
Species-Specific Secondary Metabolites from Primula veris subsp. veris Obtained In Vitro Adventitious Root Cultures: An Alternative for Sustainable Production
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
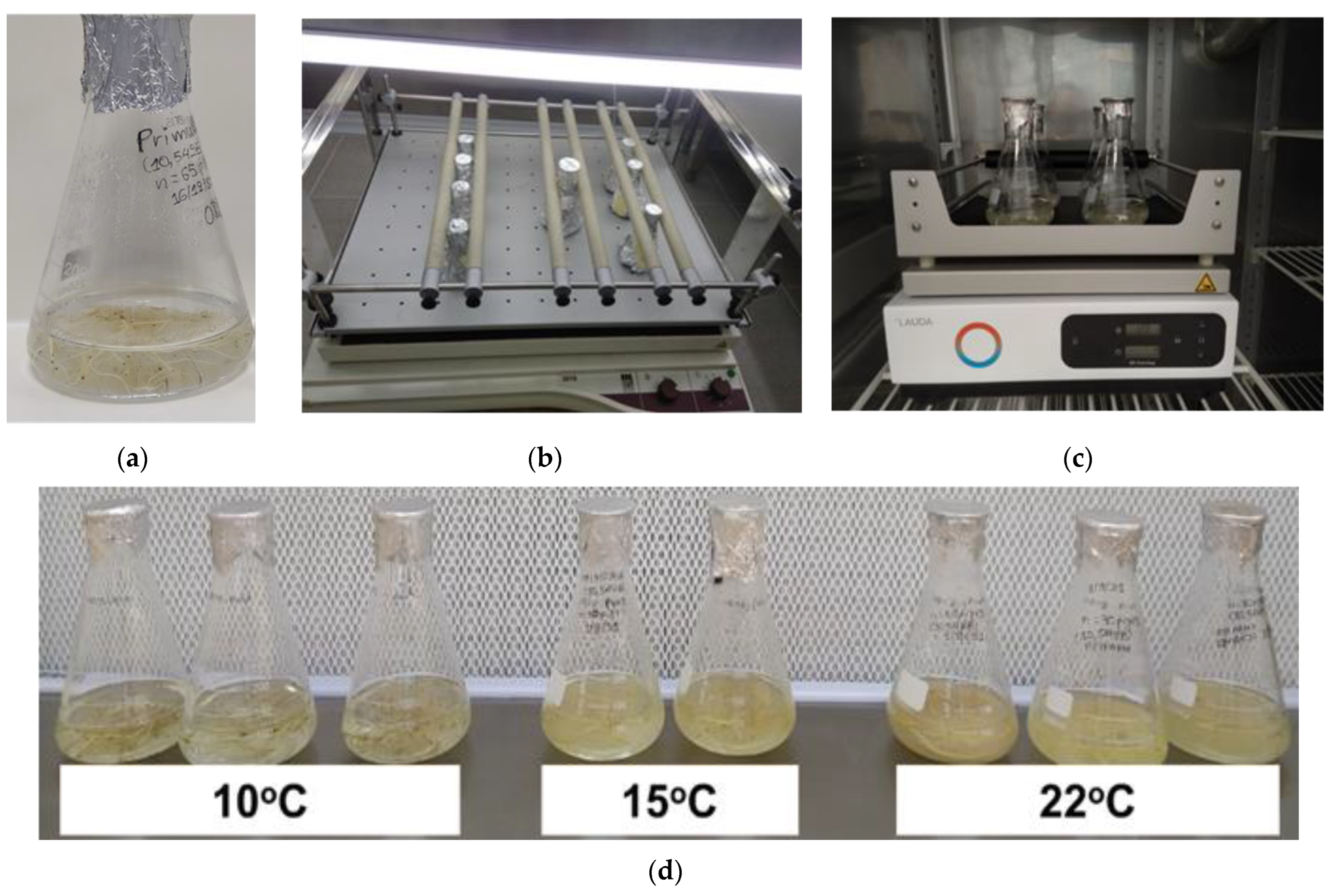
2.1. Plant Material and In Vitro Culture Conditions
2.2. Effect of Temperature on Root Biomass and Species-Specific Secondary Metabolites in Adventitious Root In Vitro Production
2.3. Comparison of Roots Secondary Metabolites Produced In Vivo Versus In Vitro
2.4. Extraction of Flavonoids and Triterpene Saponins
2.5. Ultra Performance Liquid Chromatography–Mass Spectrometry (UPLC-MS/MS, MRM) Analyses
2.6. Statistical Analysis
3. Results
3.1. Effect of Temperature on In Vitro Development and in Species-Specific Secondary Metabolites’ Production
3.2. Comparison of Species-Specific Secondary Metabolites of Roots Produced In Vivo and In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Budzianowski, J.; Morozowska, M.; Wesołowska, M. Lipophilic flavones of Primula veris L. from field cultivation and in vitro cultures. Phytochemistry 2005, 66, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Budzianowski, J.; Morozowska, M.; Wesołowska, M. Comparative study of lipophilic flavones in Primula veris L. from natural localities, field cultivation and in vitro culture. In Proceedings of the Application of Chromatographic Methods in Phytochemical and Biomedical Analysis, Abstracts of the 5th International Symposium on Chromatography of Natural Products (ISCNP), Lublin, Poland, 19–22 June 2006; p. 66. [Google Scholar]
- Neubauer, N.; März, R.W. Placebo-controlled, randomized double-blind clinical trial with Sinupret sugar coated tablets on the basis of a therapy with antibiotics and decongestant nasal drops in acute sinusitis. Phytomedicine 1994, 1, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Strzelecka, H.; Kowalski, J. Encyklopedia Zielarstwa I Ziołolecznictwa; PWN: Warszawa, Poland, 2000; p. 429. [Google Scholar]
- März, R.W. Bronchitis—Neue Erkenntnisse zu Wirkungen und Wirksamkeit von Arzneipflanzen; Karger: Basel, Switzerland, 1998. [Google Scholar]
- EMA/HMPC/104095/2012 Committee on Herbal Medicinal Products (HMPC) Community herbal monograph on Primula veris L. and/or Primula elatior (L.) Hill, radix. European Medicines Agency, 2013. Available online: https://www.ema.europa.eu›herbal-monograph (accessed on 25 October 2022).
- Wichtl, M.; Bisset, N.G. Herbal Drugs and Phytopharmaceuticals; CRC Press: Stuttgart, Germany, 1994. [Google Scholar]
- Lindborg, R.; Cousins, S.A.; Eriksson, O. Plant species response to land use change: Campanula rotundifolia, Primula veris and Rhinanthus minor. Ecography 2005, 28, 29–36. [Google Scholar] [CrossRef]
- El Morchid, M.M.; Torres Londoño, P.; Papagiannopoulos, M.; Gobbo-Neto, L.; Müller, C. Variation in flavonoid pattern in leaves and flowers of Primula veris of different origin and impact of UV-B. Biochem. Syst. Ecol. 2014, 53, 81–88. [Google Scholar] [CrossRef]
- Müller, A.; Ganzera, M.; Stuppner, H. Analysis of phenolic glycosides and saponins in Primula elatior and Primula veris (primula root) by liquid chromatography, evaporative light scattering detection and mass spectrometry. J. Chromatogr. A 2006, 1112, 218–223. [Google Scholar] [CrossRef]
- Draxler, L.; Göhler, I.; Franz, C. Cultivation and breeding of cowslip (Primula veris) for producing medicinal drugs. Rev. Fitoter. 2002, 2, 327. [Google Scholar]
- Morozowska, M. Some factors affecting viability, dormancy and germination of Primula veris (L.) seeds. Folia. Hortic. 2002, 14, 243–253. [Google Scholar]
- Morozowska, M.; Wesołowska, M. In vitro clonal propagation of Primula veris L. and preliminary phytochemical analysis. Acta Biol. Cracov. Ser. Bot. 2004, 46, 169–175. [Google Scholar]
- Sudha, C.G.; Seeni, S. Establishment and analysis of fast-growing normal root culture of Decalepis arayalpathra, a rare endemic medicinal plant. Curr. Sci. 2001, 81, 371–374. [Google Scholar]
- Paek, K.Y.; Murthy, H.N.; Hahn, E.J.; Zhong, J.J. Large scale culture of ginseng adventitious roots for production of ginsenosides. In Biotechnology in China I. Advances in Biochemical Engineering/Biotechnology; Zhong, J.J., Bai, F.W., Zhang, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 113, pp. 151–176. [Google Scholar] [CrossRef]
- Baque, M.A.; Elgirban, A.; Lee, E.J.; Paek, K.Y. Sucrose regulated enhanced induction of anthraquinone, phenolics, flavonoids biosynthesis and activities of antioxidant enzymes in adventitious root suspension cultures of Morinda citrifolia (L.). Acta Physiol. Plant. 2012, 34, 405–415. [Google Scholar] [CrossRef]
- Bais, H.P.; Loyola-Vargas, V.M.; Flores, H.E.; Vivanco, J.M. Root-specific metabolism: The biology and biochemistry of underground organs. In Vitro Cell Dev. Biol. Plant 2001, 37, 730–741. [Google Scholar] [CrossRef]
- Bhamarapravati, S.; Juthapruth, S.; Mahachai, W.; Mahady, G. Antibacterial activity of Boesenbergia rotunda (L.) Mansf. and Myristica fragrans Houtt. against Helicobacter pylori. Songklanakarin J. Sci. Technol. 2006, 28, 157–163. [Google Scholar]
- Jimenez-Garcia, S.N.; Vazquez-Cruz, M.A.; Guevara-Gonzale, R.G.; Torres-Pacheco, I.; Cruz-Hernandez, A.; Feregrino-Perez, A.A. Current approaches for enhanced expression of secondary metabolites as bioactive compounds in plants for agronomic and human health purposes—A review. Pol. J. Food Nutr. Sci. 2013, 63, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Nandagopal, K.; Halder, M.; Dash, B.; Nayak, S.; Jha, S. Biotechnological approaches for production of anti-cancerous compounds resveratrol, podophyllotoxin and zerumbone. Curr. Med. Chem. 2018, 25, 4693–4717. [Google Scholar] [CrossRef]
- Kim, Y.S.; Hahn, E.J.; Murthy, H.; Paek, K.Y. Adventitious root growth and ginsenoside accumulation in Panax ginseng cultures as affected by methyl jasmonate. Biotechnol. Lett. 2004, 26, 1619–1622. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Sarropoulou, V.; Krigas, N.; Maloupa, E. In vitro propagation of Primula veris L. subsp. veris (Primulaceae): A valuable medicinal plant with ornamental potential. Int. J. Bot. Stud. 2020, 5, 532–539. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Ionkova, I. Optimization of flavonoid production in cell cultures of Astragalus missouriensis Nutt. (Fabaceae). Phcog. Mag. 2009, 4, 92–97. [Google Scholar]
- Luczkiewicz, M.; Kokotkiewicz, A.; Glod, D. Plant growth regulators affect biosynthesis and accumulation profile of isoflavone phytoestrogens in high-productive in vitro cultures of Genista tinctoria. Plant Cell Tiss Organ Cult. 2014, 118, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Babich, O.; Sukhikh, S.; Pungin, A.; Astahova, L.; Chupakhin, E.; Belova, D.; Prosekov, A.; Ivanova, S. Evaluation of the conditions for the cultivation of callus cultures of Hyssopus officinalis regarding the yield of polyphenolic compounds. Plants 2021, 10, 915. [Google Scholar] [CrossRef]
- Shen, Y.; Jin, L.; Xiao, P.; Lu, Y.; Bao, J.S. Total phenolics, flavonoids, antioxidant capacity in rice grain and their relations to grain color, size and weight. J. Cereal. Sci. 2009, 49, 106–111. [Google Scholar] [CrossRef]
- Masoumian, M.; Arbakariya, A.; Syahida, A.; Maziah, M. Effect of precursors on flavonoid production by Hydrocotyle bonariensis callus tissues. Afr. J. Biotechnol. 2011, 10, 6021–6029. [Google Scholar] [CrossRef]
- Mohammadrezakhani, S.; Hajilou, J.; Rezanejad, F.; Zaare-Nahandi, F. Assessment of exogenous application of proline on antioxidant compounds in three Citrus species under low temperature stress. J. Plant Interact. 2019, 14, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Blinstrubiene, A.; Burbulis, N.; Juškevičiūte, N.; Vaitkevičiene, N.; Žūkiene, R. Effect of growth regulators on Stevia rebaudiana Bertoni callus genesis and influence of auxin and proline to steviol glycosides, phenols, flavonoids accumulation, and antioxidant activity in vitro. Molecules 2020, 25, 2759. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, Z.; Karimi, N.; Modarresi, M.; Mollayi, S. Enhancement of compatible solute and secondary metabolites production in Plantago ovata Forsk. by salinity stress. J. Med. Plants Res. 2012, 6, 3495–3500. [Google Scholar] [CrossRef]
- Al-Jibouri, A.M.J.; Abd, A.S.; Majeed, D.M.; Ismail, E.N. Influence of abiotic elicitors on accumulation of thymol in callus cultures of Origanum vulgare L. J. Life Sci. 2012, 6, 1094–1099. [Google Scholar]
- Al-Jibouri, A.M.J.; Al-Samarraei, K.W.; Abd, A.S.; Mageed, D.M.; Ali, A.A. Alkaloids production from callus of Hyoscyamus niger L. in vitro. J. Life Sci. 2012, 6, 874–882. [Google Scholar]
- Al-Jibouri, A.M.J.; Abed, A.S.; Ali, A.J.A.; Majeed, D.M. Improvement of phenols production by amino acids in callus cultures of Verbascum thapsus L. Am. J. Plant Sci. 2016, 7, 84–91. [Google Scholar] [CrossRef]
- Salehi, M.; Moieni, A.; Safaie, N. A novel medium for enhancing callus growth of hazel (Corylus avellana L.). Sci. Rep. 2017, 7, 15598. [Google Scholar] [CrossRef] [Green Version]
- Saed-Moucheshi, A.; Mozafari, A.A. Alternate gene expression profiling of monoterpenes in Hymenocrater longiflorus as a novel pharmaceutical plant under water deficit. Sci. Rep. 2022, 12, 4084. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant. 2008, 132, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Sarrou, E.; Martens, S.; Chatzopoulou, P. Metabolite profiling and antioxidative activity of sage (Salvia fruticosa Mill.) under the influence of genotype and harvesting period. Ind. Crops Prod. 2016, 94, 240–250. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Masuero, D.; Gasperotti, M.; Franceschi, P.; Caputi, L.; Viola, R.; Mattivi, F. A versatile targeted metabolomics method for the rapid quantification of multiple classes of phenolics in fruits and beverages. J. Agric. Food Chem. 2012, 60, 8831–8840. [Google Scholar] [CrossRef]
- Lopes, S.M.S.; Francisco, M.G.; Higashi, B.; Almeida, R.T.R.; Krausová, G.; Pilau, E.J.; Gonçalves, J.E.; Gonçalves, R.A.C.; Oliveira, A.J.B. Chemical characterization and prebiotic activity of fructo-oligosaccharides from Stevia rebaudiana (Bertoni) roots and in vitro adventitious root cultures. Carbohydr. Polym. 2016, 152, 718–725. [Google Scholar] [CrossRef]
- Bączek, K.; Przybył, J.L.; Mirgos, M.; Kosakowska, O.; Szymborska-Sandhu, I.; Węglarz, Z. Phenolics in Primula veris L. and Primula elatior (L.) Hill raw materials. Int. J. Anal Chem. 2017, 2017, 2871579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansel, R.; Keller, K.; Rimpler, H.; Schneider, G. Hagers Handbuch der Pharmazeutischen Praxis: Band 6; Springer: Berlin, Heidelberg, Germany, 1994. [Google Scholar]
- Apel, L.; Kammerer, D.R.; Stintzing, F.C.; Spring, O. Comparative metabolite profiling of triterpenoid saponins and flavonoids in flower color mutations of Primula veris L. Int. J. Mol. Sci. 2017, 18, 153. [Google Scholar] [CrossRef]
- Morozowka, M.; Krzakowa, M. Genetic variation of Primula veris in natural and cultivated populations. Acta Biol. Cracov. Bot. 2003, 45, 177–182. [Google Scholar]
- Palazón, J.; Navarro-Ocaña, A.; Hernandez-Vazquez, L.; Mirjalili, M.H. Application of metabolic engineering to the production of scopolamine. Molecules 2008, 13, 1722–1742. [Google Scholar] [CrossRef] [Green Version]
- Kamada, H.; Okamura, N.; Satake, M.; Harada, H.; Shimomura, K. Alkaloid production by hairy root cultures in Atropa belladonna. Plant Cell Rep. 1986, 5, 239–242. [Google Scholar] [CrossRef]
- Morozowska, M. Vegetative development, flowering, fruiting and saponin content in cultivated cowslip (Primula veris L.) plants. Herba Pol. 2004, 50, 28–35. [Google Scholar] [CrossRef]
- Shen, H.; Tang, Y.; Muraoka, H.; Washitani, I. Characteristics of leaf photosynthesis and simulated individual carbon budget in Primula nutans under contrasting light and temperature conditions. J. Plant Res. 2008, 121, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Morison, J.I.L.; Lawlor, D.W. Interactions between increasing CO2 concentration and temperature on plant growth. Plant Cell Environ. 1999, 22, 659–682. [Google Scholar] [CrossRef] [Green Version]
- Jochum, G.M.; Mudge, K.G.; Thomas, R.B. Elevated temperatures increase leaf senescence and root secondary metabolite concentrations in the understory herb Panax quinquefolius (Araliaceae). Am. J. Bot. 2007, 94, 819–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.H.; Chakrabarty, D.; Lee, E.J.; Paek, K.Y. Production of adventitious roots and secondary metabolites by Hypericum perforatum L. in a bioreactor. Bioresour. Technol. 2010, 101, 4708–4716. [Google Scholar] [CrossRef]
- Okrslar, V.; Plaper, I.; Kovac, M.; Erjavec, A.; Obermajer, T.; Rebec, A.; Ravnikar, M.; Žel, J. Saponins in tissue culture of Primula veris L. Vitr. Cell Dev. Biol. Plant 2007, 43, 644–651. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Quantifier Ion | Qualifier Ion | |||||||
|---|---|---|---|---|---|---|---|---|
| Compound | Rt (min) | ES | Cone Voltage (V) | Q1 (m/z) | Collision Energy (V) | Q2 (m/z) | Collision Energy (V) | Q2 (m/z) |
| Primeverin | 4.1 | - | 22 | 475.2234 | 6 | 293.076 | 24 | 181.027 |
| Primulaverin | 3.82 | - | 22 | 475.16 | 6 | 293.08 | 44 | 166.013 |
| Primulic acid I | 10.23 | - | 80 | 1103.649 | 58 | 455.412 | 38 | 205.069 |
| Primulic acid II | 10.01 | - | 84 | 1235.713 | 64 | 100.947 | 78 | 455.403 |
| Secondary Metabolites | Content (mg/g DW) and Yield (mg/100 mL Medium/250 mL Flask) In Vitro | Content (mg/g DW) and Yield (mg/2.5 L Pot) In Vivo | |||
|---|---|---|---|---|---|
| T10 | T15 | T22 | p-Value | ||
| Primulaverin content | 8.70 ± 0.83 a | 9.93 ± 0.66 a | 9.71 ± 0.38 a | 0.420 ns | 10.23 ± 0.80 |
| Primeverin content | 0.03 ± 0.02 b | 0.10 ± 0.01 a | 0.09 ± 0.01 a | 0.048 * | 0.28 ± 0.03 |
| Cumulative flavonoids | 8.73 ± 0.85 a | 10.03 ± 0.67 a | 9.80 ± 0.38 a | 0.398 ns | 10.51 ± 0.82 |
| Primulic acid I content | 2.85 ± 0.31 c | 4.14 ± 0.05 b | 6.09 ± 0.20 a | 0.000 *** | 17.01 ± 0.36 |
| Primulic acid II content | 0.20 ± 0.03 b | 0.53 ± 0.01 a | 0.51 ± 0.06 a | 0.045 * | 0.09 ± 0.01 |
| Cumulative saponins content | 3.05 ± 0.33 c | 4.67 ± 0.03 b | 6.60 ± 0.40 a | 0.000 *** | 17.10 ± 0.52 |
| Primulaverin yield | 4.09 ± 0.51 c | 7.19 ± 0.76 ab | 10.04 ± 2.19 a | 0.049 * | 33.55 ± 2.85 |
| Primeverin yield | 0.01 ± 0.01 b | 0.07 ± 0.01 a | 0.09 ± 0.02 a | 0.018 * | 0.92 ± 0.13 |
| Cumulative flavonoids yield | 4.10 ± 0.52 c | 7.26 ± 0.77 ab | 10.13 ± 2.21 a | 0.038 * | 34.47 ± 0.67 |
| Primulic acid I yield | 1.33 ± 0.14 c | 2.98 ± 0.09 b | 6.28 ± 1.31 a | 0.010 * | 55.79 ± 1.27 |
| Primulic acid II yield | 0.10 ± 0.02 b | 0.38 ± 0.02 a | 0.59 ± 0.38 a | 0.045 * | 0.30 ± 0.02 |
| Cumulative saponins yield | 1.42 ± 0.15 c | 3.36 ± 0.11 b | 6.87 ± 1.66 a | 0.019 * | 56.09 ± 1.96 |
| Biomass | In Vitro | In Vivo | |||
| T10 | T15 | T22 | p-Value | ||
| Total fresh weight (g) | 2.38 ± 0.08 c | 4.09 ± 0.10 b | 6.04 ± 0.73 a | 0.003 ** | 18.40 ± 0.81 |
| Total dry weight (g) | 0.47 ± 0.02 b | 0.72 ± 0.03 ab | 1.02 ± 0.18 a | 0.029 * | 3.28 ± 0.13 |
| Fresh weight/dry weight ratio | 5.10 ± 0.08 a | 5.68 ± 0.08 a | 6.05 ± 0.46 a | 0.123 ns | 5.61 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarropoulou, V.; Sarrou, E.; Angeli, A.; Martens, S.; Maloupa, E.; Grigoriadou, K. Species-Specific Secondary Metabolites from Primula veris subsp. veris Obtained In Vitro Adventitious Root Cultures: An Alternative for Sustainable Production. Sustainability 2023, 15, 2452. https://doi.org/10.3390/su15032452
Sarropoulou V, Sarrou E, Angeli A, Martens S, Maloupa E, Grigoriadou K. Species-Specific Secondary Metabolites from Primula veris subsp. veris Obtained In Vitro Adventitious Root Cultures: An Alternative for Sustainable Production. Sustainability. 2023; 15(3):2452. https://doi.org/10.3390/su15032452
Chicago/Turabian StyleSarropoulou, Virginia, Eirini Sarrou, Andrea Angeli, Stefan Martens, Eleni Maloupa, and Katerina Grigoriadou. 2023. "Species-Specific Secondary Metabolites from Primula veris subsp. veris Obtained In Vitro Adventitious Root Cultures: An Alternative for Sustainable Production" Sustainability 15, no. 3: 2452. https://doi.org/10.3390/su15032452