High Maternal Omega-3 Supplementation Dysregulates Body Weight and Leptin in Newborn Male and Female Rats: Implications for Hypothalamic Developmental Programming

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Animals
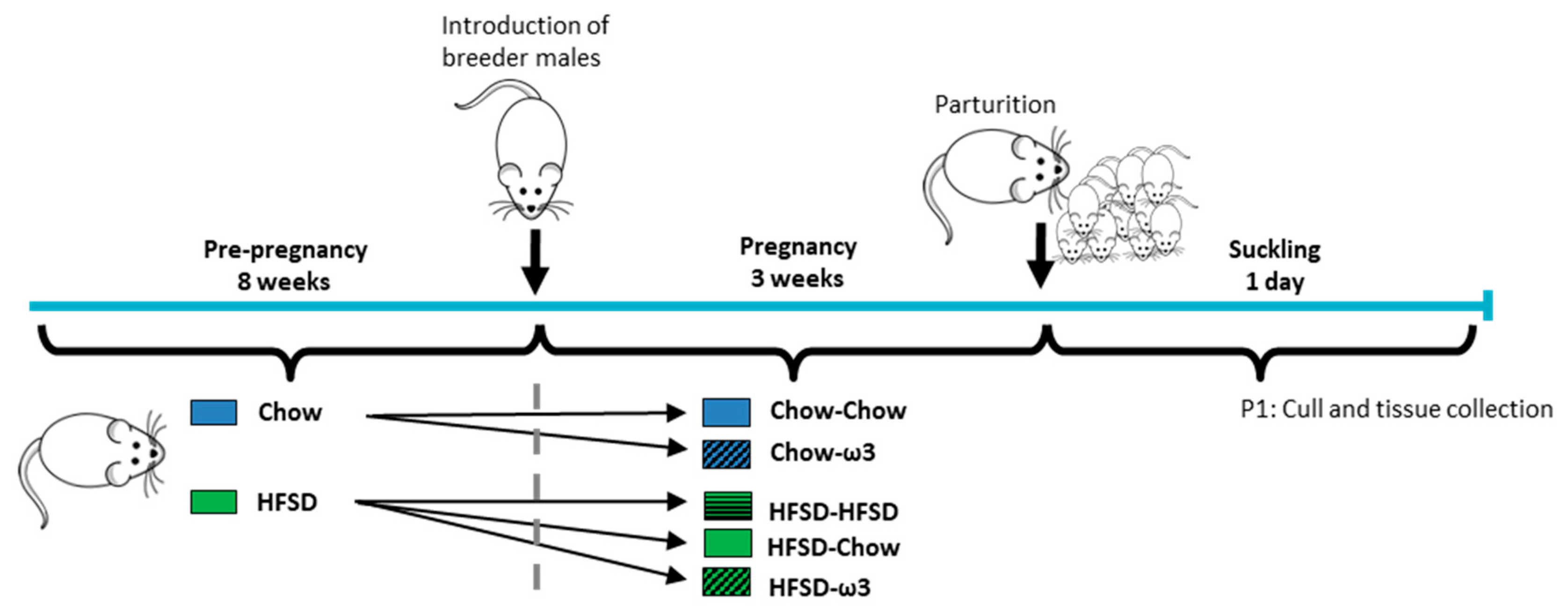
2.2. Mating and Pregnancy Dietary Protocol
2.3. Tissue Processing
2.4. Measurement of Fatty Acid Composition
2.5. Circulating Hormones
2.6. NPY Immunohistochemistry
2.7. Immunohistochemistry Analysis
2.8. Gene Expression
2.9. Statistical Analyses
3. Results
3.1. Effects of Maternal Pre- and Post-Conception Diet on Pup Survival and Morphometric Outcomes
3.2. Effects of Maternal Pre- and Post-Conception Diet on Brain Fatty Acid Composition
3.3. Effects of Maternal Pre- and Post-Conception Diet on Pup Circulating Metabolic Factors
3.4. Effects of Maternal Pre- and Post-Conception Diet on the Hypothalamic Satiety System
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfrey, K.M.; Gluckman, P.D.; Hanson, M.A. Developmental origins of metabolic disease: Life course and intergenerational perspectives. Trends Endocrinol. Metab. 2010, 21, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Frias, A.E.; Grove, K.L. Obesity: A transgenerational problem linked to nutrition during pregnancy. Semin. Reprod. Med. 2012, 30, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, S.J. Early life programming of obesity: The impact of the perinatal environment on the development of obesity and metabolic dysfunction in the offspring. Curr. Diabetes Rev. 2012, 8, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.J. Perinatal nutrition programs neuroimmune function long-term: Mechanisms and implications. Front. Neurosci. 2013, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Murrin, C.M.; Heinen, M.M.; Kelleher, C.C. Are Dietary Patterns of Mothers during Pregnancy Related to Children’s Weight Status? Evidence from the Lifeways Cross—Generational Cohort Study. AIMS Public Health 2015, 2, 274–296. [Google Scholar] [CrossRef]
- Fisk, C.M.; Crozier, S.R.; Inskip, H.M.; Godfrey, K.M.; Cooper, C.; Robinson, S.M.; Southampton Women’s Survey Study, G. Influences on the quality of young children’s diets: The importance of maternal food choices. Br. J. Nutr. 2011, 105, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Musial, B.; Vaughan, O.R.; Fernandez-Twinn, D.S.; Voshol, P.; Ozanne, S.E.; Fowden, A.L.; Sferruzzi-Perri, A.N. A Western-style obesogenic diet alters maternal metabolic physiology with consequences for fetal nutrient acquisition in mice. J. Physiol. 2017, 595, 4875–4892. [Google Scholar] [CrossRef]
- Ashino, N.G.; Saito, K.N.; Souza, F.D.; Nakutz, F.S.; Roman, E.A.; Velloso, L.A.; Torsoni, A.S.; Torsoni, M.A. Maternal high-fat feeding through pregnancy and lactation predisposes mouse offspring to molecular insulin resistance and fatty liver. J. Nutr. Biochem. 2012, 23, 341–348. [Google Scholar] [CrossRef]
- Srinivasan, M.; Katewa, S.D.; Palaniyappan, A.; Pandya, J.D.; Patel, M.S. Maternal high-fat diet consumption results in fetal malprogramming predisposing to the onset of metabolic syndrome-like phenotype in adulthood. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E792–E799. [Google Scholar] [CrossRef]
- Frihauf, J.B.; Fekete, E.M.; Nagy, T.R.; Levin, B.E.; Zorrilla, E.P. Maternal Western diet increases adiposity even in male offspring of obesity-resistant rat dams: Early endocrine risk markers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 311, R1045–R1059. [Google Scholar] [CrossRef]
- Chang, E.; Hafner, H.; Varghese, M.; Griffin, C.; Clemente, J.; Islam, M.; Carlson, Z.; Zhu, A.; Hak, L.; Abrishami, S.; et al. Programming effects of maternal and gestational obesity on offspring metabolism and metabolic inflammation. Sci. Rep. 2019, 9, 16027. [Google Scholar] [CrossRef] [PubMed]
- Dearden, L.; Bouret, S.G.; Ozanne, S.E. Sex and gender differences in developmental programming of metabolism. Mol. Metab. 2018, 15, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Sominsky, L.; Jasoni, C.L.; Twigg, H.R.; Spencer, S.J. Hormonal and nutritional regulation of postnatal hypothalamic development. J. Endocrinol. 2018, 237, R47–R64. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.G.; Draper, S.J.; Simerly, R.B. Formation of projection pathways from the arcuate nucleus of the hypothalamus to hypothalamic regions implicated in the neural control of feeding behavior in mice. J. Neurosci. 2004, 24, 2797–2805. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.G.; Draper, S.J.; Simerly, R.B. Trophic action of leptin on hypothalamic neurons that regulate feeding. Science 2004, 304, 108–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, E.C.; Cripps, R.L.; Duncan, J.S.; Barrett, P.; Mercer, J.G.; Herwig, A.; Ozanne, S.E. Developmental changes in hypothalamic leptin receptor: Relationship with the postnatal leptin surge and energy balance neuropeptides in the postnatal rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R631–R639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, S.L.; Samuelsson, A.M.; Argenton, M.; Dhonye, H.; Kalamatianos, T.; Poston, L.; Taylor, P.D.; Coen, C.W. Maternal obesity induced by diet in rats permanently influences central processes regulating food intake in offspring. PLoS ONE 2009, 4, e5870. [Google Scholar] [CrossRef]
- Morris, M.J.; Chen, H. Established maternal obesity in the rat reprograms hypothalamic appetite regulators and leptin signaling at birth. Int. J. Obes. 2009, 33, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Vickers, M.H.; Gluckman, P.D.; Coveny, A.H.; Hofman, P.L.; Cutfield, W.S.; Gertler, A.; Breier, B.H.; Harris, M. Neonatal leptin treatment reverses developmental programming. Endocrinology 2005, 146, 4211–4216. [Google Scholar] [CrossRef] [Green Version]
- Sominsky, L.; Ziko, I.; Nguyen, T.X.; Andrews, Z.B.; Spencer, S.J. Early life disruption to the ghrelin system with over-eating is resolved in adulthood in male rats. Neuropharmacology 2016, 113, 21–30. [Google Scholar] [CrossRef]
- Sominsky, L.; Ziko, I.; Nguyen, T.X.; Quach, J.; Spencer, S.J. Hypothalamic effects of neonatal diet: Reversible and only partially leptin dependent. J. Endocrinol. 2017, 234, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T. Animal studies of the functional consequences of suboptimal polyunsaturated fatty acid status during pregnancy, lactation and early post-natal life. Matern. Child Nutr. 2011, 7 (Suppl. S2), 59–79. [Google Scholar] [CrossRef] [PubMed]
- Devarshi, P.P.; Grant, R.W.; Ikonte, C.J.; Hazels Mitmesser, S. Maternal Omega-3 Nutrition, Placental Transfer and Fetal Brain Development in Gestational Diabetes and Preeclampsia. Nutrients 2019, 11, 1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanebutt, F.L.; Demmelmair, H.; Schiessl, B.; Larque, E.; Koletzko, B. Long-chain polyunsaturated fatty acid (LC-PUFA) transfer across the placenta. Clin. Nutr. 2008, 27, 685–693. [Google Scholar] [CrossRef]
- Hussain, A.; Nookaew, I.; Khoomrung, S.; Andersson, L.; Larsson, I.; Hulthen, L.; Jansson, N.; Jakubowicz, R.; Nilsson, S.; Sandberg, A.S.; et al. A maternal diet of fatty fish reduces body fat of offspring compared with a maternal diet of beef and a post-weaning diet of fish improves insulin sensitivity and lipid profile in adult C57BL/6 male mice. Acta Physiol. 2013, 209, 220–234. [Google Scholar] [CrossRef]
- Sardinha, F.L.; Fernandes, F.S.; Tavares do Carmo, M.G.; Herrera, E. Sex-dependent nutritional programming: Fish oil intake during early pregnancy in rats reduces age-dependent insulin resistance in male, but not female, offspring. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R313–R320. [Google Scholar] [CrossRef]
- Lei, X.; Zhang, W.; Liu, T.; Xiao, H.; Liang, W.; Xia, W.; Zhang, J. Perinatal supplementation with omega-3 polyunsaturated fatty acids improves sevoflurane-induced neurodegeneration and memory impairment in neonatal rats. PLoS ONE 2013, 8, e70645. [Google Scholar] [CrossRef] [Green Version]
- Colombo, J.; Gustafson, K.M.; Gajewski, B.J.; Shaddy, D.J.; Kerling, E.H.; Thodosoff, J.M.; Doty, T.; Brez, C.C.; Carlson, S.E. Prenatal DHA supplementation and infant attention. Pediatr. Res. 2016, 80, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Judge, M.P.; Harel, O.; Lammi-Keefe, C.J. Maternal consumption of a docosahexaenoic acid-containing functional food during pregnancy: Benefit for infant performance on problem-solving but not on recognition memory tasks at age 9 mo. Am. J. Clin. Nutr. 2007, 85, 1572–1577. [Google Scholar] [CrossRef] [Green Version]
- Stefanidis, A.; Spencer, S.J. Effects of neonatal overfeeding on juvenile and adult feeding and energy expenditure in the rat. PLoS ONE 2012, 7, e52130. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L.; Mark, P.J.; Mori, T.A.; Keelan, J.A.; Waddell, B.J. Maternal dietary omega-3 fatty acid supplementation reduces placental oxidative stress and increases fetal and placental growth in the rat. Biol. Reprod. 2013, 88, 37. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, H.; Doi, K.; Nagaya, N.; Okumura, H.; Nakagawa, E.; Enomoto, M.; Ono, F.; Kangawa, K. Optimum collection and storage conditions for ghrelin measurements: Octanoyl modification of ghrelin is rapidly hydrolyzed to desacyl ghrelin in blood samples. Clin. Chem. 2004, 50, 1077–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoda, H.; Kojima, M.; Matsuo, H.; Kangawa, K. Ghrelin and des-acyl ghrelin: Two major forms of rat ghrelin peptide in gastrointestinal tissue. Biochem. Biophys. Res. Commun. 2000, 279, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Ziko, I.; Sominsky, L.; Nguyen, T.X.; Yam, K.Y.; De Luca, S.; Korosi, A.; Spencer, S.J. Hyperleptinemia in Neonatally Overfed Female Rats Does Not Dysregulate Feeding Circuitry. Front. Endocrinol. 2017, 8, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sominsky, L.; Ong, L.K.; Ziko, I.; Dickson, P.W.; Spencer, S.J. Neonatal overfeeding increases capacity for catecholamine biosynthesis from the adrenal gland acutely and long-term in the male rat. Mol. Cell. Endocrinol. 2018, 470, 295–303. [Google Scholar] [CrossRef] [PubMed]
- De Luca, S.N.; Sominsky, L.; Soch, A.; Wang, H.; Ziko, I.; Rank, M.M.; Spencer, S.J. Conditional microglial depletion in rats leads to reversible anorexia and weight loss by disrupting gustatory circuitry. Brain Behav. Immun. 2019, 77, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Di Natale, M.R.; Soch, A.; Ziko, I.; De Luca, S.N.; Spencer, S.J.; Sominsky, L. Chronic predator stress in female mice reduces primordial follicle numbers: Implications for the role of ghrelin. J. Endocrinol. 2019, 241, 201–219. [Google Scholar] [CrossRef]
- Sominsky, L.; Goularte, J.F.; Andrews, Z.B.; Spencer, S.J. Acylated Ghrelin Supports the Ovarian Transcriptome and Follicles in the Mouse: Implications for Fertility. Front. Endocrinol. 2019, 9, 815. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lenz, K.M.; McCarthy, M.M. Organized for sex—Steroid hormones and the developing hypothalamus. Eur. J. Neurosci. 2010, 32, 2096–2104. [Google Scholar] [CrossRef] [Green Version]
- Haggarty, P. Fatty acid supply to the human fetus. Annu. Rev. Nutr. 2010, 30, 237–255. [Google Scholar] [CrossRef] [PubMed]
- Schaefer-Graf, U.M.; Graf, K.; Kulbacka, I.; Kjos, S.L.; Dudenhausen, J.; Vetter, K.; Herrera, E. Maternal lipids as strong determinants of fetal environment and growth in pregnancies with gestational diabetes mellitus. Diabetes Care 2008, 31, 1858–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steculorum, S.M.; Collden, G.; Coupe, B.; Croizier, S.; Lockie, S.; Andrews, Z.B.; Jarosch, F.; Klussmann, S.; Bouret, S.G. Neonatal ghrelin programs development of hypothalamic feeding circuits. J. Clin. Investig. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grove, K.L.; Smith, M.S. Ontogeny of the hypothalamic neuropeptide Y system. Physiol. Behav. 2003, 79, 47–63. [Google Scholar] [CrossRef]
- Grove, K.L.; Allen, S.; Grayson, B.E.; Smith, M.S. Postnatal development of the hypothalamic neuropeptide Y system. Neuroscience 2003, 116, 393–406. [Google Scholar] [CrossRef]
- Connor, K.L.; Vickers, M.H.; Beltrand, J.; Meaney, M.J.; Sloboda, D.M. Nature, nurture or nutrition? Impact of maternal nutrition on maternal care, offspring development and reproductive function. J. Physiol. 2012, 590, 2167–2180. [Google Scholar] [CrossRef]
- Drake, A.J.; Reynolds, R.M. Impact of maternal obesity on offspring obesity and cardiometabolic disease risk. Reproduction 2010, 140, 387–398. [Google Scholar] [CrossRef]
- Kong, L.; Chen, X.; Gissler, M.; Lavebratt, C. Relationship of prenatal maternal obesity and diabetes to offspring neurodevelopmental and psychiatric disorders: A narrative review. Int. J. Obes. 2020. [Google Scholar] [CrossRef]
- Briffa, J.F.; McAinch, A.J.; Romano, T.; Wlodek, M.E.; Hryciw, D.H. Leptin in pregnancy and development: A contributor to adulthood disease? Am. J. Physiol. Endocrinol. Metab. 2015, 308, E335–E350. [Google Scholar] [CrossRef] [Green Version]
- Bouret, S.G. Leptin, nutrition, and the programming of hypothalamic feeding circuits. Nestle Nutr. Workshop Ser. Paediatr. Programme 2010, 65, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Cowley, M.A.; Smart, J.L.; Rubinstein, M.; Cerdan, M.G.; Diano, S.; Horvath, T.L.; Cone, R.D.; Low, M.J. Leptin activates anorexigenic POMC neurons through a neural network in the arcuate nucleus. Nature 2001, 411, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Vickers, M.H.; Sloboda, D.M. Leptin as mediator of the effects of developmental programming. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Simar, D.; Lambert, K.; Mercier, J.; Morris, M.J. Maternal and postnatal overnutrition differentially impact appetite regulators and fuel metabolism. Endocrinology 2008, 149, 5348–5356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.T.; Waddell, B.J. Leptin distribution and metabolism in the pregnant rat: Transplacental leptin passage increases in late gestation but is reduced by excess glucocorticoids. Endocrinology 2003, 144, 3024–3030. [Google Scholar] [CrossRef] [Green Version]
- Kawai, M.; Yamaguchi, M.; Murakami, T.; Shima, K.; Murata, Y.; Kishi, K. The placenta is not the main source of leptin production in pregnant rat: Gestational profile of leptin in plasma and adipose tissues. Biochem. Biophys. Res. Commun. 1997, 240, 798–802. [Google Scholar] [CrossRef]
- Lepercq, J.; Challier, J.C.; Guerre-Millo, M.; Cauzac, M.; Vidal, H.; Hauguel-de Mouzon, S. Prenatal leptin production: Evidence that fetal adipose tissue produces leptin. J. Clin. Endocrinol. Metab. 2001, 86, 2409–2413. [Google Scholar] [CrossRef]
- Smith, J.T.; Waddell, B.J. Developmental changes in plasma leptin and hypothalamic leptin receptor expression in the rat: Peripubertal changes and the emergence of sex differences. J. Endocrinol. 2003, 176, 313–319. [Google Scholar] [CrossRef]
- Nozhenko, Y.; Asnani-Kishnani, M.; Rodriguez, A.M.; Palou, A. Milk Leptin Surge and Biological Rhythms of Leptin and Other Regulatory Proteins in Breastmilk. PLoS ONE 2015, 10, e0145376. [Google Scholar] [CrossRef] [Green Version]
- Korotkova, M.; Gabrielsson, B.; Lonn, M.; Hanson, L.A.; Strandvik, B. Leptin levels in rat offspring are modified by the ratio of linoleic to alpha-linolenic acid in the maternal diet. J. Lipid Res. 2002, 43, 1743–1749. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Gayle, D.; Babu, J.; Ross, M.G. Programmed obesity in intrauterine growth-restricted newborns: Modulation by newborn nutrition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R91–R96. [Google Scholar] [CrossRef]
- Banks, W.A.; Farr, S.A.; Salameh, T.S.; Niehoff, M.L.; Rhea, E.M.; Morley, J.E.; Hanson, A.J.; Hansen, K.M.; Craft, S. Triglycerides cross the blood-brain barrier and induce central leptin and insulin receptor resistance. Int. J. Obes. 2018, 42, 391–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, N.R.; Joakim Ek, C.; Dziegielewska, K.M. The neonatal blood-brain barrier is functionally effective, and immaturity does not explain differential targeting of AAV9. Nat. Biotechnol. 2009, 27, 804–805. [Google Scholar] [CrossRef] [PubMed]
- Kniesel, U.; Risau, W.; Wolburg, H. Development of blood-brain barrier tight junctions in the rat cortex. Dev. Brain Res. 1996, 96, 229–240. [Google Scholar] [CrossRef]
- Saunders, N.R.; Liddelow, S.A.; Dziegielewska, K.M. Barrier mechanisms in the developing brain. Front. Pharmacol. 2012, 3, 46. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.W.; Glendining, K.A.; Grattan, D.R.; Jasoni, C.L. Maternal Obesity in the Mouse Compromises the Blood-Brain Barrier in the Arcuate Nucleus of Offspring. Endocrinology 2016, 157, 2229–2242. [Google Scholar] [CrossRef]
- Dietrich, M.O.; Horvath, T.L. Hypothalamic control of energy balance: Insights into the role of synaptic plasticity. Trends Neurosci. 2013, 36, 65–73. [Google Scholar] [CrossRef]
- Baquero, A.F.; de Solis, A.J.; Lindsley, S.R.; Kirigiti, M.A.; Smith, M.S.; Cowley, M.A.; Zeltser, L.M.; Grove, K.L. Developmental switch of leptin signaling in arcuate nucleus neurons. J. Neurosci. 2014, 34, 9982–9994. [Google Scholar] [CrossRef] [Green Version]
- Dragano, N.R.V.; Solon, C.; Ramalho, A.F.; de Moura, R.F.; Razolli, D.S.; Christiansen, E.; Azevedo, C.; Ulven, T.; Velloso, L.A. Polyunsaturated fatty acid receptors, GPR40 and GPR120, are expressed in the hypothalamus and control energy homeostasis and inflammation. J. Neuroinflamm. 2017, 14, 91. [Google Scholar] [CrossRef] [Green Version]
- Cintra, D.E.; Ropelle, E.R.; Moraes, J.C.; Pauli, J.R.; Morari, J.; Souza, C.T.; Grimaldi, R.; Stahl, M.; Carvalheira, J.B.; Saad, M.J.; et al. Unsaturated fatty acids revert diet-induced hypothalamic inflammation in obesity. PLoS ONE 2012, 7, e30571. [Google Scholar] [CrossRef]
- Church, M.W.; Jen, K.L.; Anumba, J.I.; Jackson, D.A.; Adams, B.R.; Hotra, J.W. Excess omega-3 fatty acid consumption by mothers during pregnancy and lactation caused shorter life span and abnormal ABRs in old adult offspring. Neurotoxicol. Teratol. 2010, 32, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Muhlhausler, B.S.; Miljkovic, D.; Fong, L.; Xian, C.J.; Duthoit, E.; Gibson, R.A. Maternal omega-3 supplementation increases fat mass in male and female rat offspring. Front. Genet. 2011, 2, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, M.W.; Jen, K.L.; Dowhan, L.M.; Adams, B.R.; Hotra, J.W. Excess and deficient omega-3 fatty acid during pregnancy and lactation cause impaired neural transmission in rat pups. Neurotoxicol. Teratol. 2008, 30, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Muhlhausler, B.S.; Gibson, R.A.; Makrides, M. Effect of long-chain polyunsaturated fatty acid supplementation during pregnancy or lactation on infant and child body composition: A systematic review. Am. J. Clin. Nutr. 2010, 92, 857–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieland, L.S. Omega-3 Fatty Acid Addition during Pregnancy: Summary of a Cochrane Review. Explore 2019, 15, 168–169. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.L.; Mark, P.J.; Keelan, J.A.; Barden, A.; Mas, E.; Mori, T.A.; Waddell, B.J. Maternal dietary omega-3 fatty acid intake increases resolvin and protectin levels in the rat placenta. J. Lipid Res. 2013, 54, 2247–2254. [Google Scholar] [CrossRef] [Green Version]
- Akerele, O.A.; Cheema, S.K. A balance of omega-3 and omega-6 polyunsaturated fatty acids is important in pregnancy. J. Nutr. Intermed. Metab. 2016, 5, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Reichelt, A.C.; Gibson, G.D.; Abbott, K.N.; Hare, D.J. A high-fat high-sugar diet in adolescent rats impairs social memory and alters chemical markers characteristic of atypical neuroplasticity and parvalbumin interneuron depletion in the medial prefrontal cortex. Food Funct. 2019, 10, 1985–1998. [Google Scholar] [CrossRef]
- Spencer, S.J.; Basri, B.; Sominsky, L.; Soch, A.; Ayala, M.T.; Reineck, P.; Gibson, B.C.; Barrientos, R.M. High-fat diet worsens the impact of aging on microglial function and morphology in a region-specific manner. Neurobiol. Aging 2019, 74, 121–134. [Google Scholar] [CrossRef]
- Soch, A.; Bradburn, S.; Sominsky, L.; De Luca, S.N.; Murgatroyd, C.; Spencer, S.J. Effects of exercise on adolescent and adult hypothalamic and hippocampal neuroinflammation. Hippocampus 2016, 26, 1435–1446. [Google Scholar] [CrossRef]
- Cai, G.; Dinan, T.; Barwood, J.M.; De Luca, S.N.; Soch, A.; Ziko, I.; Chan, S.M.H.; Zheng, X.Y.; Li, S.; Molero, J.; et al. Neonatal overfeeding attenuates acute central pro-inflammatory effects of short-term high fat diet. Front. Neurosci. 2015, 8, 446. [Google Scholar] [CrossRef] [Green Version]
- Pellizzon, M.A.; Ricci, M.R. The common use of improper control diets in diet-induced metabolic disease research confounds data interpretation: The fiber factor. Nutr. Metab. 2018, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Middleton, P.; Gomersall, J.C.; Gould, J.F.; Shepherd, E.; Olsen, S.F.; Makrides, M. Omega-3 fatty acid addition during pregnancy. Cochrane Database Syst. Rev. 2018, 11, CD003402. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calculated Nutritional Parameters | Diet | ||
|---|---|---|---|
| Chow | HFSD | ω3 | |
| Protein (%) | 20.0 | 19.4 | 19.4 |
| Total Fat (%) | 4.8 | 20.0 | 5.0 |
| Total Carbohydrate (%) | 59.40 | No data | 63.3 |
| Sucrose (%) | 0.0 | 39.60 | 10.0 |
| Digestible Energy (MJ/kg) | 14.0 | 18.4 | 15.8 |
| Total Calculated Energy From Protein (%) | 23.0 | 19.0 | 22.0 |
| Total Calculated Energy From Lipids (%) | 12.0 | 36.0 | 10.0 |
| Calculated Fatty Acid Composition (%) | |||
| Linoleic Acid 18:2 ω6 | 1.30 | 2.90 | 0.06 |
| α-Linolenic Acid 18:3 ω3 | 0.30 | 0.30 | 0.02 |
| Arachidonic Acid 20:4 ω6 | 0.01 | No data | 0.08 |
| EPA 20:5 ω3 | 0.02 | No data | 0.27 |
| DHA 22:6 ω3 | 0.05 | No data | 1.19 |
| Total ω3 | 0.37 | 0.31 | 1.66 |
| Total ω6 | 1.31 | 2.90 | 0.23 |
| Total Monounsaturated Fats | 2.00 | 7.32 | 1.10 |
| Total Polyunsaturated Fats | 1.77 | 3.32 | 1.91 |
| Total Saturated Fats | 0.74 | 9.30 | 1.67 |
| Target Gene | NCBI Reference Sequence | Taqman Assay ID | Product Size |
|---|---|---|---|
| Gapdh | NM_017008.3 | 4352338E | 63 |
| Lepr | NM_012596 | Rn01433205_m1 | 94 |
| Npy | NM_012614.2 | Rn00561681_m1 | 63 |
| Agrp | NM_033650.1 | Rn01431703_g1 | 67 |
| Pomc | NM_139326.2 | Rn00595020_m1 | 92 |
| Ghsr | NM_032075.3 | Rn00821417_m1 | 61 |
| Pre-Conception Diet | Chow | HFSD | |||
|---|---|---|---|---|---|
| Post-Conception Diet | Chow | ω-3 | HFSD | Chow | ω-3 |
| Leptin (ng/mL) | 28.4 ± 3.9 | 48.0 ± 4.2 | 46.6 ± 6.1 | 36.8 ± 5.9 | 69.8 ± 10.7 *,$ |
| Triglycerides (mg/dL) | 47.2 ± 5.3 # | 42.28 ± 3.6 # | 105.4 ± 12.3 | 41.8 ± 3.9 # | 46.6 ± 4.1 # |
| Insulin (ng/mL) | 3.3 ± 0.5 | 4.9 ± 0.8 | 4.7 ± 1.2 | 3.0 ± 0.4 | 4.3 ± 0.5 |
| Total ghrelin (pg/mL) | 2172.6 ± 272.6 | 1729.1 ± 197.5 | 2531.9 ± 311.1 | 2193.9 ± 197.5 | 1885.9 ± 168.9 |
| Acyl ghrelin (pg/mL) | 710.5 ± 114.2 | 769.6 ± 134.9 | 505.8 ± 81.1 | 638.5 ± 126.7 | 563.9 ± 87.4 |
| Des-acyl ghrelin (pg/mL) | 1462 ± 246.0 | 959.6 ± 119.9 # | 2026.0 ± 306.4 | 1387.0 ± 325.4 | 1247.0 ± 55.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xavier, S.; Gili, J.; McGowan, P.; Younesi, S.; Wright, P.F.A.; Walker, D.W.; Spencer, S.J.; Sominsky, L. High Maternal Omega-3 Supplementation Dysregulates Body Weight and Leptin in Newborn Male and Female Rats: Implications for Hypothalamic Developmental Programming. Nutrients 2021, 13, 89. https://doi.org/10.3390/nu13010089
Xavier S, Gili J, McGowan P, Younesi S, Wright PFA, Walker DW, Spencer SJ, Sominsky L. High Maternal Omega-3 Supplementation Dysregulates Body Weight and Leptin in Newborn Male and Female Rats: Implications for Hypothalamic Developmental Programming. Nutrients. 2021; 13(1):89. https://doi.org/10.3390/nu13010089
Chicago/Turabian StyleXavier, Soniya, Jasmine Gili, Peter McGowan, Simin Younesi, Paul F. A. Wright, David W. Walker, Sarah J. Spencer, and Luba Sominsky. 2021. "High Maternal Omega-3 Supplementation Dysregulates Body Weight and Leptin in Newborn Male and Female Rats: Implications for Hypothalamic Developmental Programming" Nutrients 13, no. 1: 89. https://doi.org/10.3390/nu13010089