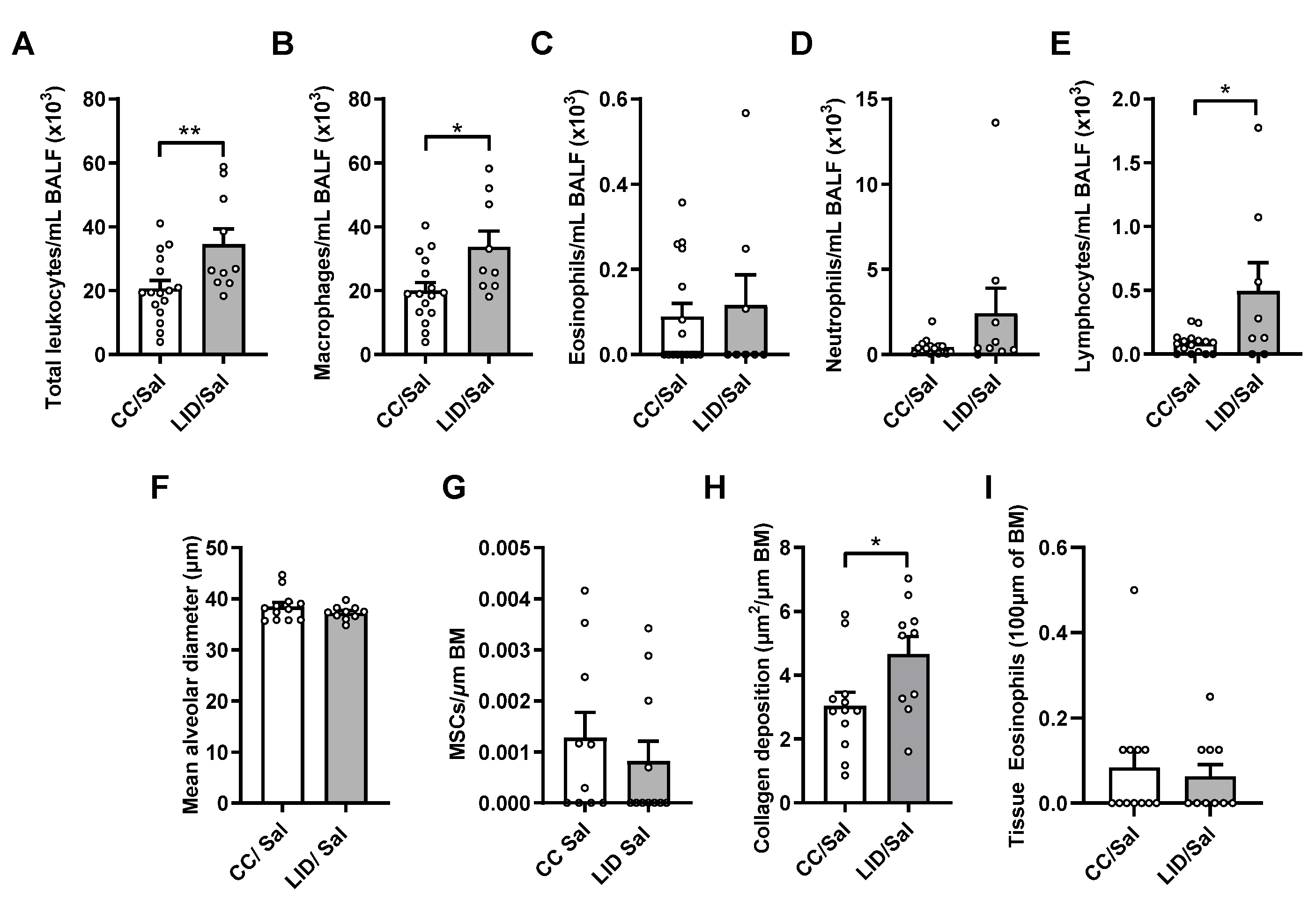
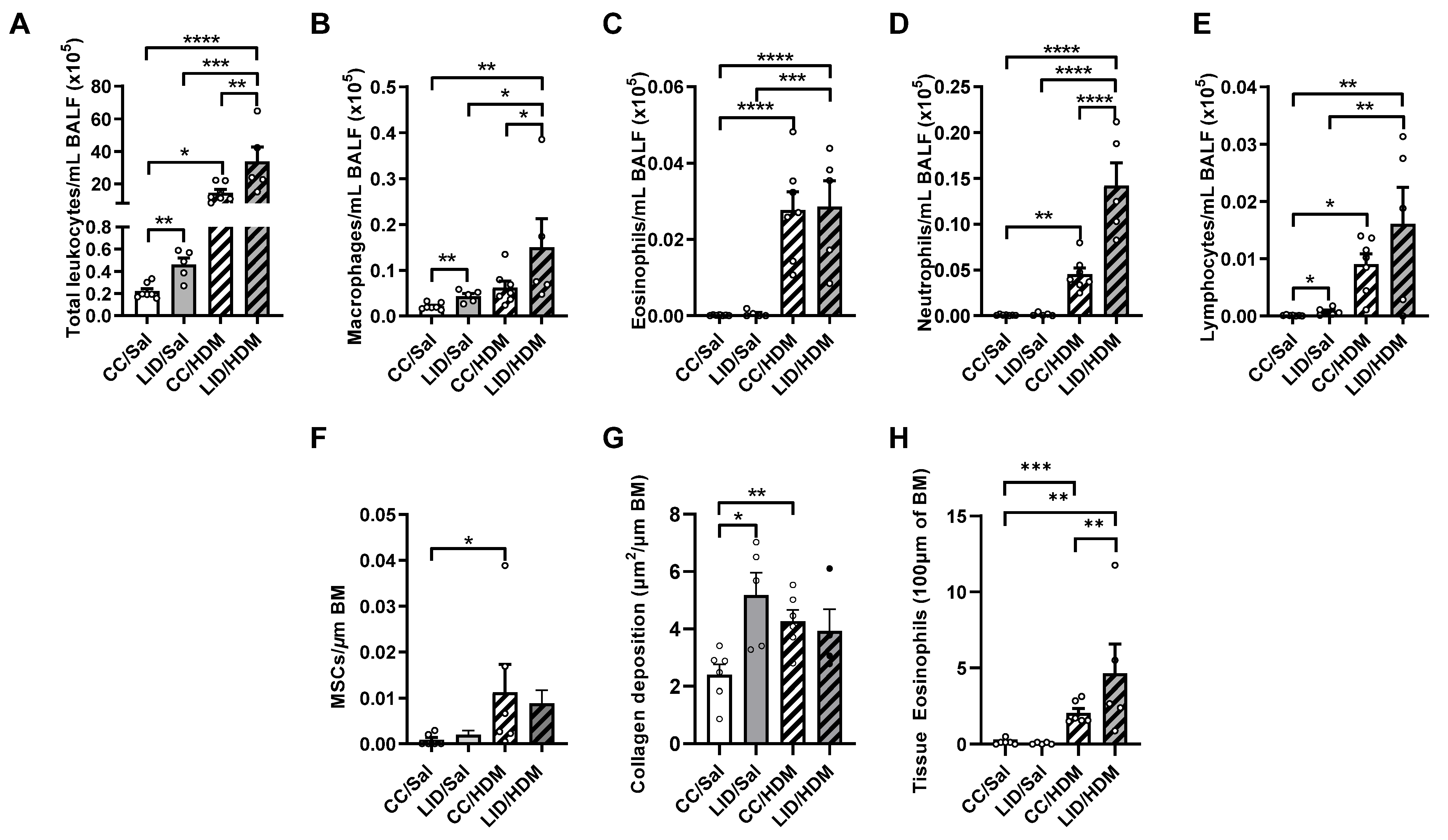
3.1. Low Iron Diet (LID) during Pregnancy Impairs Lung Function, Increases Airway Inflammation, and Increases Small Airway Collagen Deposition in Offspring
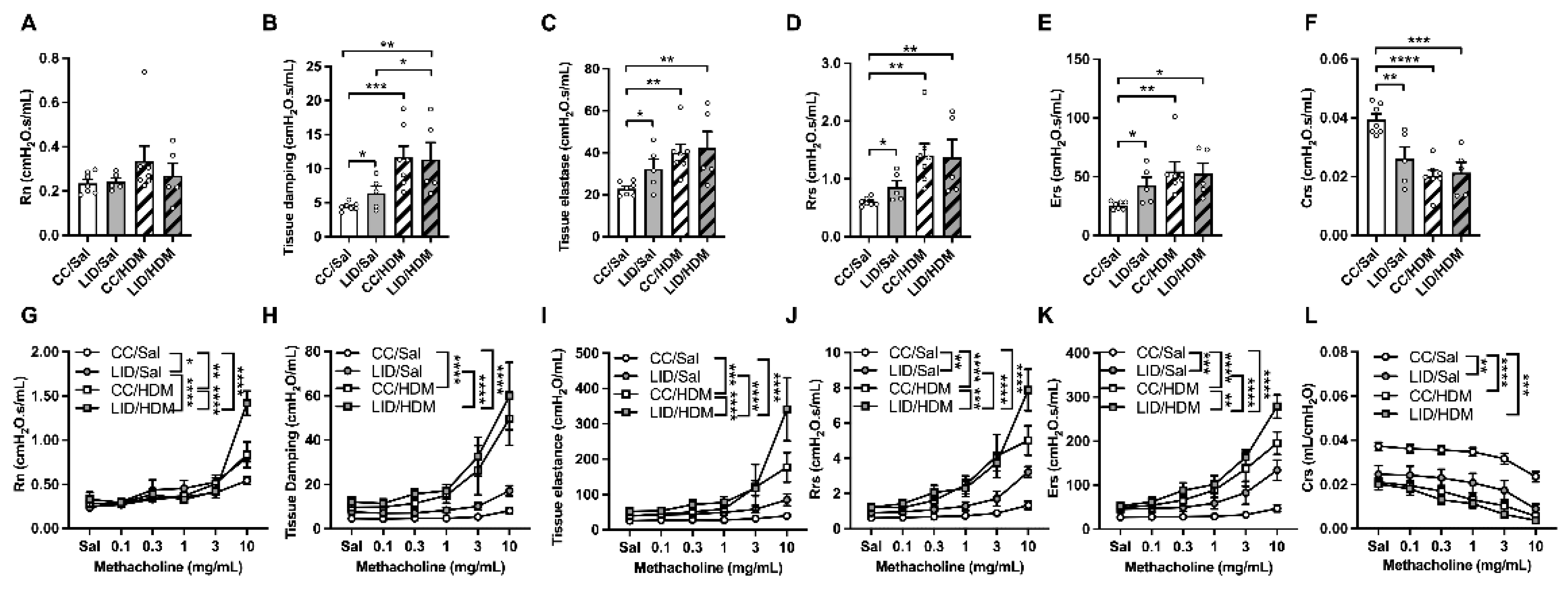
3.2. Low Iron Diet (LID) during Pregnancy Impairs Lung Function, Increases Airway Inflammation, and Increases Small Airway Collagen Deposition in Female Offspring Treated with HDM
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Abu-Ouf, N.M.; Jan, M.M. The impact of maternal iron deficiency and iron deficiency anemia on child’s health. Saudi Med. J. 2015, 36, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Annibale, B.; Capurso, G.; Chistolini, A.; D’Ambra, G.; DiGiulio, E.; Monarca, B.; DelleFave, G. Gastrointestinal causes of refractory iron deficiency anemia in patients without gastrointestinal symptoms. Am. J. Med. 2001, 111, 439–445. [Google Scholar] [CrossRef]
- Johnson-Wimbley, T.D.; Graham, D.Y. Diagnosis and management of iron deficiency anemia in the 21st century. Therap. Adv. Gastroenterol. 2011, 4, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triche, E.W.; Lundsberg, L.S.; Wickner, P.G.; Belanger, K.; Leaderer, B.P.; Bracken, M.B. Association of maternal anemia with increased wheeze and asthma in children. Ann. Allergy Asthma Immunol. 2011, 106, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Bedard, A.; Lewis, S.J.; Burgess, S.; Henderson, A.J.; Shaheen, S.O. Maternal iron status during pregnancy and respiratory and atopic outcomes in the offspring: A Mendelian randomisation study. BMJ Open Respir. Res. 2018, 5, e000275. [Google Scholar] [CrossRef] [Green Version]
- Shaheen, S.O.; Macdonald-Wallis, C.; Lawlor, D.A.; Henderson, A.J. Haemoglobin concentrations in pregnancy and respiratory and allergic outcomes in childhood: Birth cohort study. Clin. Exp. Allergy 2017, 47, 1615–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, P.N.; Tyagi, M.; Shankar, R.; Dash, D.; Kumar, A. Fetal iron status in maternal anemia. Acta Paediatr. 1996, 85, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.T.; Al Dabbagh, M.M.; Habboub, A.H.; Adel, A.; Humaidy, N.A.; Abushahin, A. Linear growth in children with iron deficiency anemia before and after treatment. J. Trop. Pediatr. 2009, 55, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.K.; Kim, R.Y.; Brown, A.C.; Mayall, J.R.; Karim, R.; Pinkerton, J.W.; Liu, G.; Martin, K.L.; Starkey, M.R.; Pillar, A.L.; et al. Crucial role for lung iron level and regulation in the pathogenesis and severity of asthma. Eur. Respir. J. 2020, 55, 1901340. [Google Scholar] [CrossRef]
- Ali, M.K.; Kim, R.Y.; Brown, A.C.; Donovan, C.; Vanka, K.S.; Mayall, J.R.; Liu, G.; Pillar, A.L.; Jones-Freeman, B.; Xenaki, D.; et al. Critical role for iron accumulation in the pathogenesis of fibrotic lung disease. J. Pathol. 2020, 251, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Horvat, J.C.; Starkey, M.R.; Kim, R.Y.; Phipps, S.; Gibson, P.G.; Beagley, K.W.; Foster, P.S.; Hansbro, P.M. Early-life chlamydial lung infection enhances allergic airways disease through age-dependent differences in immunopathology. J. Allergy Clin. Immunol. 2010, 125, 617–625. [Google Scholar] [CrossRef]
- Ali, M.K.; Kim, R.Y.; Karim, R.; Mayall, J.R.; Martin, K.L.; Shahandeh, A.; Abbasian, F.; Starkey, M.R.; Loustaud-Ratti, V.; Johnstone, D.; et al. Role of iron in the pathogenesis of respiratory disease. Int. J. Biochem. Cell Biol. 2017, 88, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.; Horvat, J.C. Casting iron in the pathogenesis of fibrotic lung disease. Am. J. Respir. Cell Mol. Biol. 2021, 65, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chang, J.; Tan, K.; Huang, S.K.; Liu, X.; Wang, X.; Cao, M.; Zhang, H.; Li, S.; Duan, X.; et al. Clioquinol attenuates pulmonary fibrosis through inactivation of fibroblasts via iron chelation. Am. J. Respir. Cell Mol. Biol. 2021, 65, 189–200. [Google Scholar] [CrossRef]
- Neves, J.; Leitz, D.; Kraut, S.; Brandenberger, C.; Agrawal, R.; Weissmann, N.; Muhlfeld, C.; Mall, M.A.; Altamura, S.; Muckenthaler, M.U. Disruption of the hepcidin/ferroportin regulatory system causes pulmonary iron overload and restrictive lung disease. eBioMedicine 2017, 20, 230–239. [Google Scholar] [CrossRef] [Green Version]
- Janssen-Heininger, Y.M.; Aesif, S.W.; van der Velden, J.; Guala, A.S.; Reiss, J.N.; Roberson, E.C.; Budd, R.C.; Reynaert, N.L.; Anathy, V. Regulation of apoptosis through cysteine oxidation: Implications for fibrotic lung disease. Ann. N. Y. Acad. Sci. 2010, 1203, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.Z.; Butler, J.J.; Cloonan, S.M. Smoking-induced iron dysregulation in the lung. Free Radic. Biol. Med. 2019, 133, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.R.; Pinniger, G.J.; Noble, P.B.; Wang, K.C.W. Intrauterine growth restriction affects diaphragm function in adult female and male mice. Pediatr. Pulmonol. 2020, 55, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Kalotas, J.O.; Wang, C.J.; Noble, P.B.; Wang, K.C.W. Intrauterine growth restriction promotes postnatal airway hyperresponsiveness independent of allergic disease. Front. Med. 2021, 8, 674324. [Google Scholar] [CrossRef]
- Weigert, R.; Dosch, N.C.; Bacsik-Campbell, M.E.; Guilbert, T.W.; Coe, C.L.; Kling, P.J. Maternal pregnancy weight gain and cord blood iron status are associated with eosinophilia in infancy. J. Perinatol. 2015, 35, 621–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, C.; Royce, S.G.; Esposito, J.; Tran, J.; Ibrahim, Z.A.; Tang, M.L.; Bailey, S.; Bourke, J.E. Differential effects of allergen challenge on large and small airway reactivity in mice. PLoS ONE 2013, 8, e74101. [Google Scholar] [CrossRef] [Green Version]
- Ward, C.; Johns, D.P.; Bish, R.; Pais, M.; Reid, D.W.; Ingram, C.; Feltis, B.; Walters, E.H. Reduced airway distensibility, fixed airflow limitation, and airway wall remodeling in asthma. Am. J. Respir. Crit. Care Med. 2001, 164, 1718–1721. [Google Scholar] [CrossRef] [PubMed]
- Ward, C.; Pais, M.; Bish, R.; Reid, D.; Feltis, B.; Johns, D.; Walters, E.H. Airway inflammation, basement membrane thickening and bronchial hyperresponsiveness in asthma. Thorax 2002, 57, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Groenman, F.A.; Rutter, M.; Wang, J.; Caniggia, I.; Tibboel, D.; Post, M. Effect of chemical stabilizers of hypoxia-inducible factors on early lung development. Am. J. Physiol. Lung Cell Mol. Physiol. 2007, 293, L557–L567. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, S.O.; Newson, R.B.; Henderson, A.J.; Emmett, P.M.; Sherriff, A.; Cooke, M.; Team, A.S. Umbilical cord trace elements and minerals and risk of early childhood wheezing and eczema. Eur. Respir. J. 2004, 24, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.Y.; Horvat, J.C.; Pinkerton, J.W.; Starkey, M.R.; Essilfie, A.T.; Mayall, J.R.; Nair, P.M.; Hansbro, N.G.; Jones, B.; Haw, T.J.; et al. MicroRNA-21 drives severe, steroid-insensitive experimental asthma by amplifying phosphoinositide 3-kinase-mediated suppression of histone deacetylase 2. J. Allergy Clin. Immunol. 2017, 139, 519–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essilfie, A.T.; Horvat, J.C.; Kim, R.Y.; Mayall, J.R.; Pinkerton, J.W.; Beckett, E.L.; Starkey, M.R.; Simpson, J.L.; Foster, P.S.; Gibson, P.G.; et al. Macrolide therapy suppresses key features of experimental steroid-sensitive and steroid-insensitive asthma. Thorax 2015, 70, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.Y.; Pinkerton, J.W.; Essilfie, A.T.; Robertson, A.A.B.; Baines, K.J.; Brown, A.C.; Mayall, J.R.; Ali, M.K.; Starkey, M.R.; Hansbro, N.G.; et al. Role for NLRP3 inflammasome-mediated, IL-1beta-dependent responses in severe, steroid-resistant asthma. Am. J. Respir. Crit. Care Med. 2017, 196, 283–297. [Google Scholar] [CrossRef]
- Pinkerton, J.W.; Kim, R.Y.; Brown, A.C.; Rae, B.E.; Donovan, C.; Mayall, J.R.; Carroll, O.R.; Khadem Ali, M.; Scott, H.A.; Berthon, B.S.; et al. Relationship between type 2 cytokine and inflammasome responses in obesity-associated asthma. J. Allergy Clin. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Rossios, C.; Pavlidis, S.; Hoda, U.; Kuo, C.H.; Wiegman, C.; Russell, K.; Sun, K.; Loza, M.J.; Baribaud, F.; Durham, A.L.; et al. Sputum transcriptomics reveal upregulation of IL-1 receptor family members in patients with severe asthma. J. Allergy Clin. Immunol. 2018, 141, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coe, G.L.; Pinkham, N.V.; Celis, A.I.; Johnson, C.; DuBois, J.L.; Walk, S.T. Dynamic gut microbiome changes in response to low-iron challenge. Appl. Environ. Microbiol. 2021, 87, e02307–e02320. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tapiainen, T.; Brinkac, L.; Lorenzi, H.A.; Moncera, K.; Tejesvi, M.V.; Salo, J.; Nelson, K.E. Vertical transmission of gut microbiome and antimicrobial resistance genes in infants exposed to antibiotics at birth. J. Infect. Dis. 2021, 224, 1236–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, J.; Hu, J.; Seki, A.; Eisele, C.; Nair, N.; Huang, R.; Tarassishin, L.; Jharap, B.; Cote-Daigneault, J.; Mao, Q.; et al. Infants born to mothers with IBD present with altered gut microbiome that transfers abnormalities of the adaptive immune system to germ-free mice. Gut 2020, 69, 42–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.; Raju Allam, V.S.R.; Paudel, K.R.; Singh, S.K.; Gulati, M.; Dhanasekaran, M.; Gupta, P.K.; Jha, N.K.; Devkota, H.P.; Gupta, G.; et al. Nutraceuticals: Unlocking newer paradigms in the mitigation of inflammatory lung diseases. Crit. Rev. Food Sci. Nutr. 2021, 1–31. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez, H.M.; Pillar, A.L.; Brown, A.C.; Kim, R.Y.; Ali, M.K.; Essilfie, A.-T.; Vanders, R.L.; Frazer, D.M.; Anderson, G.J.; Hansbro, P.M.; et al. Investigating the Links between Lower Iron Status in Pregnancy and Respiratory Disease in Offspring Using Murine Models. Nutrients 2021, 13, 4461. https://doi.org/10.3390/nu13124461
Gomez HM, Pillar AL, Brown AC, Kim RY, Ali MK, Essilfie A-T, Vanders RL, Frazer DM, Anderson GJ, Hansbro PM, et al. Investigating the Links between Lower Iron Status in Pregnancy and Respiratory Disease in Offspring Using Murine Models. Nutrients. 2021; 13(12):4461. https://doi.org/10.3390/nu13124461
Chicago/Turabian StyleGomez, Henry M., Amber L. Pillar, Alexandra C. Brown, Richard Y. Kim, Md Khadem Ali, Ama-Tawiah Essilfie, Rebecca L. Vanders, David M. Frazer, Gregory J. Anderson, Philip M. Hansbro, and et al. 2021. "Investigating the Links between Lower Iron Status in Pregnancy and Respiratory Disease in Offspring Using Murine Models" Nutrients 13, no. 12: 4461. https://doi.org/10.3390/nu13124461