
Hyperspectral Reflectance and Machine Learning Approaches for the Detection of Drought and Root–Knot Nematode Infestation in Cotton


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crop Husbandry
2.2. Treatment
2.3. Data Collection
2.3.1. Physiological and Shoot Biomass Traits
2.3.2. Leaf Hyperspectral Reflectance
2.4. Statistical Machine Learning
3. Results and Discussion
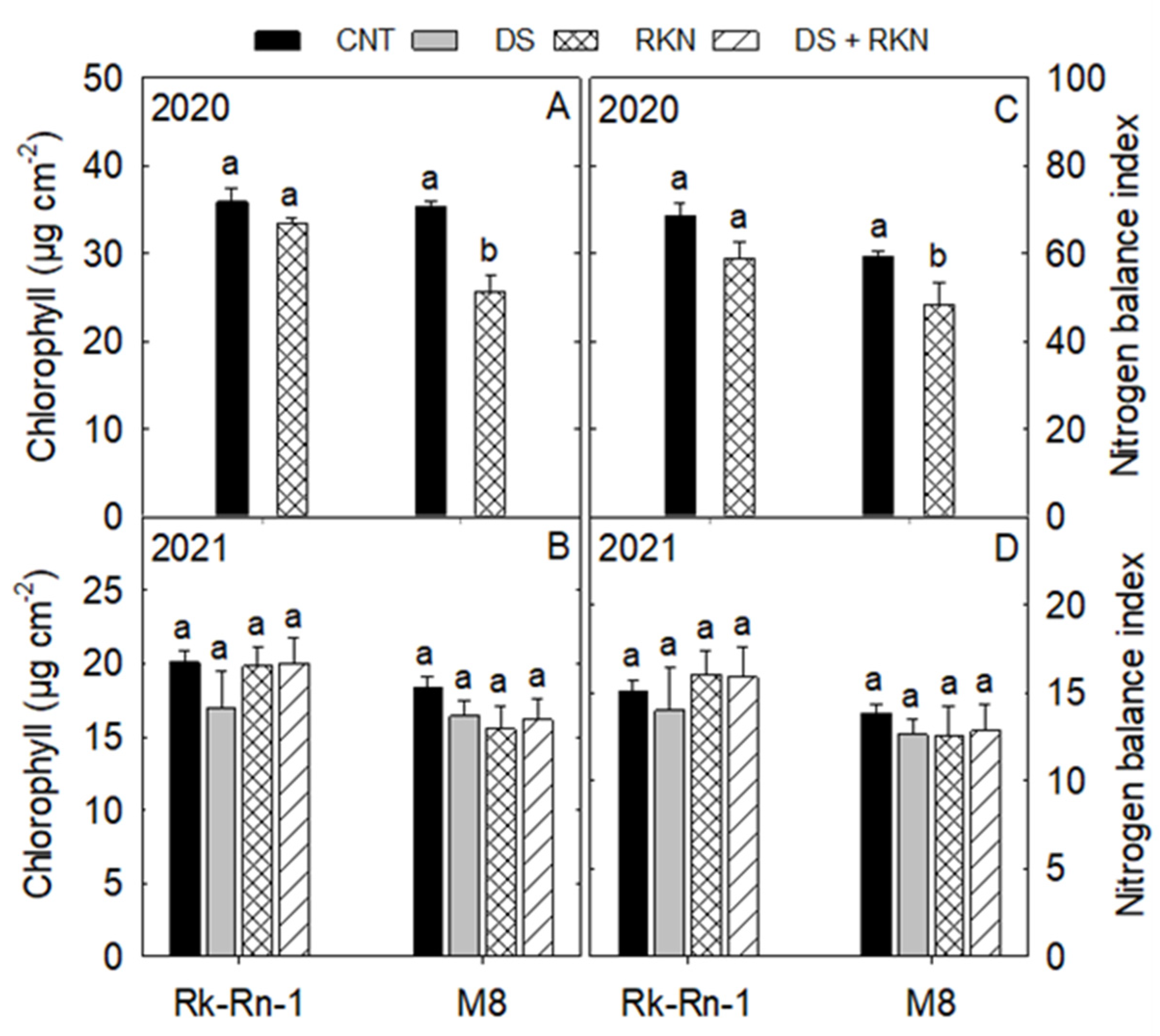
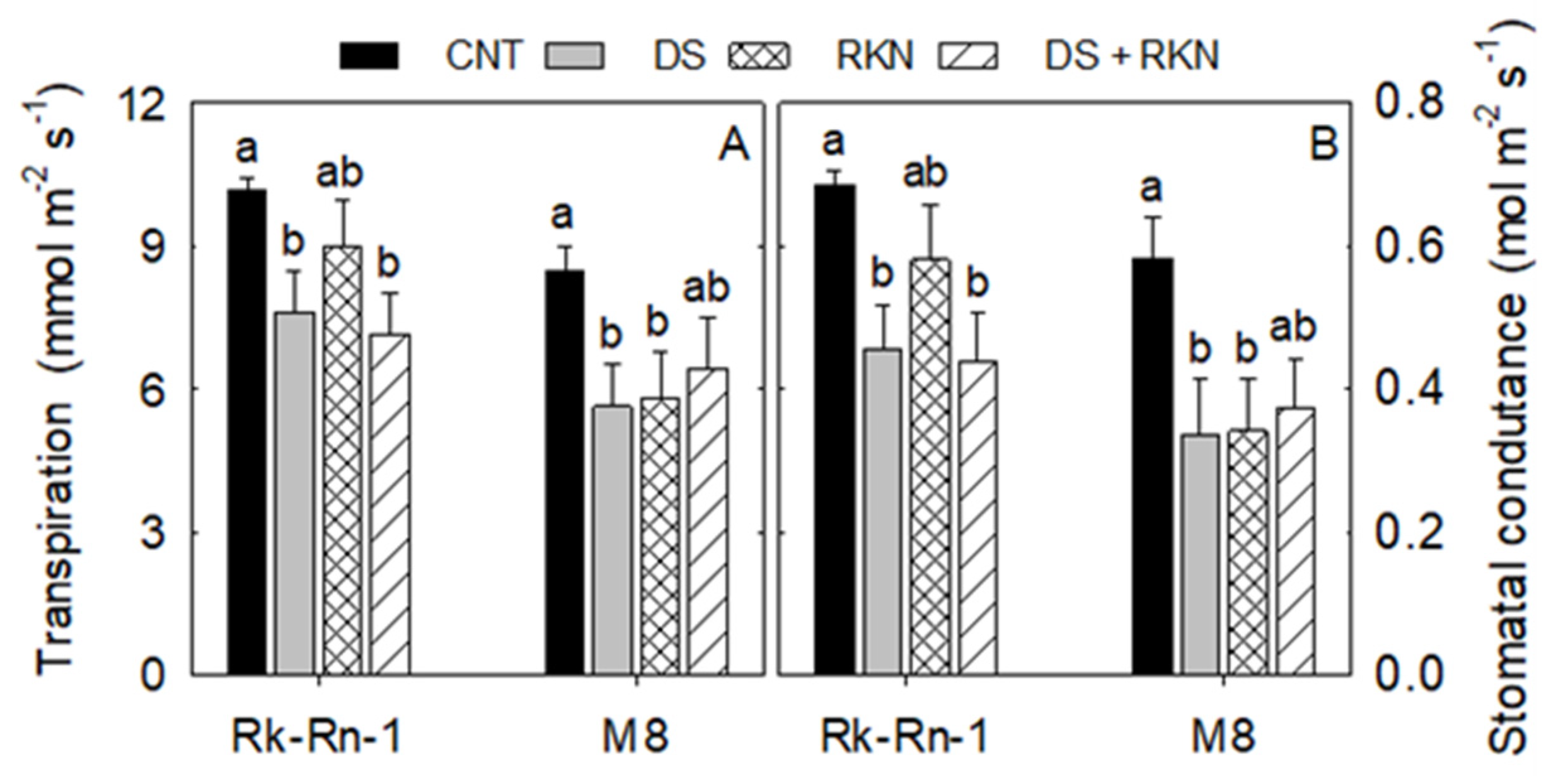
3.1. Physiological Traits
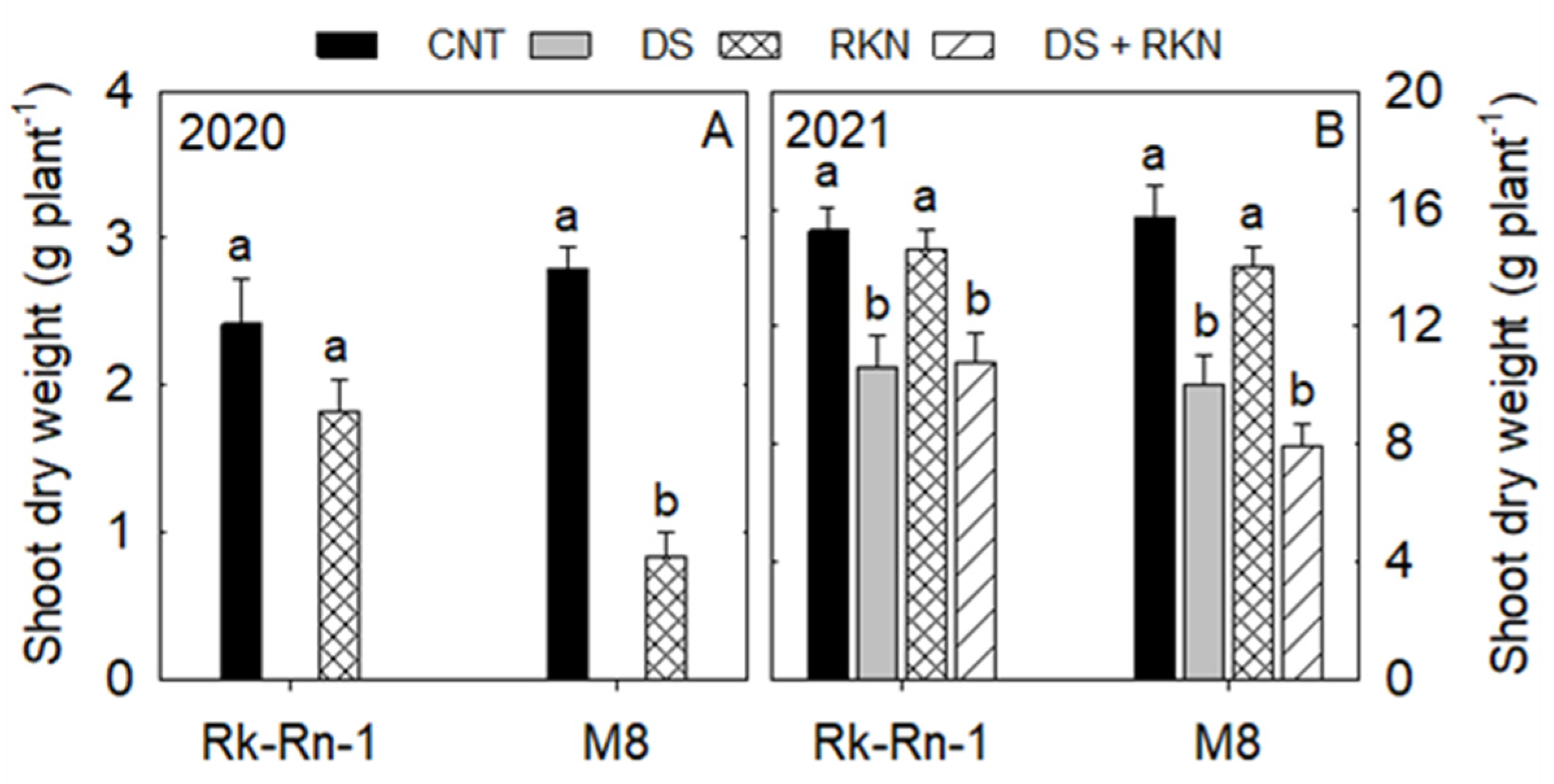
3.2. Biomass
3.3. Variation in Spectral Reflectance in Response to Stressors
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robinson, A.F. Reniform in U.S. Cotton: When, Where, Why, and Some Remedies. Annu. Rev. Phytopathol. 2007, 45, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, K.S.; Huang, P.; Lawrence, G.W.; Faske, T.; Overstreet, C.; Wheeler, T.; Young, H.; Kemerait, B.; Mehl, H. Nematode Research Report Cotton Varietal and Nematicide Responses in Nematode Soils. In Proceedings of the Beltwide nematode research and education committee 2014, San Antonio, TX, USA, 5–7 January 2015; p. 4. [Google Scholar]
- USDA. Cotton Price Statistics 2020–2021; Annual Report; United States Department of Agriculture: Memphis, TN, USA, 2021; Volume 102, pp. 1–21. Available online: https://www.ams.usda.gov/mnreports/cnaacps.pdf (accessed on 2 March 2022).
- Yeates, G.W.; Bongers, T.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding Habits in Soil Nematode Families and Genera—An Outline for Soil Ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.-M.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome Sequence of the Metazoan Plant-Parasitic Nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef]
- Wasson, A.P.; Ramsay, K.; Jones, M.G.K.; Mathesius, U. Differing Requirements for Flavonoids during the Formation of Lateral Roots, Nodules and Root Knot Nematode Galls in Medicago truncatula. New Phytol. 2009, 183, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, T.L.; Oosterhuis, D.M.; Wullschleger, S.D. Interaction of Meloidogyne Incognita and Water Stress in Two Cotton Cultivars. J. Nematol. 1991, 23, 462–467. [Google Scholar] [PubMed]
- Ynturi, P.; Jenkins, J.N.; McCarty, J.C., Jr.; Gutierrez, O.A.; Saha, S. Association of Root-Knot Nematode Resistance Genes with Simple Sequence Repeat Markers on Two Chromosomes in Cotton. Crop Sci. 2006, 46, 2670–2674. [Google Scholar] [CrossRef]
- Ralmi, N.H.A.A.; Khandaker, M.M.; Mat, N. Occurrence and Control of Root Knot Nematode in Crops: A Review. Aust. J. Crop Sci. 2016, 10, 1649–1654. [Google Scholar] [CrossRef]
- Pulavarty, A.; Egan, A.; Karpinska, A.; Horgan, K.; Kakouli-Duarte, T. Plant Parasitic Nematodes: A Review on Their Behaviour, Host Interaction, Management Approaches and Their Occurrence in Two Sites in the Republic of Ireland. Plants 2021, 10, 2352. [Google Scholar] [CrossRef]
- Barbary, A.; Djian-Caporalino, C.; Marteu, N.; Fazari, A.; Caromel, B.; Castagnone-Sereno, P.; Palloix, A. Plant Genetic Background Increasing the Efficiency and Durability of Major Resistance Genes to Root-Knot Nematodes Can Be Resolved into a Few Resistance QTLs. Front. Plant Sci. 2016, 7, 632. [Google Scholar] [CrossRef]
- Roberts, P.A. Current Status of the Availability, Development, and Use of Host Plant Resistance to Nematodes. J. Nematol. 1992, 24, 213–227. [Google Scholar]
- Sikder, M.M.; Vestergård, M. Impacts of Root Metabolites on Soil Nematodes. Front. Plant Sci. 2020, 10, 1792. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Cotton Incorporated Irrigation Management—Why Irrigate Cotton? Available online: https://www.cottoninc.com/cotton-production/ag-resources/irrigation-management/why-irrigate-cotton/ (accessed on 2 March 2022).
- McDonald, R.I.; Girvetz, E.H. Two Challenges for U.S. Irrigation Due to Climate Change: Increasing Irrigated Area in Wet States and Increasing Irrigation Rates in Dry States. PLoS ONE 2013, 8, e65589. [Google Scholar] [CrossRef]
- Qi, J.; Wang, J.; Gong, Z.; Zhou, J.-M. Apoplastic ROS Signaling in Plant Immunity. Curr. Opin. Plant Biol. 2017, 38, 92–100. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. Genetic Engineering for Modern Agriculture: Challenges and Perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, N.J.; Urwin, P.E. The Interaction of Plant Biotic and Abiotic Stresses: From Genes to the Field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed]
- Eisenback, J.D.; Dodge, D.J. Description of a Unique, Complex Feeding Socket Caused by the Putative Primitive Root-Knot Nematode, Meloidogyne kikuyensis. J. Nematol. 2012, 44, 148–152. [Google Scholar]
- Holbein, J.; Franke, R.B.; Marhavý, P.; Fujita, S.; Górecka, M.; Sobczak, M.; Geldner, N.; Schreiber, L.; Grundler, F.M.W.; Siddique, S. Root Endodermal Barrier System Contributes to Defence against Plant-Parasitic Cyst and Root-Knot Nematodes. Plant J. 2019, 100, 221–236. [Google Scholar] [CrossRef]
- Davis, R.F.; Earl, H.J.; Timper, P. Effect of Simultaneous Water Deficit Stress and Meloidogyne incognita Infection on Cotton Yield and Fiber Quality. J. Nematol. 2014, 46, 108–118. [Google Scholar]
- Kirkpatrick, T.L.; Van Iersel, M.W.; Oosterhuis, D.M. Influence of Meloidogyne incognita on the Water Relations of Cotton Grown in Microplots. J. Nematol. 1995, 27, 465–471. [Google Scholar]
- Reddy, K.R.; Hodges, H.F.; McKinion, J.M. Crop Modeling and Application: A Cotton Example. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 1997; ISBN 978-0-08-056372-5. [Google Scholar]
- Lu, P.; Davis, R.F.; Kemerait, R.C.; van Iersel, M.W.; Scherm, H. Physiological Effects of Meloidogyne Incognita Infection on Cotton Genotypes with Differing Levels of Resistance in the Greenhouse. J. Nematol. 2014, 46, 352–359. [Google Scholar] [PubMed]
- Carneiro, R.G.; Mazzafera, P.; Ferraz, L.C.C.B.; Muraoka, T.; Trivelin, P.C.O. Uptake and Translocation of Nitrogen, Phosphorus and Calcium in Soybean Infected with Meloidogyne incognita and M. Javanica. Fitopatol. Bras. 2002, 27, 141–150. [Google Scholar] [CrossRef]
- Walker, N.R.; Kirkpatrick, T.L.; Rothrock, C.S. Interaction between Meloidogyne incognita and Thielaviopsis basicola on Cotton (Gossypium hirsutum). J. Nematol. 1998, 30, 415–422. [Google Scholar]
- Bock, C.H.; Poole, G.H.; Parker, P.E.; Gottwald, T.R. Plant Disease Severity Estimated Visually, by Digital Photography and Image Analysis, and by Hyperspectral Imaging. Crit. Rev. Plant Sci. 2010, 29, 59–107. [Google Scholar] [CrossRef]
- Lowe, A.; Harrison, N.; French, A.P. Hyperspectral Image Analysis Techniques for the Detection and Classification of the Early Onset of Plant Disease and Stress. Plant Methods 2017, 13, 80. [Google Scholar] [CrossRef]
- Wong, C.Y.S.; Gamon, J.A. Three Causes of Variation in the Photochemical Reflectance Index (PRI) in Evergreen Conifers. New Phytol. 2015, 206, 187–195. [Google Scholar] [CrossRef]
- Sun, S.; Zhou, Y.; Chen, J.; Shi, J.; Zhao, H.; Zhao, H.; Song, W.; Zhang, M.; Cui, Y.; Dong, X.; et al. Extensive Intraspecific Gene Order and Gene Structural Variations between Mo17 and Other Maize Genomes. Nat. Genet. 2018, 50, 1289–1295. [Google Scholar] [CrossRef]
- Samiappan, S.; Bheemanahalli, R.; Zhou, M.; Brooks, J.; Wubben, M. Early Detection of Root-Knot Nematode (Meloidogyne incognita) Infestation in Cotton Using Hyperspectral Data. In Proceedings of the 2021 IEEE International Geoscience and Remote Sensing Symposium IGARSS, Brussels, Belgium, 11–16 July 2021; pp. 5849–5852. [Google Scholar]
- Grzybowski, M.; Wijewardane, N.K.; Atefi, A.; Ge, Y.; Schnable, J.C. Hyperspectral Reflectance-Based Phenotyping for Quantitative Genetics in Crops: Progress and Challenges. Plant Commun. 2021, 2, 100209. [Google Scholar] [CrossRef]
- Behmann, J.; Steinrücken, J.; Plümer, L. Detection of Early Plant Stress Responses in Hyperspectral Images. ISPRS J. Photogramm. Remote Sens. 2014, 93, 98–111. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a Green Channel in Remote Sensing of Global Vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Delalieux, S.; Somers, B.; Verstraeten, W.W.; van Aardt, J.A.N.; Keulemans, W.; Coppin, P. Hyperspectral Indices to Diagnose Leaf Biotic Stress of Apple Plants, Considering Leaf Phenology. Int. J. Remote Sens. 2009, 30, 1887–1912. [Google Scholar] [CrossRef]
- Melandri, G.; Thorp, K.R.; Broeckling, C.; Thompson, A.L.; Hinze, L.; Pauli, D. Assessing Drought and Heat Stress-Induced Changes in the Cotton Leaf Metabolome and Their Relationship with Hyperspectral Reflectance. Front. Plant Sci. 2021, 12, 751868. [Google Scholar] [CrossRef] [PubMed]
- Calzone, A.; Cotrozzi, L.; Lorenzini, G.; Nali, C.; Pellegrini, E. Hyperspectral Detection and Monitoring of Salt Stress in Pomegranate Cultivars. Agronomy 2021, 11, 1038. [Google Scholar] [CrossRef]
- Chen, J.J.; Zhen, S.; Sun, Y. Estimating Leaf Chlorophyll Content of Buffaloberry Using Normalized Difference Vegetation Index Sensors. HortTechnology 2021, 31, 297–303. [Google Scholar] [CrossRef]
- Prasad, S.; Kalluri, H.; Bruce, L.M.; Samiappan, S. Data Dependent Adaptation for Improved Classification of Hyperspectral Imagery. In Proceedings of the 2010 IEEE International Geoscience and Remote Sensing Symposium, Honolulu, HI, USA, 25–30 July 2010; pp. 68–71. [Google Scholar]
- Samiappan, S.; Prasad, S.; Bruce, L.M.; Robles, W. NASA’s Upcoming HyspIRI Mission—Precision Vegetation Mapping with Limited Ground Truth. In Proceedings of the 2010 IEEE International Geoscience and Remote Sensing Symposium, Honolulu, HI, USA, 25–30 July 2010; pp. 3744–3747. [Google Scholar]
- Melgani, F.; Bruzzone, L. Classification of Hyperspectral Remote Sensing Images with Support Vector Machines. IEEE Trans. Geosci. Remote Sens. 2004, 42, 1778–1790. [Google Scholar] [CrossRef]
- Cheriyadat, A.; Bruce, L.M. Why Principal Component Analysis Is Not an Appropriate Feature Extraction Method for Hyperspectral Data. In Proceedings of the IGARSS 2003, 2003 IEEE International Geoscience and Remote Sensing Symposium (IEEE Cat. No.03CH37477), Toulouse, France, 21–25 July 2003; Volume 6, pp. 3420–3422. [Google Scholar]
- Du, Q.; Younan, N.H. Dimensionality Reduction and Linear Discriminant Analysis for Hyperspectral Image Classification. In Proceedings of the Knowledge-Based Intelligent Information and Engineering Systems, Zagreb, Croatia, 3–5 September 2008; Lovrek, I., Howlett, R.J., Jain, L.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 392–399. [Google Scholar]
- Duda, R.O.; Hart, P.E.; Stork, G.E. Pattern Classification, 2nd ed.; Wiley Inter-Science: New York, NY, USA, 2000. [Google Scholar]
- Prasad, S.; Bruce, L.M. Overcoming the Small Sample Size Problem in Hyperspectral Classification and Detection Tasks. In Proceedings of the IGARSS 2008—2008 IEEE International Geoscience and Remote Sensing Symposium, Boston, MA, USA, 6–11 July 2008; Volume 5, pp. V-381–V-384. [Google Scholar]
- McCarty, J.C., Jr.; Jenkins, J.N.; Wubben, M.J.; Hayes, R.W.; Callahan, F.E.; Deng, D. Registration of Six Germplasm Lines of Cotton with Resistance to the Root-Knot and Reniform Nematodes. J. Plant Regist. 2017, 11, 168–171. [Google Scholar] [CrossRef]
- Hussey, R.; Barker, K. A Comparison of Methods of Collecting Inocula of Meloidogyne spp., Including a New Technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Foody, G.M. Status of Land Cover Classification Accuracy Assessment. Remote Sens. Environ. 2002, 80, 185–201. [Google Scholar] [CrossRef]
- da Silva, E.C.; Nogueira, R.J.; Silva, M.; Albuquerque, M. Drought Stress and Plant Nutrition. Plant Stress 2010, 5, 32–41. [Google Scholar]
- Giunta, F.; Motzo, R.; Deidda, M. SPAD Readings and Associated Leaf Traits in Durum Wheat, Barley and Triticale Cultivars. Euphytica 2002, 125, 197–205. [Google Scholar] [CrossRef]
- Nafady, N.A.; Sultan, R.; El-Zawahry, A.M.; Mostafa, Y.S.; Alamri, S.; Mostafa, R.G.; Hashem, M.; Hassan, E.A. Effective and Promising Strategy in Management of Tomato Root-Knot Nematodes by Trichoderma harzianum and Arbuscular Mycorrhizae. Agronomy 2022, 12, 315. [Google Scholar] [CrossRef]
- Ding, L.; Lu, Z.; Gao, L.; Guo, S.; Shen, Q. Is Nitrogen a Key Determinant of Water Transport and Photosynthesis in Higher Plants upon Drought Stress? Front. Plant Sci. 2018, 9, 1143. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wang, S.; Cao, B.; Cao, D.; Leng, G.; Li, H.; Yin, L.; Shan, L.; Deng, X. Genotypic Variation in Growth and Physiological Response to Drought Stress and Re-Watering Reveals the Critical Role of Recovery in Drought Adaptation in Maize Seedlings. Front. Plant Sci. 2016, 6, 1241. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, P.; Bheemanahalli, R.; Meyers, S.L.; Shankle, M.W.; Reddy, K.R. Drought, Low Nitrogen Stress, and Ultraviolet-B Radiation Effects on Growth, Development, and Physiology of Sweetpotato Cultivars during Early Season. Genes 2022, 13, 156. [Google Scholar] [CrossRef]
- Zhao, D.; Reddy, K.R.; Kakani, V.G.; Reddy, V.R. Nitrogen Deficiency Effects on Plant Growth, Leaf Photosynthesis, and Hyperspectral Reflectance Properties of Sorghum. Eur. J. Agron. 2005, 22, 391–403. [Google Scholar] [CrossRef]
- Collalti, A.; Tjoelker, M.G.; Hoch, G.; Mäkelä, A.; Guidolotti, G.; Heskel, M.; Petit, G.; Ryan, M.G.; Battipaglia, G.; Matteucci, G.; et al. Plant Respiration: Controlled by Photosynthesis or Biomass? Glob. Change Biol. 2020, 26, 1739–1753. [Google Scholar] [CrossRef]
- Zhang, H.; Sonnewald, U. Differences and Commonalities of Plant Responses to Single and Combined Stresses. Plant J. 2017, 90, 839–855. [Google Scholar] [CrossRef]
- Zhang, P.; Bonte, D.; De Deyn, G.B.; Vandegehuchte, M.L. Belowground Plant-Plant Signaling of Root Infection by Nematodes. Pedobiologia 2020, 83, 150688. [Google Scholar] [CrossRef]
- Zhang, P.; Bonte, D.; De Deyn, G.B.; Vandegehuchte, M.L. Leachates from Plants Recently Infected by Root-Feeding Nematodes Cause Increased Biomass Allocation to Roots in Neighbouring Plants. Sci. Rep. 2021, 11, 2347. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; Christie, P.; Li, X. Pre-Inoculation with Arbuscular Mycorrhizal Fungi Suppresses Root Knot Nematode (Meloidogyne incognita) on Cucumber (Cucumis sativus). Biol. Fertil. Soils 2008, 45, 205. [Google Scholar] [CrossRef]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Days after Stress | Class Accuracy | M8 | Rk-Rn-1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCA | SLDA | FLDA | PCA | SLDA | FLDA | ||||||||
| 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | ||
| 10 | Control | 96 | 96 | 100 | 92 | 80 | 68 | 100 | 100 | 100 | 88 | 72 | 52 |
| RKN | 96 | 92 | 96 | 100 | 68 | 32 | 96 | 100 | 96 | 92 | 52 | 68 | |
| OA | 96 | 94 | 98 | 96 | 74 | 50 | 98 | 100 | 98 | 90 | 62 | 60 | |
| Kappa | 0.92 | 0.88 | 0.96 | 0.92 | 0.48 | 0.0 | 0.96 | 1.0 | 0.96 | 0.80 | 0.24 | 0.20 | |
| 30 | Control | 95 | 100 | 95 | 95 | 95 | 57 | 96 | 100 | 98 | 92 | 72 | 4 |
| RKN | 96 | 96 | 100 | 100 | 64 | 72 | 100 | 96 | 100 | 88 | 80 | 76 | |
| OA | 96 | 98 | 98 | 94 | 78 | 65 | 98 | 98 | 98 | 90 | 76 | 40 | |
| Kappa | 0.91 | 0.96 | 0.96 | 0.87 | 0.57 | 0.29 | 0.96 | 0.96 | 0.96 | 0.92 | 0.52 | −0.20 | |
| 60 | Control | 87 | 87 | 93 | 93 | 53 | 40 | 100 | 96 | 96 | 100 | 84 | 80 |
| RKN | 100 | 100 | 100 | 100 | 84 | 84 | 96 | 96 | 96 | 96 | 88 | 80 | |
| OA | 95 | 95 | 98 | 98 | 73 | 68 | 98 | 96 | 96 | 98 | 86 | 80 | |
| Kappa | 0.89 | 0.89 | 0.89 | 0.95 | 0.39 | 0.26 | 0.96 | 0.92 | 0.92 | 0.96 | 0.72 | 0.80 | |
| Days after Stress | Class Accuracy | M8 | Rk-Rn-1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCA | SLDA | FLDA | PCA | SLDA | FLDA | |||||||||
| 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | |||
| 32 | Control | 96 | 96 | 96 | 98 | 96 | 98 | 96 | 98 | 94 | 100 | 100 | 100 | |
| RKN | 100 | 100 | 95 | 98 | 100 | 91 | 100 | 100 | 98 | 96 | 89 | 98 | ||
| DS | 100 | 94 | 83 | 85 | 100 | 96 | 94 | 98 | 92 | 96 | 98 | 94 | ||
| DS + RKN | 98 | 100 | 98 | 100 | 98 | 93 | 98 | 100 | 98 | 100 | 85 | 89 | ||
| OA | 99 | 96 | 93 | 90 | 99 | 97 | 96 | 98 | 95 | 95 | 92 | 92 | ||
| Kappa | 1.0 | 0.97 | 0.90 | 0.93 | 1.0 | 0.92 | 0.98 | 0.99 | 0.94 | 0.97 | 0.90 | 0.93 | ||
| Control | 98 | 98 | 100 | 96 | 100 | 100 | 98 | 94 | 100 | 96 | 100 | 100 | ||
| 39 | RKN | 98 | 98 | 100 | 100 | 94 | 94 | 100 | 100 | 100 | 100 | 91 | 92 | |
| DS | 100 | 100 | 94 | 89 | 98 | 98 | 98 | 98 | 94 | 96 | 96 | 92 | ||
| DS + RKN | 98 | 98 | 98 | 98 | 94 | 93 | 98 | 98 | 100 | 100 | 98 | 98 | ||
| OA | 99 | 98 | 98 | 95 | 97 | 95 | 99 | 97 | 98 | 97 | 96 | 94 | ||
| Kappa | 1.0 | 0.98 | 0.97 | 0.94 | 1.00 | 0.97 | 0.98 | 0.97 | 0.98 | 0.97 | 0.95 | 0.94 | ||
| Control | 98 | 98 | 96 | 100 | 100 | 100 | 98 | 98 | 96 | 98 | 100 | 100 | ||
| 46 | RKN | 100 | 98 | 80 | 83 | 94 | 96 | 98 | 98 | 100 | 100 | 88 | 94 | |
| DS | 99 | 100 | 100 | 100 | 94 | 95 | 98 | 98 | 91 | 86 | 98 | 98 | ||
| DS + RKN | 99 | 99 | 98 | 96 | 98 | 99 | 100 | 100 | 98 | 100 | 94 | 94 | ||
| OA | 99 | 98 | 94 | 96 | 96 | 93 | 99 | 98 | 96 | 94 | 95 | 95 | ||
| Kappa | 1.0 | 0.98 | 0.92 | 0.93 | 0.90 | 0.96 | 0.98 | 0.98 | 0.95 | 0.94 | 0.93 | 0.95 | ||
| Control | 98 | 96 | 96 | 94 | 100 | 100 | 98 | 98 | 98 | 94 | 100 | 100 | ||
| 53 | RKN | 98 | 98 | 100 | 100 | 92 | 89 | 100 | 98 | 100 | 100 | 89 | 89 | |
| DS | 98 | 98 | 82 | 94 | 98 | 96 | 98 | 97 | 94 | 94 | 96 | 94 | ||
| DS + RKN | 100 | 100 | 98 | 100 | 96 | 98 | 100 | 98 | 98 | 98 | 94 | 96 | ||
| OA | 99 | 97 | 93 | 97 | 97 | 93 | 99 | 98 | 98 | 95 | 95 | 94 | ||
| Kappa | 1.0 | 0.97 | 0.91 | 0.96 | 1.0 | 0.94 | 0.99 | 0.97 | 0.97 | 0.95 | 0.93 | 0.93 | ||
| Control | 98 | 98 | 94 | 94 | 93 | 96 | 98 | 98 | 96 | 98 | 98 | 100 | ||
| 72 | RKN | 100 | 98 | 97 | 88 | 91 | 94 | 100 | 98 | 97 | 100 | 93 | 94 | |
| DS | 88 | 98 | 80 | 94 | 93 | 96 | 88 | 98 | 78 | 86 | 89 | 96 | ||
| DS + RKN | 100 | 100 | 100 | 98 | 92 | 96 | 100 | 100 | 97 | 100 | 93 | 100 | ||
| OA | 96 | 97 | 92 | 93 | 92 | 88 | 96 | 99 | 91 | 96 | 93 | 98 | ||
| Kappa | 0.95 | 0.98 | 0.89 | 0.91 | 0.90 | 0.94 | 0.95 | 0.98 | 0.87 | 0.94 | 0.91 | 0.97 | ||
| Days after Stress/ Class Accuracy | PCA | SLDA | FLDA | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 32 | 39 | 46 | 53 | 72 | 32 | 39 | 46 | 53 | 72 | 32 | 39 | 46 | 53 | 72 | ||
| Wavelengths (350–1000 nm) | M8–DS + RKN | 92.6 | 98.0 | 98.0 | 95.2 | 97.8 | 94.3 | 100 | 96.2 | 96.2 | 97.8 | 100 | 100 | 100 | 100 | 100 |
| M8–RKN | 97.9 | 98.0 | 98.0 | 98.0 | 100 | 100 | 98.0 | 100 | 100 | 100 | 87.7 | 92.5 | 92.5 | 92.5 | 88.0 | |
| Rk-Rn-1–DS + RKN | 98.0 | 100 | 98.0 | 100 | 90.0 | 92.5 | 96.1 | 98.0 | 94.3 | 79.0 | 100 | 98.0 | 98.0 | 98.0 | 100 | |
| Rk-Rn-1–RKN | 98.0 | 98.0 | 100 | 98.0 | 100 | 98.0 | 96.1 | 96.0 | 98.0 | 97.4 | 100 | 100 | 100 | 98.0 | 97.8 | |
| M8–DS | 100 | 96.0 | 98.0 | 100 | 95.7 | 98.0 | 96.1 | 95.7 | 96.0 | 99.8 | 94.2 | 94.3 | 90.9 | 94.3 | 95.7 | |
| M8–control | 97.8 | 98.0 | 100.0 | 94.2 | 95.7 | 100 | 100 | 94.1 | 96.0 | 97.8 | 95.4 | 97.9 | 100.0 | 97.9 | 89.8 | |
| Rk-Rn-1–DS | 98.0 | 98.0 | 98.0 | 98.0 | 97.7 | 98.0 | 100 | 96.1 | 98.0 | 97.7 | 98.0 | 98.0 | 98.0 | 98.0 | 95.5 | |
| Rk-Rn-1–control | 98.0 | 98.0 | 98.0 | 98.0 | 97.8 | 98.0 | 100 | 98.0 | 100 | 97.8 | 96.2 | 98.0 | 98.0 | 98.0 | 100 | |
| OA | 98.0 | 98.0 | 98.5 | 97.8 | 96.7 | 97.2 | 98.3 | 96.8 | 97.3 | 95.0 | 96.2 | 97.3 | 97.0 | 97.0 | 95.6 | |
| Kappa | 0.97 | 0.98 | 0.98 | 0.97 | 0.96 | 0.97 | 0.98 | 0.96 | 0.97 | 0.94 | 0.96 | 0.97 | 0.97 | 0.97 | 0.95 | |
| Wavelengths (350–2500 nm) | M8–DS + RKN | 92.6 | 100 | 96.2 | 96.2 | 97.8 | 95.2 | 100 | 96.2 | 96.2 | 97.8 | 100 | 100 | 100 | 100 | 100 |
| M8–RKN | 97.8 | 100 | 97.9 | 98.0 | 100 | 100 | 98.0 | 100 | 100 | 100 | 87.7 | 94.2 | 92.5 | 92.5 | 88.2 | |
| Rk-Rn-1–DS + RKN | 98.0 | 100 | 96.1 | 100 | 88.2 | 89.3 | 96.0 | 98.0 | 94.3 | 83.3 | 100 | 98.0 | 98.0 | 98.0 | 100 | |
| Rk-Rn-1–RKN | 96.2 | 98.0 | 98.0 | 96.2 | 97.7 | 98.0 | 96.0 | 96.0 | 98.0 | 97.5 | 100 | 100 | 100 | 98.0 | 97.8 | |
| M8–DS | 100 | 94.3 | 100 | 100 | 100 | 98.0 | 96.0 | 95.7 | 100 | 97.8 | 94.2 | 94.3 | 90.9 | 94.3 | 97.8 | |
| M8–control | 93.8 | 100 | 100 | 98.0 | 95.7 | 97.7 | 100 | 94.1 | 100 | 100 | 95.4 | 97.9 | 100 | 95.9 | 93.6 | |
| Rk-Rn-1–DS | 100 | 98.0 | 96.1 | 98.0 | 97.8 | 98.0 | 98.0 | 96.1 | 98.0 | 97.8 | 98.0 | 98.0 | 98.0 | 98.0 | 93.3 | |
| Rk-Rn-1–control | 98.0 | 98.0 | 98.0 | 98.0 | 100 | 98.0 | 100 | 98.0 | 100 | 97.8 | 96.2 | 100 | 98.0 | 98.0 | 100 | |
| OA | 97.0 | 98.0 | 97.8 | 98.0 | 96.9 | 96.7 | 98.0 | 96.8 | 98.3 | 96.1 | 96.2 | 97.8 | 97.0 | 96.8 | 96.1 | |
| Kappa | 0.97 | 0.98 | 0.97 | 0.98 | 0.97 | 0.96 | 0.98 | 0.96 | 0.98 | 0.96 | 0.96 | 0.97 | 0.97 | 0.96 | 0.96 | |
| Days after Stress | Class Accuracy | M8 | Rk-Rn-1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PCA | SLDA | FLDA | PCA | SLDA | FLDA | ||||||||
| 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | 350–1000 nm | 350–2500 nm | ||
| 10 vs. 30 | Control | 39 | 52 | 70 | 78 | 65 | 35 | 56 | 40 | 96 | 36 | 32 | 48 |
| RKN | 100 | 100 | 100 | 100 | 100 | 80 | 100 | 100 | 100 | 100 | 100 | 100 | |
| OA | 70 | 77 | 85 | 90 | 83 | 58 | 78 | 70 | 98 | 68 | 66 | 74 | |
| Kappa | 0.44 | 0.53 | 0.70 | 0.90 | 0.66 | 0.15 | 0.56 | 0.40 | 0.96 | 0.36 | 0.32 | 0.48 | |
| 10 vs. 60 | Control | 44 | 55 | 60 | 30 | 25 | 30 | 48 | 36 | 88 | 80 | 24 | 40 |
| RKN | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 80 | |
| OA | 76 | 80 | 82 | 69 | 67 | 69 | 74 | 68 | 94 | 90 | 62 | 60 | |
| Kappa | 0.48 | 0.58 | 0.63 | 0.32 | 0.27 | 0.32 | 0.48 | 0.36 | 0.88 | 0.8 | 0.24 | 0.20 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramamoorthy, P.; Samiappan, S.; Wubben, M.J.; Brooks, J.P.; Shrestha, A.; Panda, R.M.; Reddy, K.R.; Bheemanahalli, R. Hyperspectral Reflectance and Machine Learning Approaches for the Detection of Drought and Root–Knot Nematode Infestation in Cotton. Remote Sens. 2022, 14, 4021. https://doi.org/10.3390/rs14164021
Ramamoorthy P, Samiappan S, Wubben MJ, Brooks JP, Shrestha A, Panda RM, Reddy KR, Bheemanahalli R. Hyperspectral Reflectance and Machine Learning Approaches for the Detection of Drought and Root–Knot Nematode Infestation in Cotton. Remote Sensing. 2022; 14(16):4021. https://doi.org/10.3390/rs14164021
Chicago/Turabian StyleRamamoorthy, Purushothaman, Sathishkumar Samiappan, Martin J. Wubben, John P. Brooks, Amrit Shrestha, Rajendra Mohan Panda, K. Raja Reddy, and Raju Bheemanahalli. 2022. "Hyperspectral Reflectance and Machine Learning Approaches for the Detection of Drought and Root–Knot Nematode Infestation in Cotton" Remote Sensing 14, no. 16: 4021. https://doi.org/10.3390/rs14164021