
The Influence of Groves on Aboveground Arthropod Diversity and Evolution in a Vineyard in Southern Romania
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
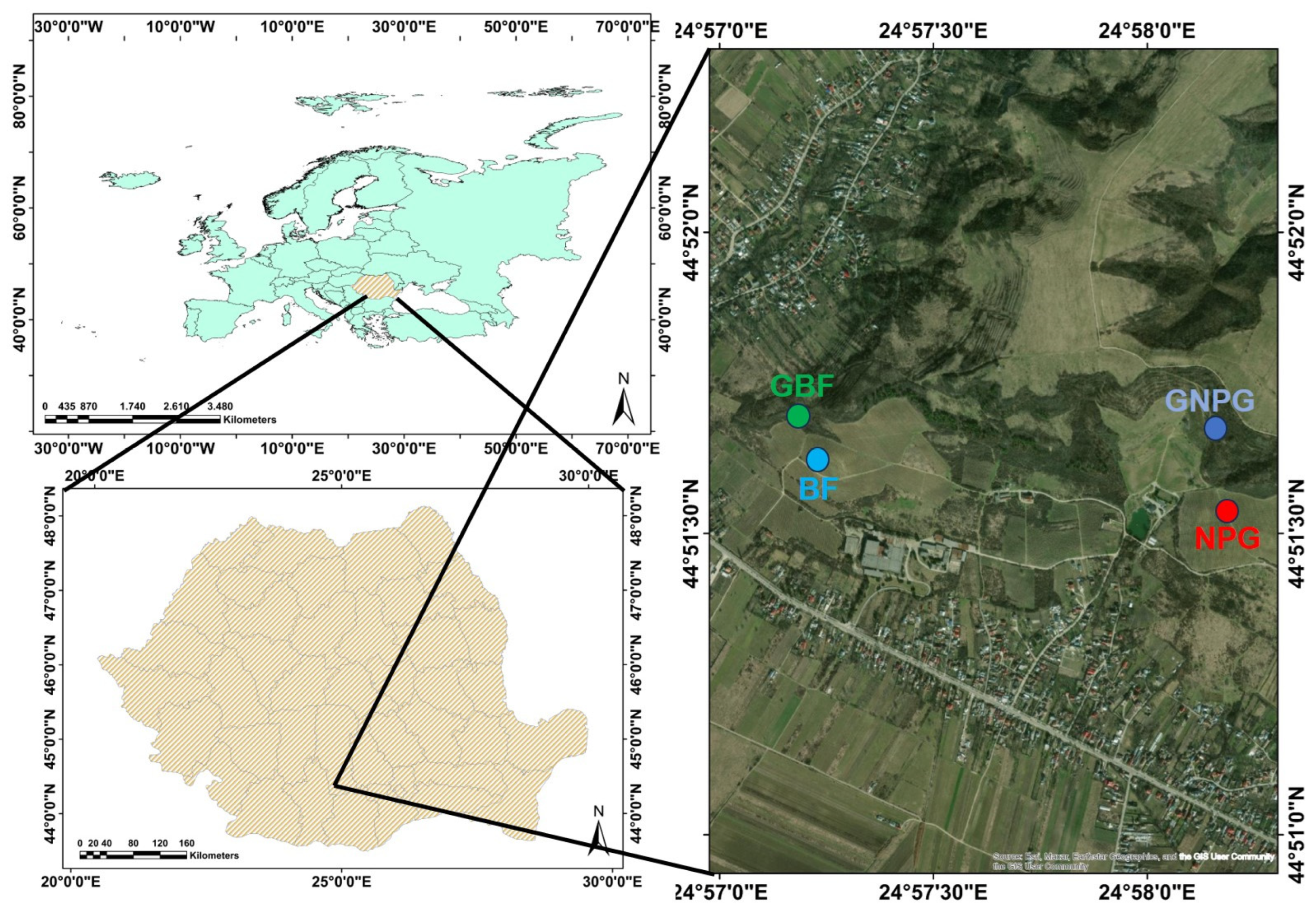
2.1. Study Site and Data Collection
2.2. Data Analysis
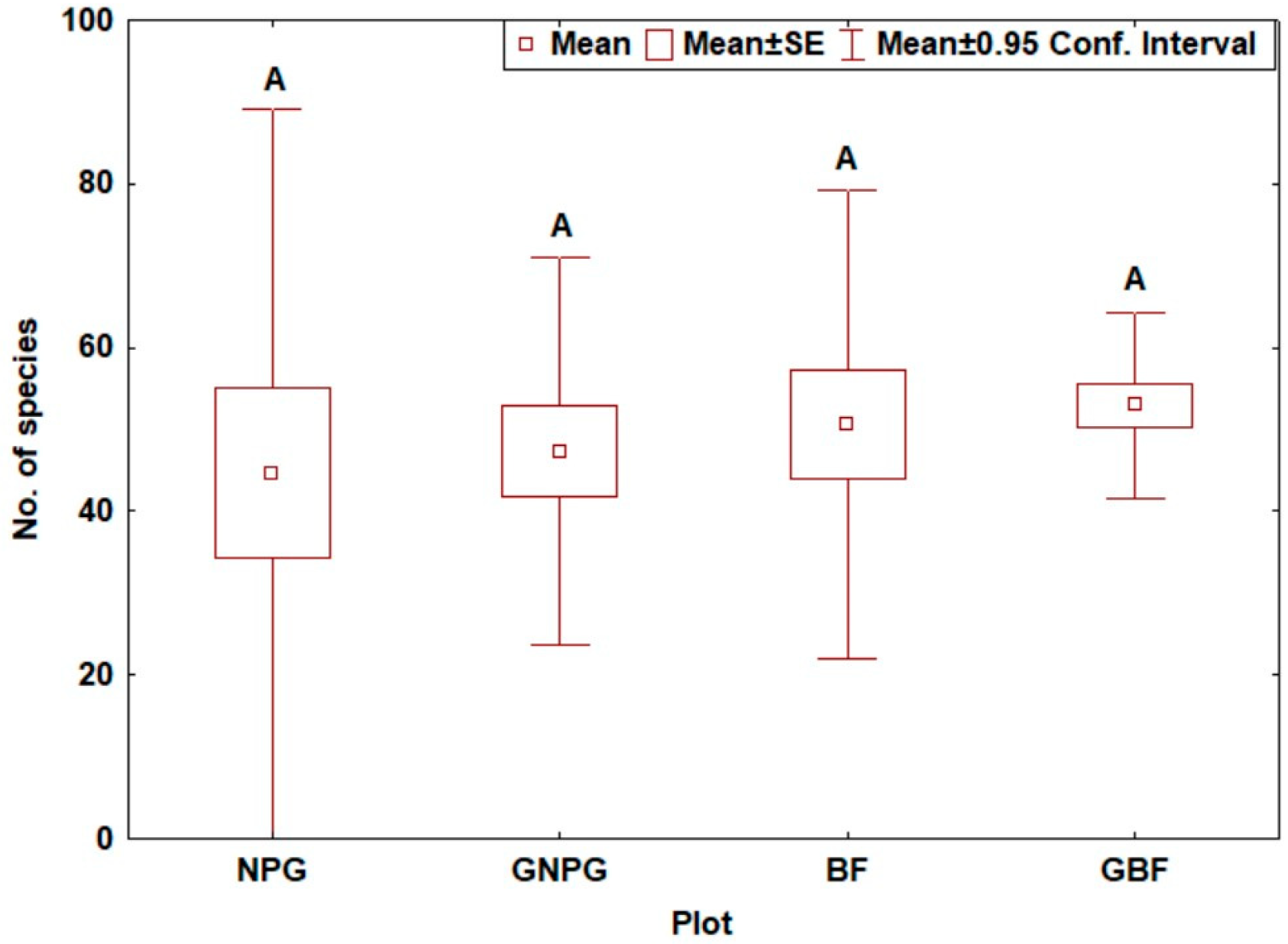
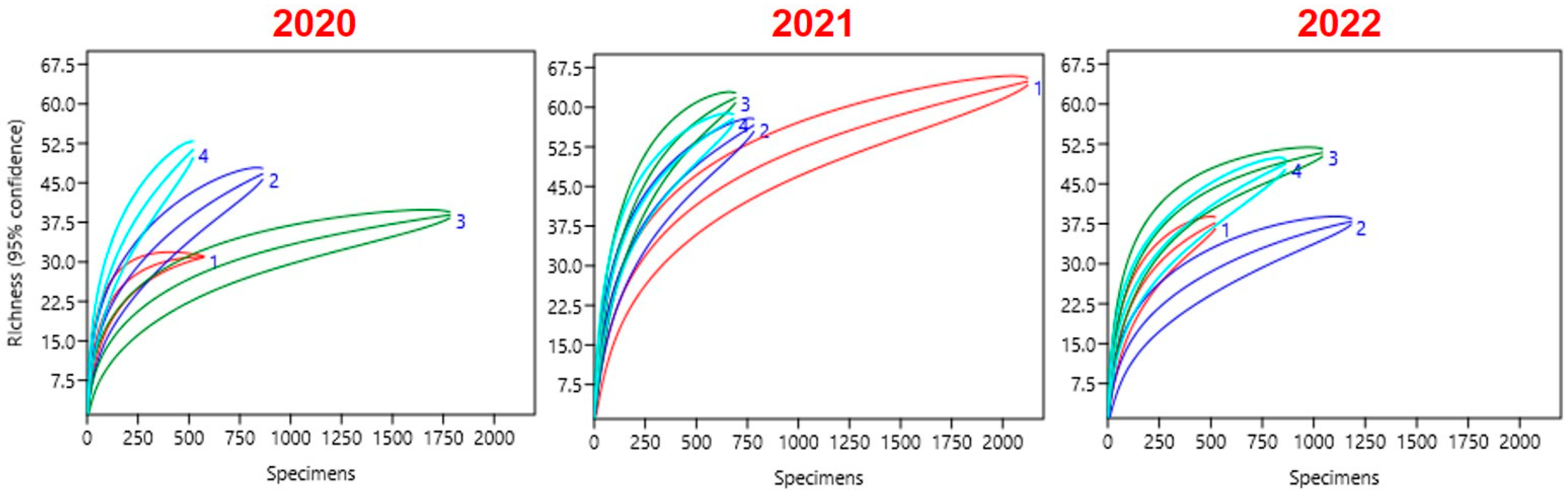
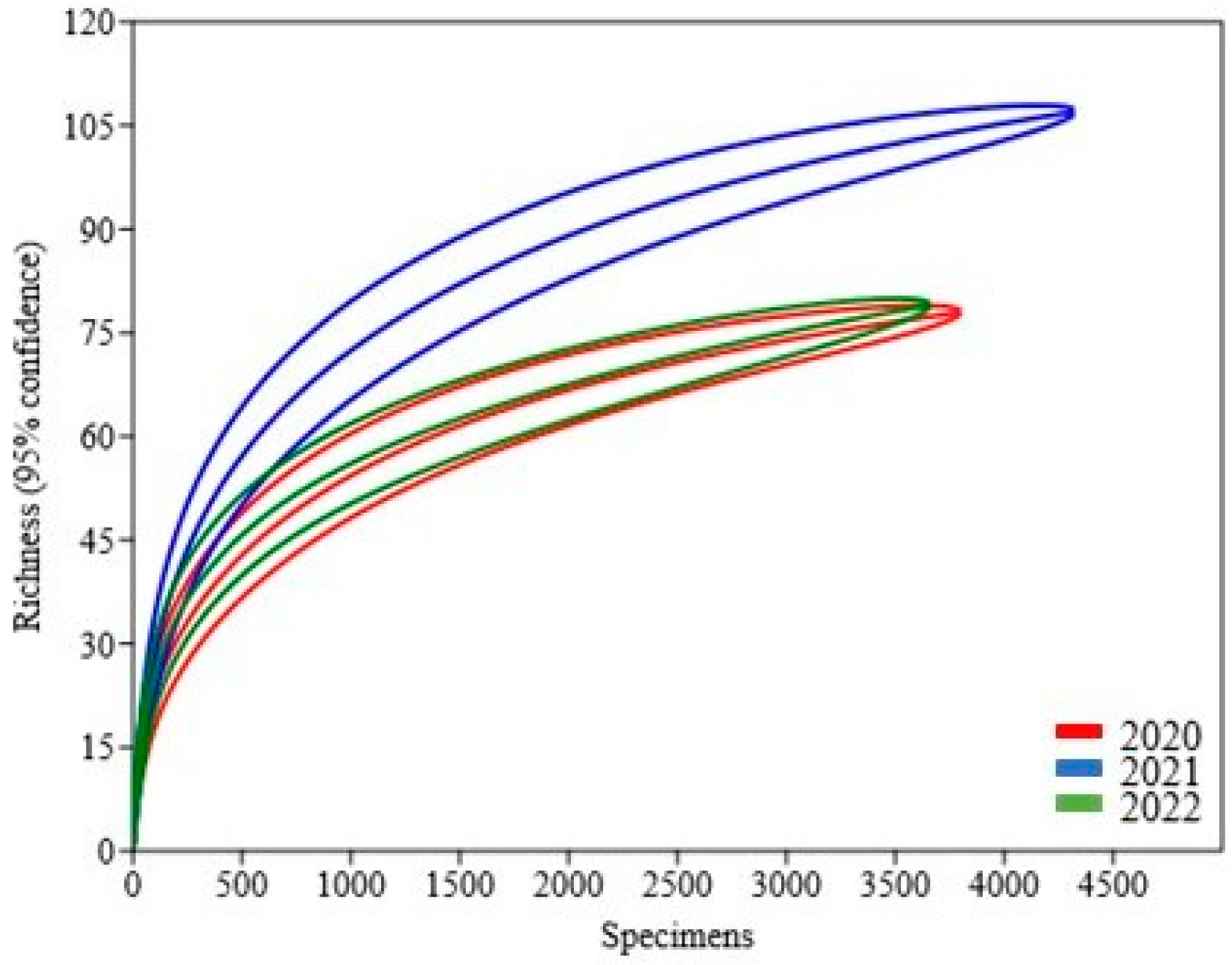
3. Results
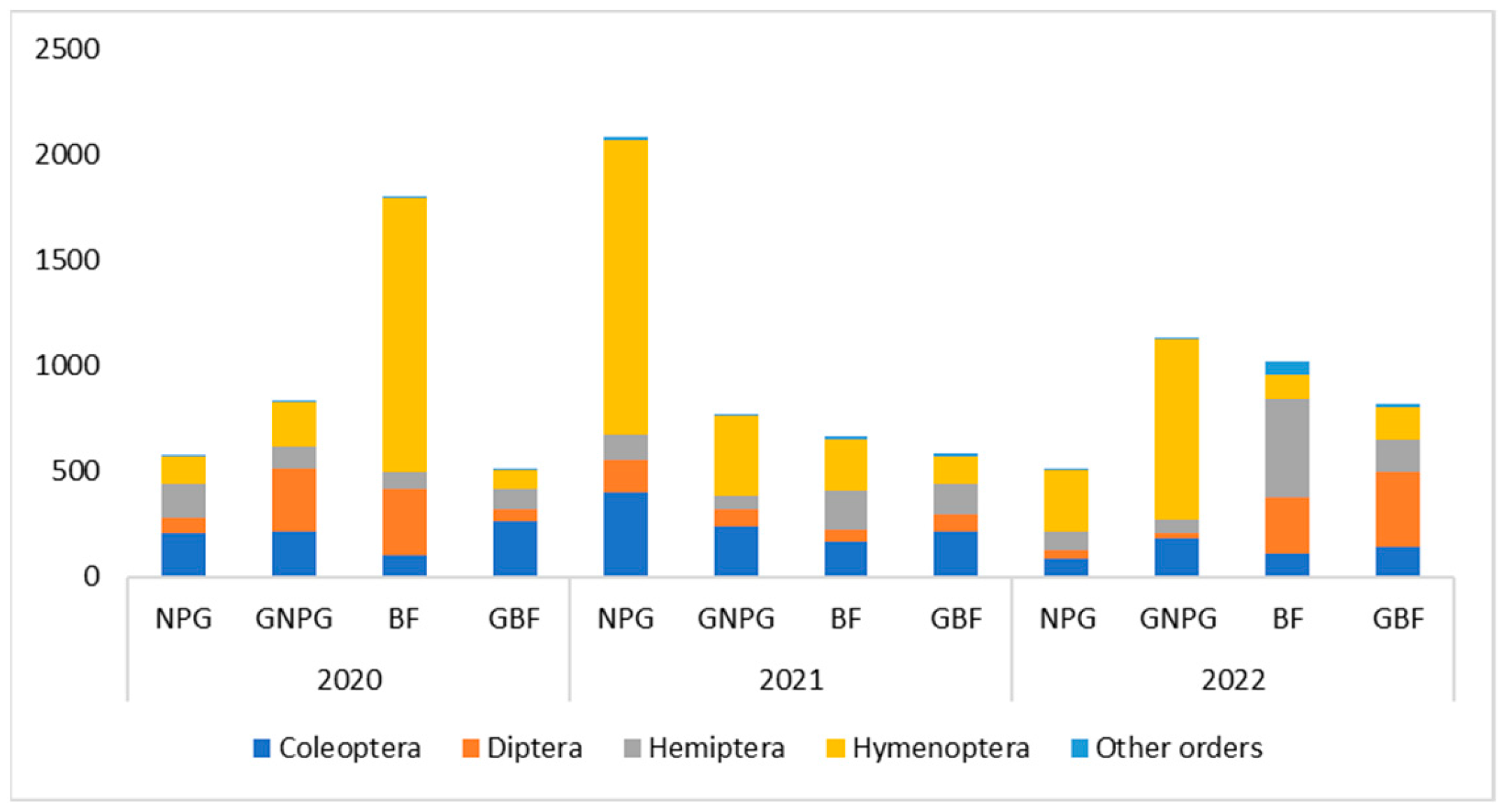
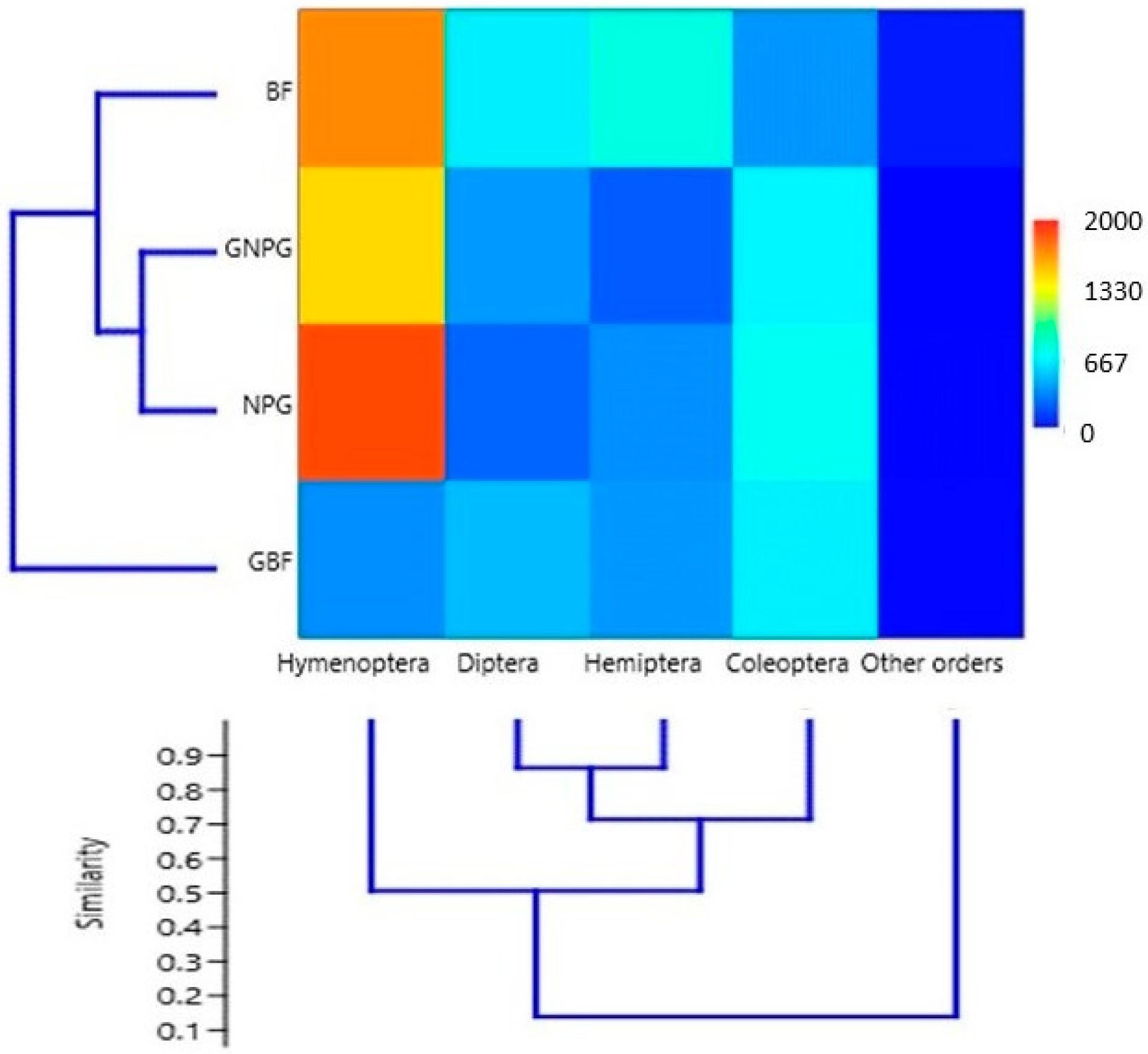
- In the case of NPG, the insects captured from the order Hymenoptera were significantly more numerous than those in the “other orders” group (p < 0.05) but not significantly different from Coleoptera, Diptera, and Hemiptera (p > 0.05).
- For GNPG, the insects captured from the Hymenoptera group were significantly more numerous than those in Diptera, Hemiptera, and other orders (p < 0.05) but not significantly different from those in Coleoptera (p > 0.05).
- Regarding BF, there were no significant differences (p > 0.05) in the number of individuals captured among the five orders.
- In GBF, the insects captured from the Diptera and Coleoptera groups were significantly more numerous (p < 0.05) than those in the “other orders” group but not significantly more numerous (p > 0.05) than those in Hemiptera and Hymenoptera.

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | NPG | GNPG | BF | GBF |
|---|---|---|---|---|---|
| 1 | Acanthoscelides sp. | 0 | 0 | 0 | 1 |
| 2 | Adonia variegata | 0 | 1 | 3 | 1 |
| 3 | Aelia acuminata | 0 | 1 | 0 | 1 |
| 4 | Agrilus sp. | 2 | 2 | 1 | 3 |
| 5 | Agriotes lineatus | 0 | 0 | 1 | 1 |
| 6 | Agriotes ustulatus | 1 | 2 | 2 | 1 |
| 7 | Ampedus nigerrimus | 1 | 0 | 0 | 0 |
| 8 | Andrena cineraria | 0 | 0 | 1 | 0 |
| 9 | Andrena subopaca | 3 | 3 | 1 | 2 |
| 10 | Andrena wilkella | 0 | 1 | 1 | 0 |
| 11 | Anisosticta novemdecimpunctata | 0 | 0 | 0 | 2 |
| 12 | Anthaxia cichorii | 1 | 1 | 1 | 1 |
| 13 | Anthaxia nitidula signaticollis | 1 | 1 | 1 | 0 |
| 14 | Anthocomus rufus | 1 | 0 | 0 | 0 |
| 15 | Anthomyia procellaris | 3 | 3 | 3 | 3 |
| 16 | Apis mellifera | 3 | 3 | 3 | 1 |
| 17 | Araneus diadematus | 1 | 0 | 0 | 0 |
| 18 | Arge ochropus | 2 | 1 | 0 | 0 |
| 19 | Athalia rosae | 1 | 2 | 1 | 1 |
| 20 | Bombus terrestris | 2 | 1 | 0 | 0 |
| 21 | Bruchidius villosus | 0 | 0 | 1 | 0 |
| 22 | Calliptamus barbarus | 0 | 0 | 0 | 1 |
| 23 | Cantharis rubra | 0 | 0 | 0 | 1 |
| 24 | Cantharis rustica | 0 | 1 | 0 | 0 |
| 25 | Carpomya incompleta | 0 | 0 | 2 | 1 |
| 26 | Carpomya schineri | 1 | 1 | 1 | 2 |
| 27 | Centrotus cornutus | 1 | 1 | 1 | 0 |
| 28 | Ceresa bubalus | 1 | 2 | 2 | 3 |
| 29 | Cheiracanthium mildei | 2 | 1 | 1 | 2 |
| 30 | Chlorophorus sartor | 1 | 1 | 0 | 0 |
| 31 | Chlorophorus varius | 3 | 3 | 3 | 2 |
| 32 | Chorthippus brunneus | 0 | 1 | 1 | 0 |
| 33 | Chorthippus vagans | 1 | 0 | 0 | 0 |
| 34 | Chrysis ignita | 1 | 0 | 0 | 1 |
| 35 | Chrysoperla carnea | 2 | 1 | 2 | 1 |
| 36 | Chrysura refulgens | 1 | 0 | 0 | 0 |
| 37 | Cicadetta hannekeae sp. | 1 | 2 | 2 | 2 |
| 38 | Clytra laeviuscula | 2 | 2 | 1 | 0 |
| 39 | Clytus lama | 2 | 2 | 2 | 2 |
| 40 | Coccinella septempunctata | 1 | 2 | 2 | 1 |
| 41 | Colias croceus | 1 | 0 | 0 | 0 |
| 42 | Conocephalus strictus | 0 | 1 | 0 | 1 |
| 43 | Coreus marginatus | 1 | 1 | 0 | 1 |
| 44 | Corythucha ciliata | 3 | 3 | 3 | 3 |
| 45 | Cryptocephalus sp. | 2 | 1 | 2 | 1 |
| 46 | Danacea pallipes | 3 | 3 | 3 | 3 |
| 47 | Deraeocoris ruber | 0 | 1 | 2 | 1 |
| 48 | Dolycoris baccarum | 0 | 0 | 0 | 1 |
| 49 | Drosophila melanogaster | 1 | 1 | 1 | 1 |
| 50 | Ellychnia corrusca | 1 | 0 | 0 | 0 |
| 51 | Episyrphus balteatus | 0 | 0 | 0 | 1 |
| 52 | Eristalis tenax | 2 | 3 | 3 | 3 |
| 53 | Eupteryx atropunctata | 0 | 0 | 0 | 1 |
| 54 | Eurydema (Eurydema) oleracea | 0 | 0 | 1 | 0 |
| 55 | Eysarcoris inconspicuous | 0 | 1 | 0 | 1 |
| 56 | Eysarcoris ventralis | 0 | 0 | 0 | 1 |
| 57 | Figitidae sp. | 2 | 2 | 2 | 2 |
| 58 | Forficula auricularia | 2 | 1 | 1 | 2 |
| 59 | Geocoris erythrocephalus | 1 | 1 | 3 | 2 |
| 60 | Gonioctena fornicate | 0 | 1 | 0 | 0 |
| 61 | Graphosoma italicum | 0 | 1 | 0 | 0 |
| 62 | Graphosoma lineatum | 2 | 1 | 0 | 1 |
| 63 | Gymnocheta viridis | 0 | 0 | 2 | 0 |
| 64 | Halictus scabiosae | 0 | 0 | 1 | 1 |
| 65 | Halictus sexcinctus | 0 | 0 | 1 | 0 |
| 66 | Halyzia sedecimguttata | 0 | 0 | 0 | 1 |
| 67 | Harmonia axyridis | 3 | 3 | 3 | 3 |
| 68 | Harrisina metallica | 0 | 0 | 1 | 0 |
| 69 | Hedychrum nobile | 0 | 0 | 1 | 0 |
| 70 | Heliophanus kochii | 0 | 1 | 0 | 1 |
| 71 | Hippodamia variegata | 1 | 1 | 0 | 0 |
| 72 | Holcostethus albipes | 0 | 0 | 0 | 1 |
| 73 | Holcostethus limbolarius | 1 | 0 | 0 | 2 |
| 74 | Ichneumon gracilentus | 0 | 0 | 0 | 2 |
| 75 | Iphiclides podalirius | 0 | 2 | 0 | 1 |
| 76 | Lagria hirta | 0 | 0 | 0 | 1 |
| 77 | Lampyris noctiluca | 1 | 2 | 0 | 1 |
| 78 | Larinus centaurii | 0 | 0 | 0 | 1 |
| 79 | Lasiommata maera | 0 | 0 | 1 | 0 |
| 80 | Lasius niger | 2 | 2 | 0 | 1 |
| 81 | Libellula depressa | 1 | 0 | 0 | 0 |
| 82 | Lucilia sericata | 0 | 0 | 1 | 1 |
| 83 | Lygaeus kalmii | 0 | 1 | 0 | 0 |
| 84 | Lygus pabulinus | 0 | 1 | 0 | 0 |
| 85 | Machimus atricapillus | 1 | 0 | 0 | 0 |
| 86 | Malachius bipustulatus | 2 | 2 | 1 | 2 |
| 87 | Maniola jurtina | 0 | 0 | 0 | 1 |
| 88 | Mantis religiosa | 1 | 0 | 0 | 0 |
| 89 | Meiosimyza sp. | 0 | 1 | 1 | 0 |
| 90 | Melanargia galathea | 0 | 0 | 1 | 2 |
| 91 | Melitaea deione | 0 | 0 | 1 | 1 |
| 92 | Micrommata virescens | 0 | 0 | 1 | 0 |
| 93 | Mordella sp. | 1 | 1 | 1 | 1 |
| 94 | Musca autumnalis | 1 | 2 | 2 | 2 |
| 95 | Musca domestica | 3 | 3 | 3 | 3 |
| 96 | Myathropa florea | 0 | 0 | 0 | 1 |
| 97 | Neomyia cornicina | 0 | 0 | 1 | 0 |
| 98 | Neoscona crucifera | 0 | 0 | 0 | 1 |
| 99 | Nysius graminicola | 0 | 0 | 1 | 0 |
| 100 | Ochlodes sylvanoides | 0 | 1 | 0 | 0 |
| 101 | Ochlodes sylvanus | 1 | 0 | 0 | 0 |
| 102 | Ochlodes venatus | 1 | 0 | 1 | 1 |
| 103 | Oecanthus californicus | 0 | 0 | 0 | 1 |
| 104 | Oecanthus pellucens | 0 | 0 | 0 | 1 |
| 105 | Oedemera flavipes | 3 | 3 | 3 | 1 |
| 106 | Oedemera podagrariae | 0 | 0 | 1 | 0 |
| 107 | Onthophagus sp. | 1 | 1 | 0 | 0 |
| 108 | Oodes helopioides | 3 | 2 | 2 | 2 |
| 109 | Orius insidiosus | 1 | 1 | 1 | 1 |
| 110 | Oulema obscura | 0 | 0 | 1 | 1 |
| 111 | Oxycarenus lavaterae | 1 | 1 | 0 | 0 |
| 112 | Pachybrachis tessellatus | 0 | 0 | 0 | 2 |
| 113 | Panorpa communis | 2 | 1 | 2 | 2 |
| 114 | Penthimia nigra | 1 | 0 | 0 | 0 |
| 115 | Penthimia sp. | 0 | 1 | 1 | 0 |
| 116 | Peponapis pruinosa | 0 | 0 | 1 | 0 |
| 117 | Phaenicia sericata | 0 | 0 | 0 | 1 |
| 118 | Phaneroptera nana | 0 | 0 | 1 | 0 |
| 119 | Philaenus spumarius | 3 | 3 | 3 | 3 |
| 120 | Phlogotettix cyclops | 0 | 1 | 1 | 1 |
| 121 | Phyllotreta undulata | 3 | 3 | 3 | 2 |
| 122 | Pieris rapae | 0 | 0 | 1 | 0 |
| 123 | Polistes dominulus | 3 | 2 | 3 | 2 |
| 124 | Pollenia rudis | 0 | 0 | 0 | 1 |
| 125 | Priocnemis perturbator | 0 | 1 | 0 | 0 |
| 126 | Propylea quatuordecimpunctata | 2 | 1 | 2 | 2 |
| 127 | Protapion fulvipes | 0 | 0 | 0 | 1 |
| 128 | Psyllobora vigintiduopunctata | 0 | 2 | 3 | 2 |
| 129 | Raglius alboacuminatus | 0 | 1 | 1 | 0 |
| 130 | Rhagoletis completa | 0 | 0 | 0 | 2 |
| 131 | Rhynchomitra microrhina | 1 | 0 | 1 | 1 |
| 132 | Sarcophaga bercaea | 1 | 1 | 0 | 1 |
| 133 | Sarcophaga carnaria | 1 | 0 | 0 | 0 |
| 134 | Sarcophaga sp. | 1 | 3 | 3 | 3 |
| 135 | Scaphidium quadrimaculatum | 0 | 0 | 1 | 0 |
| 136 | Scaphoideus titanus | 2 | 2 | 3 | 3 |
| 137 | Sceliphron caementarium | 1 | 1 | 0 | 0 |
| 138 | Scutigera coleoptrata | 0 | 0 | 0 | 1 |
| 139 | Scymnus frontalis | 3 | 3 | 3 | 2 |
| 140 | Scymnus rubromaculatus | 0 | 0 | 3 | 3 |
| 141 | Scythris sinensis | 0 | 0 | 1 | 0 |
| 142 | Sitochroa verticalis | 0 | 1 | 0 | 0 |
| 143 | Sphecodes hyalinatus | 0 | 0 | 0 | 1 |
| 144 | Stenocorus sp. | 0 | 0 | 0 | 1 |
| 145 | Stenocorus vestitus | 0 | 1 | 0 | 0 |
| 146 | Stevenia deceptoria | 0 | 0 | 0 | 1 |
| 147 | Stictocephala bisonia | 1 | 1 | 0 | 1 |
| 148 | Stomoxys calcitrans | 1 | 0 | 0 | 0 |
| 149 | Strongylocoris leucocephalus | 0 | 1 | 1 | 0 |
| 150 | Synema globosum | 0 | 0 | 0 | 1 |
| 151 | Tachycixius pilosus | 0 | 0 | 0 | 1 |
| 152 | Tachysphex pompiliformis | 0 | 1 | 0 | 0 |
| 153 | Tettigonia viridissima | 1 | 0 | 2 | 0 |
| 154 | Tiphia femorata | 1 | 1 | 2 | 3 |
| 155 | Tomoplagia obliqua | 1 | 1 | 0 | 0 |
| 156 | Tomoxia bucephala | 2 | 2 | 2 | 2 |
| 157 | Trichaetipyga juniperina | 0 | 1 | 1 | 1 |
| 158 | Trichodes apiarius | 1 | 1 | 1 | 0 |
| 159 | Tritomegas sexmaculatus | 0 | 0 | 0 | 1 |
| 160 | Vespa crabro | 3 | 3 | 2 | 1 |
| 161 | Vespula germanica | 3 | 3 | 3 | 2 |
| 162 | Vespula vulgaris | 2 | 3 | 3 | 3 |
| 163 | Voria ruralis | 0 | 0 | 3 | 3 |
| 164 | Zygaena ephialtes | 2 | 1 | 0 | 0 |
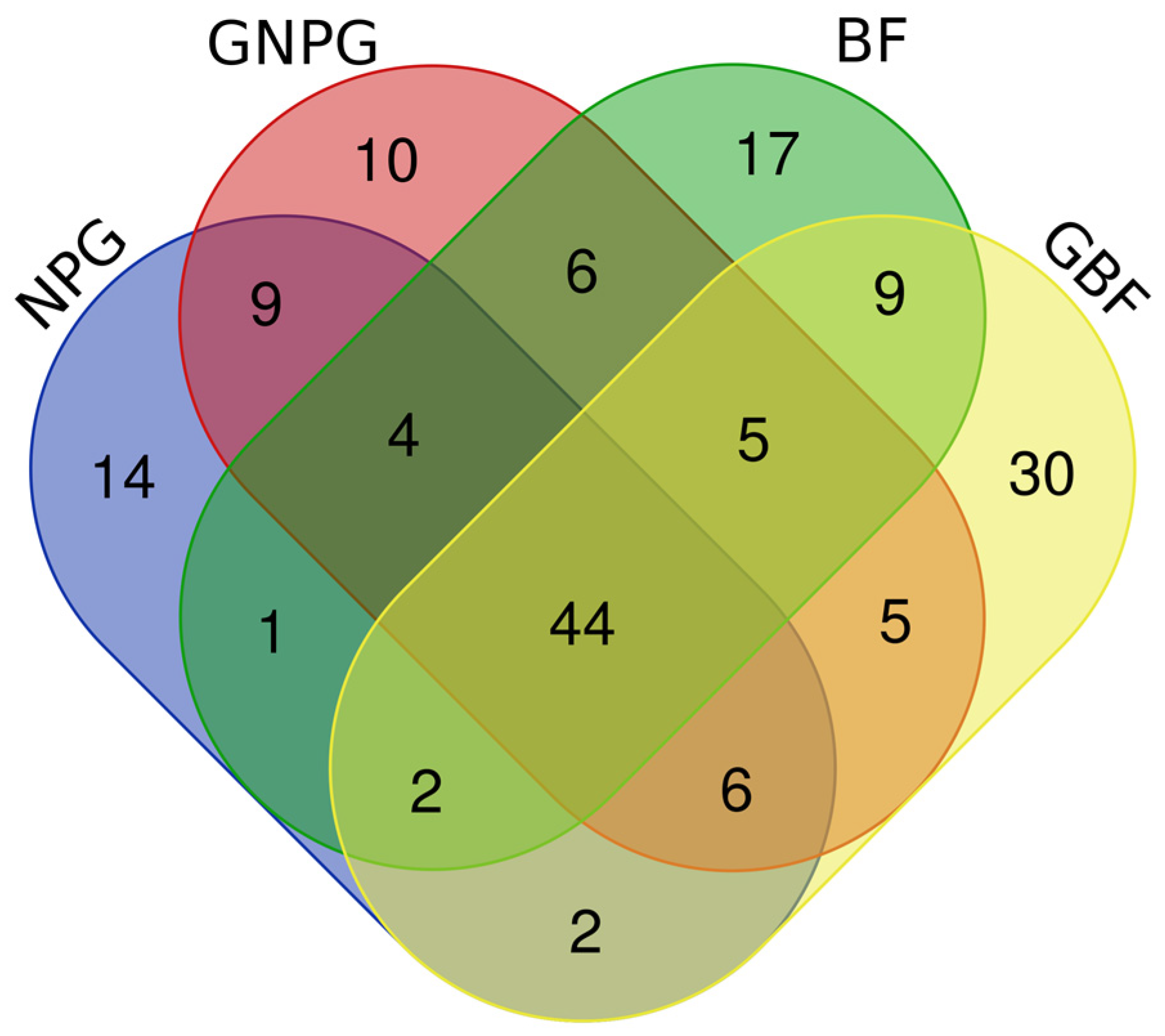
 unique species in NPG;
unique species in NPG;  unique species in GNPG;
unique species in GNPG;  unique species in BF;
unique species in BF;  unique species in GBF.
unique species in GBF.| Order | Family | Species | Feeding, Benefits | References | Lots |
| Araneae | Araneidae | Neoscona crucifera | Insects, Lepidoptera, mostly prefer soft-bodied, immature stages with more internal body fluid, especially the homopterans. | [48,49,50] | GBF. |
| Eutichuridae | Cheiracanthium mildei | Various insect pests, especially Spodoptera littoralis and Phyllonorycter blancardella. | [51] | BF, GBF, NPG, GNPG. | |
| Coleoptera yanbnb | Coccinelidae | Adonia variegata | Aphis gossypii | [52] | BF, GBF, NPG, GNPG. |
| Psyllobora vigintiduopunctata | Powdery mildew spores, such as Oidium-infected Euonymus japonica, Erysiphe holosericea, Podosphaera xanthii, pomegranate tree aphids; powdery mildew of trees, shrubs, and herbaceous plants (prefer powdery mildew on Trifolium, Melilotus, and Medicago (Fabaceae) | [53,54,55,56,57,58,59,60] | BF, GBF, GNPG. | ||
| Scymnus rubromaculatus | Mites, aphids, and crustaceans. | [61,62,63] | BF, GBF. | ||
| Anisosticta novemdecimpunctata | Aphids and other soft insects. | [64,65] | GBF. | ||
| Halyzia sedecimguttata | Powdery mildew (Ascomycotina: Erysiphales) of trees, shrubs, and herbaceous plants; prefers powdery mildew on Trifolium, Melilotus, and Medicago (Fabaceae). | [58] | GBF. | ||
| Coccinella septempunctata | Fungal spores, aphids (e.g., Rhopalosiphum padi). | [66] | BF, GBF, NPG, GNPG. | ||
| Propylea quatuordecimpunctata | Aphids (e.g., the Aphidoidea superfamily). | [67,68] | BF, GBF, NPG, GNPG. | ||
| Melyridae | Danacea pallipes | Larvae feed on fungal mycelia. | [69] | BF, GBF, NPG, GNPG. | |
| Malachius bipustulatus | Lymantria dispar; small insects found on flowers; nymphae of some xilophagous insects; pollen; Oulema melanopus L. | [70,71,72,73,74] | BF, GBF, NPG, GNPG. | ||
| Mordellidae | Mordella sp. | Native pollinators. | [75] | BF, GBF, NPG, GNPG. | |
| Diptera | Tachinidae | Gymnocheta viridis | Noctuidae species. | [76] | BF. |
| Voria ruralis | Various caterpillars, but used mainly to control the cabbage looper (Trichoplusia ni) caterpillar; feeds on aphid honeydew;lepidopteran species, particularly Trichoplusia ni. | [77,78,79] | BF, GBF. | ||
| Hemiptera | Geocoridae | Geocoris erythrocephalus | Aphids, whiteflies, thrips, mites, caterpillars, eggs, and larvae (tobacco budworm, soybean loopers). | [80,81] | BF, GBF, NPG, GNPG. |
| Miridae | Deraeocoris ruber | Cacopsylla pyrisuga, younger caterpillar instars of some butterflies, mites, and various other small insects in apple orchards, aphids, Acizzia jamatonica, and Cacopsylla pyrisuga. | [82,83,84] | BF, GBF, GNPG | |
| Heteroptera | Anthocoridae | Orius insidiosus | Thrips larvae and adults (e.g., Flankliniella occidentalis, Bemisia tabaci and Frankliniella occidentalis, Thrips palmi) and other soft-bodied insects. | [85,86,87,88,89] | BF, GBF, NPG, GNPG. |
| Hymenoptera | Andrenidae | Andrena subopaca | Insect pollinators. | [90] | BF, GBF, NPG, GNPG. |
| Andrena wilkella | Pollen, especially from Fabaceae. | [91] | BF, GNPG. | ||
| Andrena cineraria | Pollinator. | [92,93] | BF. | ||
| Apidae | Apis mellifera | Pollen. | [94,95,96] | BF, GBF, NPG, GNPG | |
| Bombus terrestris | Pollinators. | [97] | NPG, GNPG. | ||
| Peponapis pruinosa | Pollen grains from Cucurbita flowers. | [94,98] | BF. | ||
| Halictidae | Halictus scabiosae | Nectar and pollen. | [99] | GBF. | |
| Tiphiidae | Tiphia femorata | Flowers, nectar, and pollen (especially from Apiaceae species); Rhizotrogus solstitialis, Anisoplia austriaca, and several Aphodius species (Aphodiidae). | [30,100] | BF, GBF, NPG, GNPG. | |
| Mecoptera | Panorpidae | Panorpa communis | Dead arthropods, pollen, and fruit, also scavengers of dead insects and rotting fruit | [101,102] | BF, GBF, NPG, GNPG. |
| Neuroptera | Chrysopidae | Chrysoperla carnea | Grapevine moths, Helicoverpa armigera (Hubner), Bemisia tabaci, aphids (e.g., Aphis pomi), mites and mealy bugs, coccids, small homopterous pests; Jacobiasca lybica, Compsus viridivittatus eggs. | [103,104,105,106,107] | BF, GBF, NPG, GNPG. |
References
- Butiuc-Keul, A.; Coste, A. Biotechnologies and Strategies for Grapevine Improvement. Horticulturae 2023, 9, 62. [Google Scholar] [CrossRef]
- Sosa-Zuniga, V.; Vidal Valenzuela, Á.; Barba, P.; Espinoza Cancino, C.; Romero-Romero, J.L.; Arce-Johnson, P. Powdery Mildew Resistance Genes in Vines: An Opportunity to Achieve a More Sustainable Viticulture. Pathogens 2022, 11, 703. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, R.; Dobrei, A.; Eleonora, N.; Alina, D.; Mihaela, M. Study Regarding the Viticulture Activity in Satu Mare County. J. Hortic. For. Biotechnol. 2021, 25, 15–19. [Google Scholar]
- Mihaela, M.; Georgeta, D.A.; Narcisa, S.; Talita, H.; Dobrei, A. Cultivation Technological Features of Grapevine in a Vineyard Holding in Arad County, Romania. J. Hortic. For. Biotechnol. 2022, 26, 31–37. [Google Scholar]
- Oprea, S. Viticultura; AcademicPres: Cluj-Napoca, Romania, 2001; ISBN 973-8266-09-2. [Google Scholar]
- Bernard, M.; Wainer, J.; Carter, V.; Semeraro, L.; Yen, A.; Wratten, S. Beneficial Insects and Spiders in Vineyards: Predators in South-East Australia. Aust. N. Z. Grapegrow Winemak. 2006, 512, 37–48. [Google Scholar]
- Jankielsohn, A. The Importance of Insects in Agricultural Ecosystems. Adv. Entomol. 2018, 6, 62–73. [Google Scholar] [CrossRef]
- Pillai, A.K.; Kumar, M.G.; Kuttalam, S. Computation of Arthropod Biodiversity in Grapes Ecosystem. 2017. Available online: https://www.entomoljournal.com/archives/2017/vol5issue5/PartF/5-4-362-489.pdf (accessed on 25 September 2023).
- Goussard, P. A Guide to Grapevine Abnormalities in South Africa: Grapevine Pests–Leafhoppers, Bollworm, Leaf Roller Caterpillars, Spring Beetles, Wilting Beetles and Thrips (Part 5.6). 2013. Available online: https://www.wineland.co.za/a-guide-to-grapevineabnormalities-in-south-africa-grapevine-pestsleafhoppers-boll-worm-leaf-roller-caterpillars-springbeetles-wilting-beetles-and-thrips-part-5-6/ (accessed on 15 March 2023).
- Reineke, A.; Thiéry, D. Grapevine Insect Pests and Their Natural Enemies in the Age of Global Warming. J. Pest Sci. 2016, 89, 313–328. [Google Scholar] [CrossRef]
- Fîciu, L.; Bădulescu, A.; Brînduse, E. Characterization of Useful and Harmful Insects Biodiversity in Valea Calugareasca Viticultural Ecosystem. Curr. Trends Nat. Sci. Vol 2017, 6, 71–75. [Google Scholar]
- Retallack, M.; Vineyard Biodiversity and Insect Interactions-Establishing and Monitoring Insectariums. GWRDC Regional—SA Central, Adelaide Hills, Currency Creek, Kangaroo Island, Langhorne Creek, McLaren Vale, and Southern Fleurieu Wine Regions. 2011. Available online: https://www.wineaustralia.com/getmedia/fba82be2-ae5d-4b18-a59e 1129854199ac/Rmjr0811VineyardBiodiversityandIns (accessed on 15 March 2023).
- Boudon-Padieu, E.; Maixner, M. Potential Effects of Climate Change on Distribution and Activity of Insect Vectors of Grapevine Pathogens. In Proceedings of the Colloque International et Pluridisciplinaire Sous L’égide de la Chaire UNESCO Vin et Culture, Dijon, France, 28–30 May 2007; Volume 23. Available online: https://pdfs.semanticscholar.org/22a1/de6d85279d1fa 3d61c86feaae2a61c7af138.pdf (accessed on 25 September 2023).
- Bois, B.; Zito, S.; Calonnec, A. Climate vs Grapevine Pests and Diseases Worldwide: The First Results of a Global Survey. OENO One 2017, 51, 133–139. [Google Scholar] [CrossRef]
- Culliney, T.W. Crop Losses to Arthropods. In Integrated Pest Management: Pesticide Problems; Springer: Dordrecht, The Netherlands, 2014; Volume 3, pp. 201–225. [Google Scholar] [CrossRef]
- Lucchi, A.; Benelli, G. Towards Pesticide-Free Farming? Sharing Needs and Knowledge Promotes Integrated Pest Management. Environ. Sci. Pollut. Res. 2018, 25, 13439–13445. [Google Scholar] [CrossRef]
- Scaramozzino, P.L.; Di Giovanni, F.; Loni, A.; Gisondi, S.; Lucchi, A.; Cerretti, P. Tachinid (Diptera, Tachinidae) Parasitoids of Lobesia botrana (Denis & Schiffermüller, 1775) (Lepidoptera, Tortricidae) and Other Moths. Zookeys 2020, 934, 111. [Google Scholar] [CrossRef] [PubMed]
- Iltis, C.; Moreau, J.; Manière, C.; Thiéry, D.; Delbac, L.; Louâpre, P. Where You Come from Matters: Temperature Influences Host–Parasitoid Interaction through Parental Effects. Oecologia 2020, 192, 853–863. [Google Scholar] [CrossRef] [PubMed]
- da Costa-Lima, T.C.; de Araújo, A.T.P.; Torris, A.F. Biology and Population Dynamics of the American Vine Moth and the Potential Biocontrol with Trichogramma pretiosum. Neotrop. Entomol. 2021, 50, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Thiéry, D.; Louâpre, P.; Muneret, L.; Rusch, A.; Sentenac, G.; Vogelweith, F.; Iltis, C.; Moreau, J. Biological Protection against Grape Berry Moths. A Review. Agron. Sustain. Dev. 2018, 38, 1–18. [Google Scholar] [CrossRef]
- Bărbuceanu, D.; Bărbuceanu, M.; Andriescu, I. Parasitoids of Sparganothis pilleriana (Den. Et Schiff.) (Lepidoptera: Tortricidae) in Southern Vineyards of Romania. Curr. Trends Nat. Sci. 2020, 9, 307–316. [Google Scholar] [CrossRef]
- Moreau, J.; Villemant, C.; Benrey, B.; Thiéry, D. Species Diversity of Larval Parasitoids of the European Grapevine Moth (Lobesia botrana, Lepidoptera: Tortricidae): The Influence of Region and Cultivar. Biol. Control 2010, 54, 300–306. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Educaton and Data Anlysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Lester, P.J.; Beggs, J.R. Invasion Success and Management Strategies for Social Vespula Wasps. Annu. Rev. Entomol. 2019, 64, 51–71. [Google Scholar] [CrossRef]
- Beranová, V.; Abrahám, R.; Matečný, I.; Beracko, P.; Milics, G. Impact of Environmental Conditions on the Distribution of Insect Pests in Nitra Region Vineyards (Slovakia). Available online: https://uge-share.science.upjs.sk/webshared/GCass_web_files/articles/GC-2022-16-2/04_Beranova_et_al.pdf (accessed on 15 November 2023).
- Valentini, B.; Barbero, F.; Casacci, L.P.; Luganini, A.; Stefanini, I. Forests Influence Yeast Populations Vectored by Insects into Vineyards. Front. Microbiol. 2022, 13, 1039939. [Google Scholar] [CrossRef]
- Stefanini, I.; Dapporto, L.; Legras, J.-L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S. Role of Social Wasps in Saccharomyces Cerevisiae Ecology and Evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef]
- Rodríguez-Gasol, N.; Alins, G.; Veronesi, E.R.; Wratten, S. The Ecology of Predatory Hoverflies as Ecosystem-Service Providers in Agricultural Systems. Biol. Control 2020, 151, 104405. [Google Scholar] [CrossRef]
- Suvák, M. Predatory and Parasitic Insects in Greenhouses of Botanical Garden of PJ Šafárik University in Košice, Slovakia. Thaiszia–J. Bot. Košice 2011, 21, 185–205. [Google Scholar]
- Crowley, L.M.; Pointon, D.-L.; Darwin Tree of Life Consortium. of L. The Genome Sequence of a Beetle-Killing Wasp, Tiphia femorata (Fabricius, 1775). Wellcome Open Res. 2023, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Vlug, H. Rozekeverdolkwesp (Tiphia femorata F.). Bzzz/HymenoVaria 2011, 2, 13–14. [Google Scholar]
- Wood, T.J.; Holland, J.M.; Goulson, D. Diet Characterisation of Solitary Bees on Farmland: Dietary Specialisation Predicts Rarity. Biodivers. Conserv. 2016, 25, 2655–2671. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A.; Ponti, L.; Nicholls, C.I. Manipulating Vineyard Biodiversity for Improved Insect Pest Management: Case Studies from Northern California. Int. J. Biodivers. Sci. Manag. 2005, 1, 191–203. [Google Scholar] [CrossRef]
- Santini, L.; Belmaker, J.; Costello, M.J.; Pereira, H.M.; Rossberg, A.G.; Schipper, A.M.; Ceaușu, S.; Dornelas, M.; Hilbers, J.P.; Hortal, J. Assessing the Suitability of Diversity Metrics to Detect Biodiversity Change. Biol. Conserv. 2017, 213, 341–350. [Google Scholar] [CrossRef]
- Nagendra, H. Opposite Trends in Response for the Shannon and Simpson Indices of Landscape Diversity. Appl. Geogr. 2002, 22, 175–186. [Google Scholar] [CrossRef]
- Gamito, S. Caution Is Needed When Applying Margalef Diversity Index. Ecol. Indic. 2010, 10, 550–551. [Google Scholar] [CrossRef]
- Janković, M.; Nedeljković, Z.; Milovac, Ž.; Vujić, A. Diversity and Structure of Hoverfly (Diptera: Syrphidae) Communities in Agricultural Areas in Vojvodina Province (Serbia) a Case Study on Brassica napus L. J. Entomol. Res. Soc. 2019, 21, 129–144. [Google Scholar]
- Robert, M.M. Patterns of Species Abundance and Diversity. Ecol. Evol. Communities 1979, 40, 81–120. [Google Scholar] [CrossRef]
- Berger, W.H.; Parker, F.L. Diversity of Planktonic Foraminifera in Deep-Sea Sediments. Science 1970, 168, 1345–1347. [Google Scholar] [CrossRef] [PubMed]
- Caruso, T.; Pigino, G.; Bernini, F.; Bargagli, R.; Migliorini, M. The Berger–Parker Index as an Effective Tool for Monitoring the Biodiversity of Disturbed Soils: A Case Study on Mediterranean Oribatid (Acari: Oribatida) Assemblages. Biodivers. Conserv. Eur. 2007, 16, 3277–3285. [Google Scholar] [CrossRef]
- Beaumelle, L.; Auriol, A.; Grasset, M.; Pavy, A.; Thiéry, D.; Rusch, A. Benefits of Increased Cover Crop Diversity for Predators and Biological Pest Control Depend on the Landscape Context. Ecol. Solut. Evid. 2021, 2, e12086. [Google Scholar] [CrossRef]
- Altimira, F.; De La Barra, N.; Rebufel, P.; Soto, S.; Soto, R.; Estay, P.; Vitta, N.; Tapia, E. Potential Biological Control of the Pupal Stage of the European Grapevine Moth Lobesia botrana by the Entomopathogenic Fungus Beauveria pseudobassiana in the Winter Season in Chile. BMC Res. Notes 2019, 12, 548. [Google Scholar] [CrossRef] [PubMed]
- Altimira, F.; De La Barra, N.; Godoy, P.; Roa, J.; Godoy, S.; Vitta, N.; Tapia, E. Lobesia botrana: A Biological Control Approach with a Biopesticide Based on Entomopathogenic Fungi in the Winter Season in Chile. Insects 2021, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Picciotti, U.; Araujo Dalbon, V.; Ciancio, A.; Colagiero, M.; Cozzi, G.; De Bellis, L.; Finetti-Sialer, M.M.; Greco, D.; Ippolito, A.; Lahbib, N. “Ectomosphere”: Insects and Microorganism Interactions. Microorganisms 2023, 11, 440. [Google Scholar] [CrossRef]
- Vicente-Díez, I.; Blanco-Pérez, R.; Chelkha, M.; Puelles, M.; Pou, A.; Campos-Herrera, R. Exploring the Use of Entomopathogenic Nematodes and the Natural Products Derived from Their Symbiotic Bacteria to Control the Grapevine Moth, Lobesia botrana (Lepidoptera: Tortricidae). Insects 2021, 12, 1033. [Google Scholar] [CrossRef]
- Hommay, G.; Gertz, C.; Kienlen, J.C.; Pizzol, J.; Chavigny, P. Comparison between the Control Efficacy of Trichogramma evanescens Westwood (Hymenoptera: Trichogrammatidae) and Two Trichogramma cacoeciae Marchal Strains against Grapevine Moth (Lobesia botrana Den. & Schiff.), Depending on Their Release Density. Biocontrol Sci. Technol. 2002, 12, 569–581. [Google Scholar] [CrossRef]
- Larsson Herrera, S.; Rikk, P.; Köblös, G.; Szelényi, M.O.; Molnár, B.P.; Dekker, T.; Tasin, M. Designing a Species-Selective Lure Based on Microbial Volatiles to Target Lobesia botrana. Sci. Rep. 2020, 10, 6512. [Google Scholar] [CrossRef]
- Adams, M.R. Choosing Hunting Sites: Web Site Preferences of the Orb Weaver Spider, Neoscona crucifera, Relative to Light Cues. J. Insect Behav. 2000, 13, 299–305. [Google Scholar] [CrossRef]
- Martinez, J.I.; Crane, J.H.; Wasielewski, J.; Miller, J.Y.; Carrillo, D. Lepidoptera Pests of Sapodilla (Manilkara zapota (L.) van Royen) in South Florida, with Some Comments on Life History and Natural Control. 2019. Available online: https://digitalcommons.unl.edu/insectamundi/1234/ (accessed on 15 November 2023).
- Mishra, R.; Ahmad, G.; Chaubey, S.N. Study on the Morphology, Feeding Capcity and Prey Preference of Neoscona crucifera and N. adianta (Orb-Weaving Spiders). Indian J. Life Sci. 2012, 1, 29. [Google Scholar]
- Corrigan, J.E.; Bennett, R.G. Predation by Cheiracanthium mildei (Araneae, Clubionidae) on Larval Phyllonorycter blancardella (Lepidoptera, Gracillariidae) in a Greenhouse. J. Arachnol. 1987, 15, 132–134. [Google Scholar]
- Yanbing, L.I.; Adalbieke, N.; Wumuerhan, P.; Deying, M.A. Effects of Artificial Migration of Harmonia axyridis and Chemical Control on the Control Effect of Cotton Aphid and the Population Dynamics of Natural Enemies. Xinjiang Agric. Sci. 2022, 59, 1173. [Google Scholar] [CrossRef]
- EPPO. Available online: https://gd.eppo.int/taxon/PSYBVI (accessed on 27 March 2023).
- Sutherland, A.M.; Parrella, M.P. Mycophagy in Coccinellidae: Review and Synthesis. Biol. Control 2009, 51, 284–293. [Google Scholar] [CrossRef]
- Ratti, E. Coleoptera Associated with Oidium evonymi-japonici (Arc.) Sacc.(Fungi Imperfecti) in the Urban Gardens of Venice. Boll. Mus. Civ. Stor. Nat. Venezia 1996, 45, 47–51. [Google Scholar]
- Karataraki, K.; Goumenaki, E.; Raftakis, E.; Goutos, D.; Kapetanakis, E. First Record of the Mycophagous Ladybird Psyllobora vigintiduopunctata on Greenhouse Cucumber Plants in Crete (Greece). Entomol. Hell. 2015, 24, 37–41. [Google Scholar] [CrossRef]
- Poor, A.M.; Jafari, R.; Biranvand, A.; Zare, M.; Karahrudi, Z.R. Ladybirds Associated with Pomegranate Trees in Lorestan Province of Iran (Coleoptera: Coccinellidae). Int. Res. J. Appl. Basic Sci. 2013, 5, 1585–1589. [Google Scholar]
- Tyumaseva, Z.I.; Guskova, E.V. Trophic Relations of Lady Beetles (Coleoptera, Coccinellidae) of the Urals. Ukr. J. Ecol. 2016, 6, 61–66. [Google Scholar] [CrossRef]
- Younes, G.H.; Ahmad, M.; Ali, N. Morphological, Biological and Ecological Studies of the Mycophagous Ladybird Psyllobora vigintiduopunctata L.(Coleoptera: Coccinellidae) on Powdery Mildew Fungi in the Coastal Region of Syria. Jordan J. Agric. Sci. 2015, 11, 483–494. [Google Scholar]
- Dillen, M.; Smit, C.; Buyse, M.; Höfte, M.; De Clercq, P.; Verheyen, K. Stronger Diversity Effects with Increased Environmental Stress: A Study of Multitrophic Interactions between Oak, Powdery Mildew and Ladybirds. PLoS ONE 2017, 12, e0176104. [Google Scholar] [CrossRef] [PubMed]
- Başar, M.K.; Yaşar, B. Determination of Ladybird Species (Coleoptera: Coccinellidae) on Fruit Trees in Isparta, Turkey. Turk. J. Entomol. 2011, 35, 519–534. [Google Scholar]
- Elekçioğlu, N.Z.; Şenal, D. Pest and Natural Enemy Fauna in Organic Citrus Production in the Eastern Mediterranean Region of Turkey. Int. J. Nat. Eng. Sci. 2007, 1, 29–34. [Google Scholar]
- Ölmez, S. Diyarbakır Ilinde Aphidoidea (Homoptera) Türleri Ile Bunların Parazitoit ve Predatörlerinin Saptanması. Ph.D. Dissertation, Çukurova University, Adana, Türkiye, 2000. [Google Scholar]
- UK Beetles. Available online: https://www.ukbeetles.co.uk/anisosticta-novem (accessed on 27 March 2023).
- Tinsley, M.C.; Majerus, M.E.N. A New Male-Killing Parasitism: Spiroplasma bacteria Infect the Ladybird Beetle Anisosticta novemdecimpunctata (Coleoptera: Coccinellidae). Parasitology 2006, 132, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Radonjic, A.; Terenius, O.; Ninkovic, V. The Phytopathogen Powdery Mildew Affects Food-Searching Behavior and Survival of Coccinella septempunctata. Arthropod Plant Interact. 2018, 12, 685–690. [Google Scholar] [CrossRef]
- EPPO. Aleurocanthus citriperdus, Aleurocanthus spiniferus and Aleurocanthus woglumi. EPPO Bull. 2022, 52, 346–361. [Google Scholar] [CrossRef]
- Suga, H.; Hirano, W.; Katoh, T.; Matsubayashi, K.W.; Katakura, H. Introgression and Habitat Segregation in a Pair of Ladybird Beetle Species in the Genus Propylea (Coccinellidae, Coccinellinae) in Northern Japan. Zool. Sci. 2016, 33, 603–610. [Google Scholar] [CrossRef]
- Kolibáč, J.; Perkovsky, E.E. A Reclassification of Acanthocnemoides Sukatshevae Zherikhin, 1977 from the Mid-Cretaceous Taimyr amber (Coleoptera). Cretac. Res. 2020, 115, 104548. [Google Scholar] [CrossRef]
- Nonveiller, G. Predators of Gypsy Moth Eggs Found in Yugoslavia during the 1945–1950 Gypsy Moth Plague. Sel. Artic. Plant Prot. 1950, 10, 15–35. [Google Scholar]
- Maican, S.; Munteanu, C. The Diversity of Urban Coleopterans. In Species Monitoring in the Central Parks of Bucharest; Onete, M., Ed.; Ars Docendi: Bucharest, Romania, 2008; pp. 106–112. ISBN 978-973-558-392-7. [Google Scholar]
- Crowley, L.M.; University of Oxford; Wytham Woods Genome Acquisition Lab; Wellcome Sanger Institute Tree of Life; Darwin Tree of Life Consortium. The Genome Sequence of the Common Malachite Beetle, Malachius bipustulatus (Linnaeus, 1758). Wellcome Open Res. 2021, 6, 322. [Google Scholar] [CrossRef]
- Gutowski, J.M. Pollination of the Orchid Dactylorhiza fuchsii by Longhorn Beetles in Primeval Forests of Northeastern Poland. Biol. Conserv. 1990, 51, 287–297. [Google Scholar] [CrossRef]
- Malschi, D. Actual Aspect Regarding the Control of Cereal Leaf Beetles (Oulema melanopus L.) in Central Transylvania. Probl. Proteçtia Plantelor 2000, 28, 17–28. [Google Scholar]
- Rather, Z.A.; Ollerton, J.; Parey, S.H.; Ara, S.; Watts, S.; Paray, M.A.; Khuroo, A.A. Plant-Pollinator Meta-Network of the Kashmir Himalaya: Structure, Modularity, Integration of Alien Species and Extinction Simulation. Flora 2023, 298, 152197. [Google Scholar] [CrossRef]
- Sahebari, F.S.; Khaghaninia, S.; Talebi, A.A. New Data of the Subfamily Tachininae (Diptera: Tachinidae) from North-Western Iran. Zool. Ecol. 2018, 28, 252–258. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An International Database for Pesticide Risk Assessments and Management. Hum. Ecol. Risk Assess. Int. J. 2016, 22, 1050–1064. [Google Scholar] [CrossRef]
- Grant, J.F.; Shepard, M. Biological Characteristics of a South American Population of Voria ruralis (Diptera: Tachinidae), a Larval Parasitoid of the Soybean looper (Lepidoptera: Noctuidae). Environ. Entomol. 1983, 12, 1673–1677. [Google Scholar] [CrossRef]
- Cambero Campos, O.J.; Rios Velasco, C.; Gallegos Morales, G.; Del Rincon Castro, M.A.C.; Ulloa Rubio, K.; Valenzuela Garcia, R.D. First Record of Archytas marmoratus and Voria ruralis (Diptera Tachinidae) and Levels of Parasitoidism In Two Lepidoptera Pest in Coahuila Mexico. Acta Zool. Mex. 2011, 27, 577–582. [Google Scholar]
- Rajan, S.J.; ES-Sathish, R. Biology of Big-Eyed Bug, Geocoris erythrocephalus (Lepeletier & Serville) on Cabbage Aphid, Brevicoryne brassicae (L.). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 3301–3305. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Parra, J.R.P. Insect Bioecology and Nutrition for Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2012; ISBN 1439837082. [Google Scholar] [CrossRef]
- Jerinić-Prodanović, D. Diverzitet Lisnih Buva (Homoptera: Psylloidea) i Njihovih Prirodnih Neprijatelja u Srbiji, Sa Posebnim Osvrtom Na Vrste Značajne u Poljoprivredi. Ph.D. Thesis, Faculty of Agriculture, University of Belgrade, Belgrade, Serbia, 2010; 339p. [Google Scholar] [CrossRef]
- Jerinić-Prodanović, D.; Protić, L. True Bugs (Hemiptera, Heteroptera) as Psyllid Predators (Hemiptera, Psylloidea). Zookeys 2013, 319, 169–189. [Google Scholar] [CrossRef]
- Harizanova, V.; Stoeva, A.; Mohamedova, M. Preliminary Study on the Invasive Acizzia jamatonica (Hemiptera: Psyllidae) and Its Predators in Bulgaria. Agric. Sci. Technol. 2012, 4, 56–61. [Google Scholar]
- Mendes, S.M.; Bueno, V.H.P. Biologia de Orius insidiosus (Say)(Hemiptera: Anthocoridae) Alimentado Com Caliothrips Phaseoli (Hood)(Thysanoptera: Thripidae). Neotrop. Entomol. 2001, 30, 423–428. [Google Scholar] [CrossRef]
- Lorenzo, M.E.; Bao, L.; Mendez, L.; Grille, G.; Bonato, O.; Basso, C. Preference of Orius insidiosus and Orius tristicolor (Hemiptera: Anthocoridae) for Host Plants in Olfactometry and Free-Choice Experiments. Fla. Entomol. 2021, 103, 492–498. [Google Scholar] [CrossRef]
- Lorenzo, M.E.; Bao, L.; Grille, G.; Mendez, L.; Bonato, O.; Basso, C. Compatibility of Orius insidiosus (Hemiptera: Anthocoridae) with Amblyseius swirskii (Acari: Phytoseiidae) for Control of Flankliniella occidentalis (Thysanoptera: Thripidae) in Pepper. Agrociencia Urug. 2021, 25, e838. [Google Scholar] [CrossRef]
- Paul, K.; Khan, A. Behaviour of Orius insidiosus Say (Hemiptera: Anthocoridae) towards Its Prey, Thrips palmi Karny (Thysanoptera: Thripidae). J. Biol. Control 2020, 34, 298–302. [Google Scholar] [CrossRef]
- Baez, I.; Reitz, S.R.; Funderburk, J.E. Predation by Orius insidiosus (Heteroptera: Anthocoridae) on Life Stages and Species of Frankliniella Flower Thrips (Thysanoptera: Thripidae) in Pepper Flowers. Environ. Entomol. 2004, 33, 662–670. [Google Scholar] [CrossRef]
- Krivosheina, M.G. To the Knowledge of Insects, Pollinators of Sorbus aucuparia Linnaeus, 1753 and Aronia mitschurinii AK Skvortsov et Maitul., 1982 in Moscow Region, Russia. Cauc. Entomol. Bull. 2018, 14, 105–111. [Google Scholar]
- Wood, T.J.; Roberts, S.P.M. An Assessment of Historical and Contemporary Diet Breadth in Polylectic andrena Bee Species. Biol. Conserv. 2017, 215, 72–80. [Google Scholar] [CrossRef]
- Dar, S.A.; Sofi, M.A.; El-Sharnouby, M.; Hassan, M.; Rashid, R.; Mir, S.H.; Al Naggar, Y.; Salah, M.; Gajger, I.T.; Sayed, S. Nesting Behaviour and Foraging Characteristics of Andrena cineraria (Hymenoptera: Andrenidae). Saudi J. Biol. Sci. 2021, 28, 4147–4154. [Google Scholar] [CrossRef]
- Antonelli, A.; Johan Dahlberg, C.; Carlgren, K.H.I.; Appelqvist, T. Pollination of the Lady’s Slipper Orchid (Cypripedium calceolus) in Scandinavia–Taxonomic and Conservational Aspects. Nord. J. Bot. 2009, 27, 266–273. [Google Scholar] [CrossRef]
- Cane, J.H.; Sampson, B.J.; Miller, S.A. Pollination Value of Male Bees: The Specialist Bee Peponapis pruinosa (Apidae) at Summer Squash (Cucurbita pepo). Environ. Entomol. 2011, 40, 614–620. [Google Scholar] [CrossRef]
- Hung, K.-L.J.; Kingston, J.M.; Albrecht, M.; Holway, D.A.; Kohn, J.R. The Worldwide Importance of Honey Bees as Pollinators in Natural Habitats. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172140. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Kardum Hjort, C.; Paris, J.R.; Olsson, P.; Herbertsson, L.; de Miranda, J.R.; Dudaniec, R.Y.; Smith, H.G. Genomic Divergence and a Lack of Recent Introgression between Commercial and Wild Bumblebees (Bombus terrestris). Evol. Appl. 2022, 15, 365–382. [Google Scholar] [CrossRef] [PubMed]
- Giannini, T.C.; Lira-Saade, R.; Ayala, R.; Saraiva, A.M.; Alves-dos-Santos, I. Ecological Niche Similarities of Peponapis Bees and Non-Domesticated Cucurbita Species. Ecol. Model. 2011, 222, 2011–2018. [Google Scholar] [CrossRef]
- Brand, N. Social Evolution in the Sweat Bee Halictus scabiosae. Ph.D. Dissertation, Faculté de Biologie et Médecine, Université de Lausanne, Lausanne, France, 2014. [Google Scholar]
- Bogusch, P. Vespoidea: Tiphiidae (Trněnkovití). Acta Entomol. Musei Natl. Pragae Suppl. 2007, 11, 85–92. Available online: https://www.aemnp.eu/data/article-1211/1192-11_0_85.pdf (accessed on 25 September 2023).
- Sauer, K.P.; Vermeulen, A.; Aumann, N. Temperature-dependent Competition Hierarchy: A Mechanism Stabilizing the Phenological Strategy in the Scorpionfly Panorpa communis L. J. Zool. Syst. Evol. Res. 2003, 41, 109–117. [Google Scholar] [CrossRef]
- Burrows, M. Jumping and Take-off in a Winged Scorpion Fly (Mecoptera, Panorpa communis). J. Exp. Biol. 2019, 222, jeb205385. [Google Scholar] [CrossRef]
- Rehman, H.; Bukero, A.; Lanjar, A.G.; Bashir, L.; Lanjar, Z.; Nahiyoon, S.A. Use of Chrysoperla carnea Larvae to Control Whitefly (Aleyrodidea: Hemiptera) on Tomato Plant in Greenhouse. Pure Appl. Biol. (PAB) 2020, 9, 2128–2137. [Google Scholar] [CrossRef]
- Usman, M.; Inayatullah, M.; Usman, A.; Sohail, K.; Shah, S.F. Effect of Egg Parasitoid, Trichogramma chilonis, in Combination with Chrysoperla carnea and Neem Seed Extract against Tomato Fruitworm, Helicoverpa Armigera. Sarhad J. Agric 2012, 28, 253–257. [Google Scholar]
- Hagley, E.A.C. Release of Chrysoperla carnea Stephens (Neuroptera: Chrysopidae) for Control of the Green Apple Aphid, Aphis Pomi DeGeer (Homoptera: Aphididae). Can. Entomol. 1989, 121, 309–314. [Google Scholar] [CrossRef]
- Khfif, K.; Labaioui, Z.; Koledenkova, K.; Zaid, A.; El Rhaffari, L.; Brostaux, Y. Predatory Performance of Chrysoperla carnea (Neuroptera: Chrysopidae) on Jacobiasca lybica (Hemiptera: Cicadellidae) under Laboratory Conditions. J. Agric. Urban Entomol. 2023, 39, 1–8. [Google Scholar] [CrossRef]
- Pinzón-Hamón, D.A.; Martínez-Osorio, J.W.; Castro-López, M.A. Parasitism of Trichogramma and Prédation of Chrysoperla on Compsus viridivittatus Eggs, a Vitis vinifera Pest, under Laboratory Conditions. Rev. Colomb. Cienc. Hortícolas 2018, 12, 348–357. [Google Scholar] [CrossRef]







| Year | Crop | No. of Species | Individuals | Biodiversity Index | ||||
|---|---|---|---|---|---|---|---|---|
| Shannon–Wiener | Simpson | Evenness | Margalef | Berger-Parker | ||||
| 2020 | NPG | 31 | 588 | 2.636 | 0.8831 | 0.4501 | 4.705 | 0.2517 |
| GNPG | 47 | 878 | 2.533 | 0.86 | 0.2678 | 6.787 | 0.3007 | |
| BF | 39 | 1796 | 1.566 | 0.5652 | 0.1227 | 5.071 | 0.6487 | |
| GBF | 52 | 539 | 3.032 | 0.9267 | 0.3986 | 8.108 | 0.141 | |
| 2021 | NPG | 65 | 2134 | 1.935 | 0.6085 | 0.1066 | 8.349 | 0.6195 |
| GNPG | 57 | 798 | 2.603 | 0.8044 | 0.2368 | 8.381 | 0.4211 | |
| BF | 62 | 705 | 2.939 | 0.8758 | 0.3047 | 9.301 | 0.3007 | |
| GBF | 58 | 690 | 3.182 | 0.933 | 0.4152 | 8.72 | 0.1348 | |
| 2022 | NPG | 38 | 539 | 2.258 | 0.7531 | 0.2517 | 5.883 | 0.4768 |
| GNPG | 38 | 1190 | 1.467 | 0.5109 | 0.1141 | 5.225 | 0.6924 | |
| BF | 51 | 1057 | 2.793 | 0.8625 | 0.3201 | 7.181 | 0.3359 | |
| GBF | 49 | 876 | 2.823 | 0.9048 | 0.3434 | 7.084 | 0.2158 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vizitiu, D.E.; Sardarescu, I.-D.; Buciumeanu, E.C.; Guta, I.-C.; Dincă, L.; Bălăcenoiu, F.; Toma, D.; Crișan, V.; Din, A. The Influence of Groves on Aboveground Arthropod Diversity and Evolution in a Vineyard in Southern Romania. Sustainability 2023, 15, 16543. https://doi.org/10.3390/su152316543
Vizitiu DE, Sardarescu I-D, Buciumeanu EC, Guta I-C, Dincă L, Bălăcenoiu F, Toma D, Crișan V, Din A. The Influence of Groves on Aboveground Arthropod Diversity and Evolution in a Vineyard in Southern Romania. Sustainability. 2023; 15(23):16543. https://doi.org/10.3390/su152316543
Chicago/Turabian StyleVizitiu, Diana Elena, Ionela-Daniela Sardarescu, Elena Cocuta Buciumeanu, Ionela-Cătălina Guta, Lucian Dincă, Flavius Bălăcenoiu, Dragoș Toma, Vlad Crișan, and Alin Din. 2023. "The Influence of Groves on Aboveground Arthropod Diversity and Evolution in a Vineyard in Southern Romania" Sustainability 15, no. 23: 16543. https://doi.org/10.3390/su152316543