4.1. Forage Yield and Composition
The DM production in the “control” grass was similar to the averages of 2.1 to 2.5 tons/ha obtained at 28 and 33 days of growth during the dry season of tropical regions in Mexico and Colombia [
43,
44]. The forage yield analysis indicated that, in the “bank” physiographic position under floodplain savannah conditions, the biomass production of the native species
P. plicatulum (on a fresh and dry matter basis) was comparable to that of the “control” grass until 50 days of age. This behavior was largely attributed to the similar morphological characteristics between both plants and their bunch growth habit, which provided them with similar growth traits and propagation mechanisms [
17].
The height measured in the “control” grass was greater than the 61.93 cm and 64.92 cm values reported in other studies at 56 and 60 days of age [
17,
45]. However, it was similar to the 100 cm to 110 cm range observed at 51 and 54 days of growth [
46]. Different height measurements can be due to differences in the soil’s physicochemical properties and climatic variables, such as rainfall and temperature [
45]. The control and
P. plicatulum species were the tallest plants among the studied grasses, which can also be attributed to the bunch growth habit of these species, which easily outperformed the other grasses with stoloniferous, rhizomatous, or decumbent growths patterns [
17].
With respect to the DM concentrations, in the “control” grass, the values observed were similar to those found in the Colombian Caribbean and in different
Brachiaria accessions [
47,
48]. In this study, the DM increased over time in all of the evaluated plants, which was expected, as this variable tended to increase as the plant reached maturity [
49,
50]. Grasses such as
A. purpussi and
P. plicatulum were similar to the “control” grass during the first 40 days (20% to 25.5%), while
Paspalum sp. and
A. compresus presented higher values during the same period (28.1% to 34%). The high DM content observed during the early to medium growth phase of these plants suggests a major proportion in the cell content and a lower fiber fraction. This suggests that potentially digestible fractions are conserved for a longer time, which is useful in livestock systems with long grazing periods [
51].
The CP content of the evaluated grasses ranged between 4.3% and 10.2% during the experimental period, being comparable to the reports for other grasses grown under tropical environments in Latin American regions [
14,
52,
53]. The CP concentration in the “control” grass was similar to the 7.7% average [
43] and the 5.8% to 7.5% range obtained in 24 accessions of
Brachiaria spp. [
47]. In general, native grasses were found to be similar to or higher than the “control” plant in terms of CP concentration. These results suggest that, in the evaluated ecosystem, native plants presented physiological mechanisms of soil nitrogen extraction and utilization similar or superior to those of the control grass. In this way, native plants could also be considered as an alternative protein source under flooded savannah conditions. These results are interesting, as protein serves as an indicator of the nutritional potential of pastures [
16] and is considered a limiting nutrient, especially in highly fibrous diets, such as those commonly found in tropical environments [
54].
A. purpussi,
Paspalum sp., and the “control” grass exhibited a drastic reduction in the CP content until 40 days, when it then stabilized. The reduction in the CP concentration over time is an expected behavior, as the metabolic activity of plants is reduced as they reach maturity [
17,
51,
52]. Similarly, the stability achieved after 40 days suggests that the CP contents in these grasses were preserved for up to 50 days, and higher losses could be expected after this age. In
A. compresus, the CP content presented a growing pattern during the experimental period, while in
P. plicatulum, the highest CP value was found at 40 days, which then decreased until day 50. This suggests that
A. compresus and
P. plicatulum have slower maturation rates than other grasses, allowing them to use nitrogen efficiently for longer periods during the early growth phases [
45]. This effect could be attributed to their adaptability to the floodable savannah agroecosystem.
The estimated ash levels (3.2% to 8.7%) were within the range found in other improved pastures commonly used in animal nutrition [
47,
52,
55].
P. plicatulum, Paspalum sp., and
A. purpussi exhibited higher ash concentrations than the control plant during the experimental period. These results confirm the differential abilities of native plants to extract minerals, even in acidic soil, with low exchangeable base and high microelement concentrations (Mn, Zn, B, and Fe), similar to those found in the evaluated soil. These results suggest the necessity to evaluate the individual mineral profile of the ash fraction in the studied native plants, because, under these conditions, plants can absorb these elements in excess, producing toxicity in the same plant and in the animals when they are consumed [
56].
The ash reduction observed in
A. compresus and the “control” grass during the experimental period could be related to changes in mineral requirements according to the phenological state [
51]. Similarly, the soil type and other climatic variables, such as precipitation and temperature, can influence the availability of elements in the soil solution to be assimilated by a plant’s roots [
55]. Otherwise, the ascending ash concentration until 40 days and the subsequent stabilization presented in
Paspalum sp.,
P. plicatulum, and
A. purpussi grasses is similar to the pattern observed in another report, where the ash content exhibited a quadratic behavior, reaching its maximum value at 40 days of age [
55].
The Ca and P levels presented between the studied grasses were similar to those found in tropical forage grown in humid conditions [
57]. Only
P. plicatulum showed superiority in terms of the Ca levels compared with the control grass; however, in terms of the P concentrations, they were similar in all plants. Ca and P are important elements required in high concentrations as they are involved in different processes as enzyme activators (Ca) or constituents of organic compounds (P) [
58]. The Ca:P ratio is a parameter widely used in animal nutrition to assess the availability, absorption, and utilization of these elements [
15]. The expected range for the maintenance of optimal ruminal performance is between 1 and 7, resulting in increased possibility of metabolic disorders with higher values [
59]. Among the studied grasses, Ca:P ratios between 0.61 and 2.54 were found, with the lowest value being associated with the “control” grass at 50 days. The native species had a minimum value of 1.14, suggesting adequate availability of these elements for animal nutrition.
The observed NDF values (61% to 73.9%) were consistent with those reported in
Brachiaria and
Panicum genera [
44,
45,
60], as well as other grass species used for grazing herbivorous animals [
52,
55,
57]. All grasses presented similar NDF concentrations during the experimental period. In
P. plicatulum,
Paspalum sp., and the “control” plant, the NDF levels increased over time because of cell wall component accumulation (cellulose, hemicellulose, and lignin) that usually occurs with advancing plant age and is commonly reported in forage grasses [
17,
55]. The native
A. compresus plant showed relatively stable NDF values. Similar results have been reported for this species during the growth period between 21 and 63 days of age under tropical Mexican dry conditions [
61]. The results suggest that, in
A. compresus, significant growth of the cell wall components could occur after 50 days of age.
The ADF concentrations among the studied plants (30.7% and 42.6%) were in correspondence with the observed values in
B. brizantha,
B. humidicola,
B. hybrid Mulato, and
P. notatum under tropical conditions [
44,
57]. In this study, the control grass showed the lowest ADF values at 30 and 40 days; however, no difference was observed at 50 days with respect to that of the native plants. Under warm conditions typical of tropical areas, plants’ metabolic processes are accelerated, encouraging photosynthetic products to be used mainly in the continuous formation of cell wall components [
53,
62]. In native plants, this adaptive mechanism possibly develops in the early stages to guarantee their subsistence and longevity. This may be the reason for the higher cell wall lignification observed in native plants. On the other hand, in the
A. compresus grass, the ADF increase rate was slower than that in the other grasses. This is similar to the observation of the NDF values and suggests that
A. compresus grass presents a slower maturation rate within the “savannah bank” ecosystem.
Regarding the lignin content, the range found among grasses (7.8% to 10.9%) was also similar to that in the results reported in other species [
44,
57]. The lignin concentrations were comparable among the evaluated plants. However, it should be emphasized that the values in the “control” plant were higher than those reported for the same species in the Colombian Caribbean [
43], different cultivars of
B. brizantha,
B. humidicola, and other grasses [
55,
62,
63]. This can be attributed to the environmental effects, such as the high temperatures in the study area (>30 °C), which accelerate plant maturation, resulting in an increase in the cell wall content, including the lignin level [
42,
62].
In general terms, fiber variables (NDF, ADF, and lignin) presented similar behavior among the evaluated species. Fiber is important because it constitutes the partially digestible fraction in the gastrointestinal tract of herbivorous animals and is composed of complex polysaccharides, such as cellulose, hemicellulose, and pectin, as well as lignin, which is rich in phenolic compounds [
64]. The fiber content increases as plants mature [
51,
52,
65], and this effect is more intense in tropical species, with increases ranging from 11% to 18% when compared with temperate climate species [
53]. This is consistent with the findings obtained in the present study, where increases in the fiber levels, especially in the lignin concentrations, were found. Adverse environmental conditions (high temperatures, solar radiation, and low precipitation) induce phenological and physiological changes in plants as an adaptation mechanism to prevent water loss, including the preference of energy expenditure for the formation of support and defense structures over leaf formation for rapid growth. In warm conditions, plants express traits that ensure their longevity, rather than those that promote rapid growth [
53].
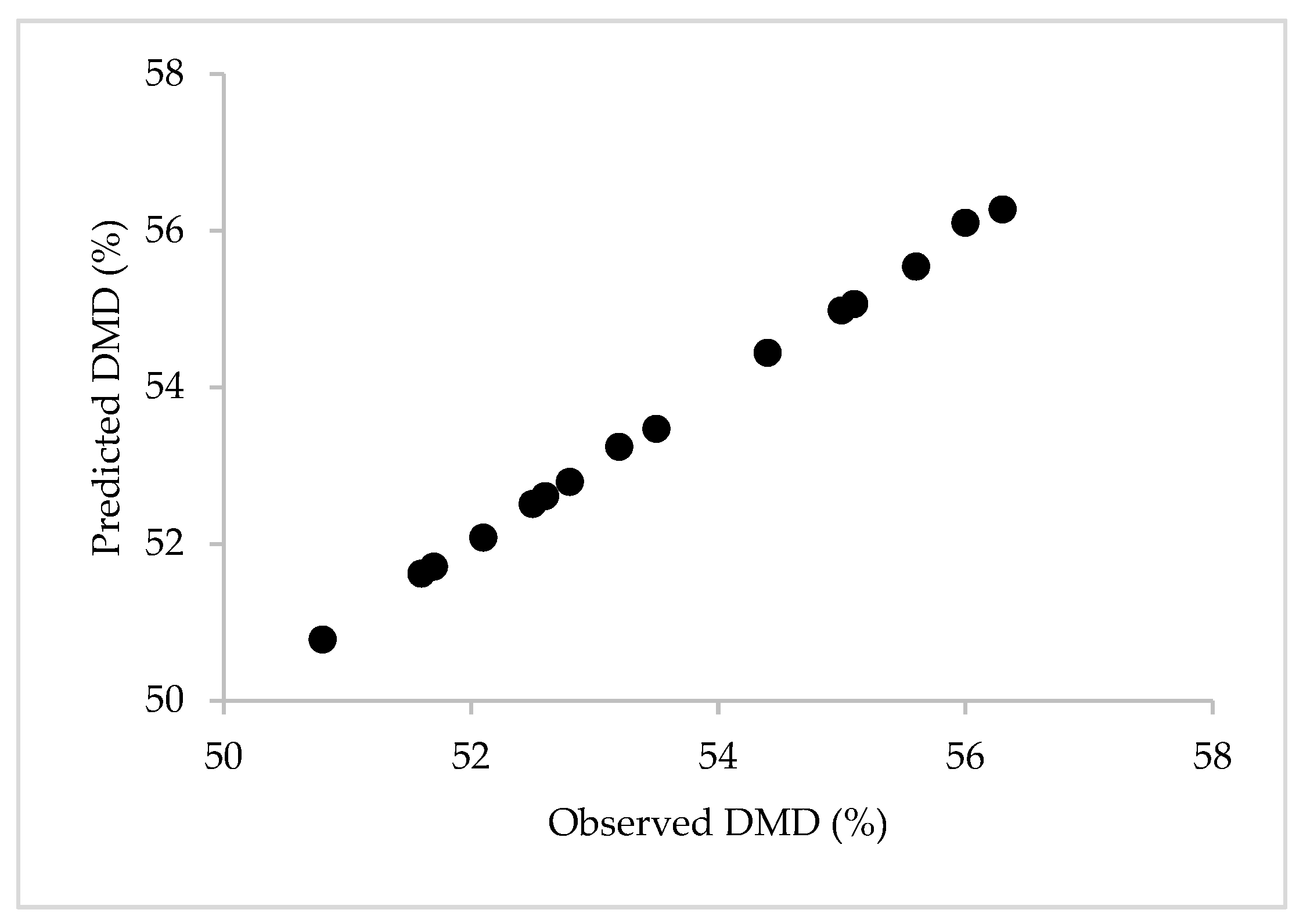
The estimated DMD values (50.8% to 56.3%) were comparable to those in other reports described in tropical pastures [
14,
53]. DMD is one of the main criteria for evaluating the nutritional potential of forage as a base diet in animal nutrition, as it represents the proportion of plant material that can be digested by herbivores [
14,
42]. Native grasses were comparable to the control plant in terms of DMD, although some of them, such as
A. purpussi, P. plicatulum, and
A. compresus, presented higher values during some of the evaluated periods. This occurred because, although these plants had fiber levels, similar to the control, their protein concentrations were higher, and under this condition, digestibility may be improved [
66]. Generally, it was observed that DMD was reduced over time in all of the evaluated plants, which can be attributed to the negative association between DMD and the fiber content. Increases in the cellulose, hemicellulose, and lignin levels reduce forage digestibility and intake potential [
42]. In the case of
A. compresus, DMD remained stable at first before increasing in the final sampling phase. The lower fiber levels and constant DMD obtained in
A. compresus allowed us to ensure that, under the evaluated conditions, this grass presented a lower maturation rate than the other species under study. However, this implied lower DM production per hectare, which was an undesirable characteristic as it would reduce the feed availability for animals.
4.2. Correlations and Regression Analysis
The DM levels were positively associated with ADF, which can be attributed to the fact that DM increases as the grass grows, causing cell wall thickening and thus an increase in fibrous components, including the ADF fraction [
51]. These results are consistent with the positive correlation observed between the NDF and lignin contents, both of which are also part of the fibrous fraction and increase over time [
51,
52,
65]. The ash and Ca levels also showed a direct correlation, which was expected because Ca was one of the elements present in the ash fraction.
In this study, the grasses’ heights showed negative correlations with the DM and ADF contents. As mentioned previously, as grasses reached maturity a parallel increase in the DM and fiber (ADF) content would also be expected [
51,
59]. However, the inverse relationship observed between these variables could be attributed to differences in growth habits among the evaluated plants. The bunch growth of the “control” grass and
P. plicatulum implied a taller height than that of the other native species, which presented stoloniferous or rhizomatous growth. This behavior was reflected in data dispersion with an inverse relationship between the variables, which was declared statistically significant.
DMD presented a positive association with CP and a negative association with different fiber fractions, such as NDF and lignin. Reports in the literature indicate that digestibility is largely determined by the chemical composition of the plant, with higher values in plants with high CP contents and lower in those with high fiber values [
14,
52,
53].
The result of multiple linear regression analysis showed that the CP and ADF levels significantly influenced the DMD. These results agree with those of other studies, which indicate that CP and ADF are two of the most important factors in determining the DMD of pastures [
62]. The parameter’s magnitude suggests that, for a unit increase in the CP percentage and keeping the ADF constant, the DMD increased by 0.80%, while it decreased by 0.31% for a unit increase in the ADF level. Protein favors DMD because it can be easily degraded by ruminal microorganisms [
66]. Grasses with a low protein content due to advanced maturity are less digestible [
52]. Similarly, ADF is a nearly indigestible fraction composed of cellulose, lignin, cutting, and lignified proteins that limit the cell wall carbohydrate degradation at the ruminal level. Therefore, ADF is used to estimate the digestibility, energy content, and consumption potential of forage species [
65,
67].
Plants expressing adaptability traits, high forage biomass production, and nutritional quality are the best choices for grazing animals [
64]. Understanding the productive and quality characteristics of forage species with nutritional potential is crucial for optimizing pasture management and establishing forage mixtures with desirable nutritional characteristics to ensure adequate animal consumption and productive performance [
16]. The results obtained showed that, in the evaluated agroecosystem, native grasses performed similarly to the “control” grass in terms of the forage biomass production (
P. plicatulum) and protein and fiber contents (
P. plicatulum,
A. purpussi, and
Paspalum sp.), and exhibited higher mineral levels, including Ca and P. These observations are promising, especially because the observed performance was obtained without the use of agronomic management techniques, such as fertilization and irrigation. This suggests that an integrated management program can improve the productivity of native pastures [
23].
Under tropical conditions and in different agroecological scenarios, such as floodplain savannahs, livestock activity with herbivorous animals relies on pastures because they are the most abundant source of nutrients in the area [
23,
68]. The native grasses’ species diversity present in the “bank” physiographic position constitutes a valuable resource to find nutritional alternatives that ensure feed for animals facing the current climate change scenarios [
15]. In livestock production systems on floodplain savannahs, it is common to establish improved pastures as the first alternative for animal feeding. However, it is not considered that many of these species present adaptive difficulties that affect their yield and nutritional value [
64,
69], and overcoming these difficulties requires intensive management that is not common in most of the production systems in such regions. The results indicate that, under the typical “savannah banks” unit dynamics, some introduced species, such as
B. hybrid Mulato (“control”), present similar or even lower biomass production and chemical compositions than some of the native grasses included in the present study.
These results encourage research efforts to characterize the productive potential of native plants in different ecosystems, with the aim of incorporating them into livestock farms, offering to producers a greater diversity of feed alternatives with more flexible management. It is expected that, through their adaptive characteristics, these forage species can contribute to strengthening soil–plant–animal interactions and promote the generation of environmental services, such as the protection and improvement of soils’ physical–chemical properties, nutrient recycling, and greenhouse gas fixation [
15,
64].
To evaluate the forage potential of a plant species, in addition to its nutritional composition and forage production, it is also necessary to know the animal response in terms of consumption and productive performance [
64]. Although most of the native grasses evaluated in this study are recognized as the main food source in the area, especially during the rainy season [
6], experimental trials are still required to precisely evaluate animal consumption and the transformation efficiency of the supplied nutrients in animal products [
65].


 ,
, 




{kind=link}
{kind=link}
{kind=link}