3.2. Screening of Fungal Strains for Lipid Production
Fungal strains were initially screened on PKC for their ability to grow and accumulate intracellular lipids during a 15-day cultivation period.
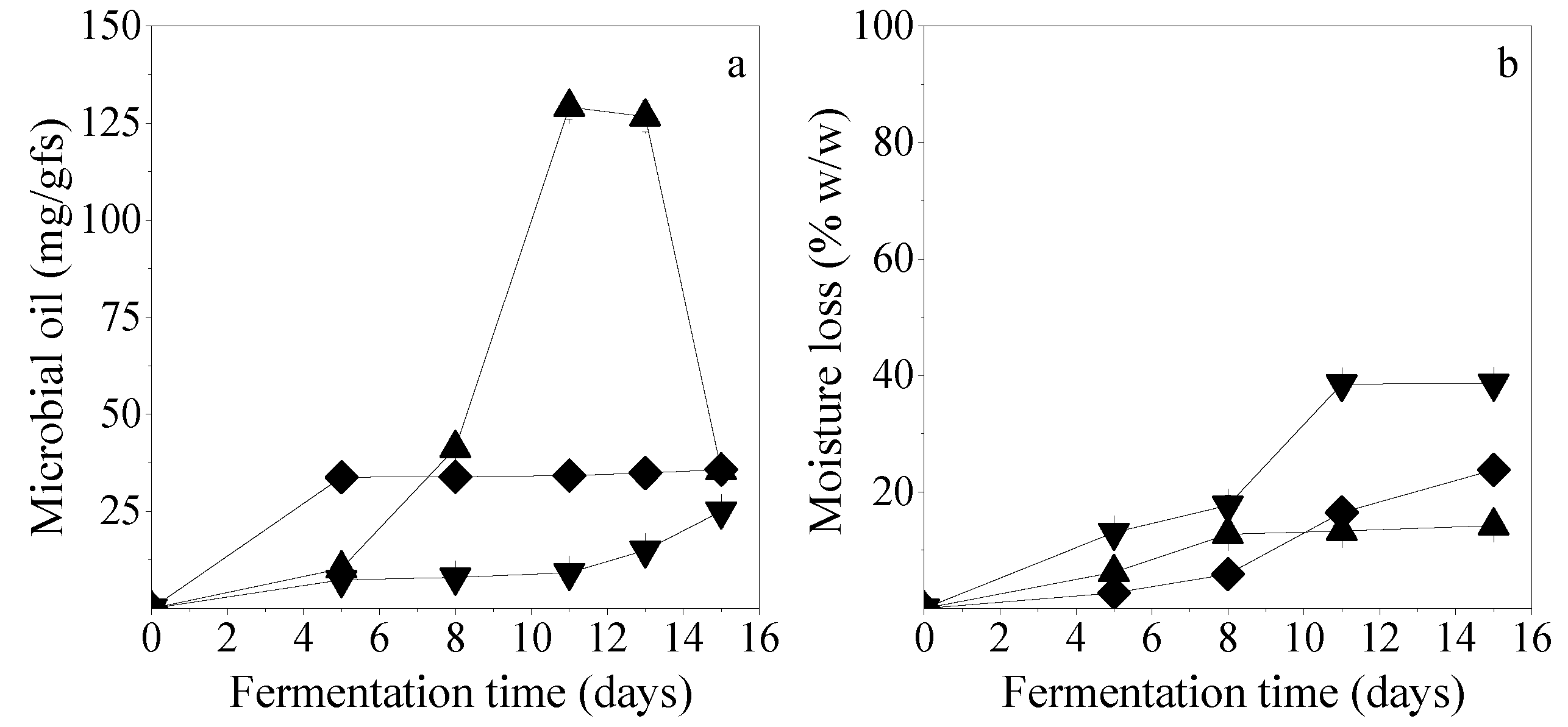
Figure 1a presents the microbial oil production of
C. echinulata,
M. ramanniana MUCL, and
M. ramanniana ATHUM obtained in SSF with 65% initial moisture content in non-humidified chambers. The maximum microbial oil production was determined for
C. echinulata, reaching 129.1 mg/gfs after 11 days of fermentation. In this case, the maximum productivity (11.7 mg/gfs/day) was also observed. After 15-days, cultures of
C. echinulata showed enhanced sporulation tendency, due to nutrient depletion, while lipid concentration was reduced to 35.5 mg/gfs.
M. ramanniana MUCL and ATHUM strains presented lower lipid production capacity, reaching up to 33.8 mg/gfs (after 5 days of fermentation) and 25.0 mg/gfs (15 days of fermentation), respectively. Statistically significant differences (
p < 0.05) were detected in lipids produced by
C. echinulata at 11 days of fermentation,
M. ramanniana MUCL at 5 days of fermentation, and
M. ramanniana ATHUM at 15 days of fermentation. Tukey test revealed significant differences in all possible combinations.
T. elegans and
M. isabellina displayed a lipolytic tendency (data not presented in
Figure 1), resulting in the depletion of the initial oil content of PKC, by 94.2% (after 15 days) and 62.2% (after 11 days), respectively. In these cases, the biotransformation of the initial PKC oil to fungal microbial oil was not possible to determine. This observation suggests that these two strains used the lipids contained in PKC as an energy source for growth, probably owing to their inability to utilize other carbon and nutrient sources of the growth medium. Similar phenomena have been reported by Ferreiera et al. [
39] who performed SSF with
M. aplina and verified that the content of microbial oil in the fermented rapeseed cake, sunflower cake, and soybean cake was lower compared to the unfermented ones.
When SSF was carried out using
M. isabellina, moisture losses were higher than 50% whereas the minimum moisture loss (14%) was determined in the case that SSF was performed with
C. echinulata (
Figure 1b). These results indicated that the higher lipid production was positively related to lower moisture losses. This may be attributed to the higher mycelium growth and therefore lipid accumulation, of
C. echinulata,
M. ramanniana MUCL and
M. ramanniana ATHUM than the other fungal strains, which was based on a macroscopic observation at this stage of the study.
The synthesis of microbial oil is a bioprocess that is highly affected by numerous macro- and micro-factors of the fermentation environment including inoculum size, temperature, substrate’s supplementation with nutrients, initial pH, and pretreatment strategy. The specificity and affinity of the microbial strains (reflected by their active enzymatic complexes and their ability to catalyze targeted molecules and processes) to the fermentation matrix can lead to variations in the production of microbial oil. For example, Stredansky et al. [
19] screened twenty-three strains of the genera
Mortierella, Cunninghamella, Rhizopus, Mucor, and
Thamnidium for lipid production on apple pomace and spent malt grains impregnated with a nutrient solution.
Cunninghamella strains produced lipids varying between 150–166 mg/gfs during 8 days of cultivation while
M. isabellina, reached a lipid content of 168 mg/gfs. Slightly higher lipids production (172 mg/gfs) was achieved with
T. elegans. In another study, several fungal strains of the order
Mucorales were screened for lipid production on pearl barley.
Cunninghamella strains showed higher ability to produce microbial oil (141–158 mg/gfs) compared to
Mortierella strains (106–121 mg/gfs). Lower microbial oil production (47.9 mg/gfs) was achieved when
M. isabellina was cultivated on soybean hull applying an inoculums size of 104/g soybean hull, 75% initial humidity, and 7-day spores without nutrient addition [
21].
Table 2 depicts the fatty acid composition of unfermented PKC oil and fermented PKC by the aforementioned fungal strains. PKC oil mainly consisted of lauric acid (C12:0) (47.7%), myristic acid (C14:0) (15.3%), C16:0 (9.3%), and C18:1 (15.1%) with a RSU equal to 4.7. At the end of SSF, all fungal strains were able to transform the fatty acid composition of unfermented PKC oil, reducing the RSU within the range of 0.8–1.5. It is remarkable that C12:0 was reduced to 13.1–27.7%, while C18:1 and C16:0 were substantially increased, varying within 27.6–34.9% and 12.2–17.3%, respectively. The highest γ-linolenic acid (GLA) (C18:3) content was observed in lipids produced by
C. echinulata (5.3%), followed by
M. ramanniana MUCL (2.7%),
M. isabellina (1.7%), and
M. ramanniana ATHUM (0.8%). In the case of
T. elegans, GLA was not detected. Evidently, the increased production of C18:1 may be attributed mostly to de novo lipogenesis rather than the partial saturation of linoleic acid (C18:2), since C18:2 was also increased throughout the fermentation. Higher GLA contents have been reported for
M. isabellina (4.2–5.3%) when SSF was implemented on pearl barley or mixtures of apple pomace and spent malt grains supplemented with nutrients [
19,
20]. GLA production in lipids derived from
Cunninghamella strains has been determined up to 11.6% [
20] while significant GLA amounts (7.5–9.1%) have been reported for
T. elegans under solid-state cultivation [
19,
20].
Lower filamentous fungi are able to follow versatile metabolic pathways, leading to fatty acid biosynthesis and bioconversion processes, as pointed out in this section [
40]. As it was observed in
Table 2, the initial oil contained in unfermented PKC showed much higher RSU value compared to the fermented cake. The saturated fraction of PKC oil was 82.4% with respect to C8:0, C10:0, C12:0, C14:0, C16:0, and C18:0. Decreasing the RSU ratio of several discarded and low-value cakes applying fungi can be a very interesting strategy to obtain lipids rich in mono-unsaturated and poly-unsaturated fatty acids. Cakes enriched in unsaturated fatty acids could be directly supplied in animal diets, with several health benefits such as enhanced milk production, decrease of the saturated lipids in cheese, etc. [
41]. Alternatively, unsaturated lipids (low RSU ratios) could be extracted from these cakes and further integrated in food and cosmetics formulations, as they function as key compounds of antimicrobial food additives preventing the growth of undesired microorganisms. Both C18:1 and C18:2 act synergistically as an important antibacterial agent in herbs [
42]. Unsaturated lipids are also valuable for the chemical industry to produce numerous oleochemicals [
43]. Novel and sustainable enzymatic and biotechnological techniques have been recently employed by the modern organic chemistry to produce fatty acid-based chemicals [
44].
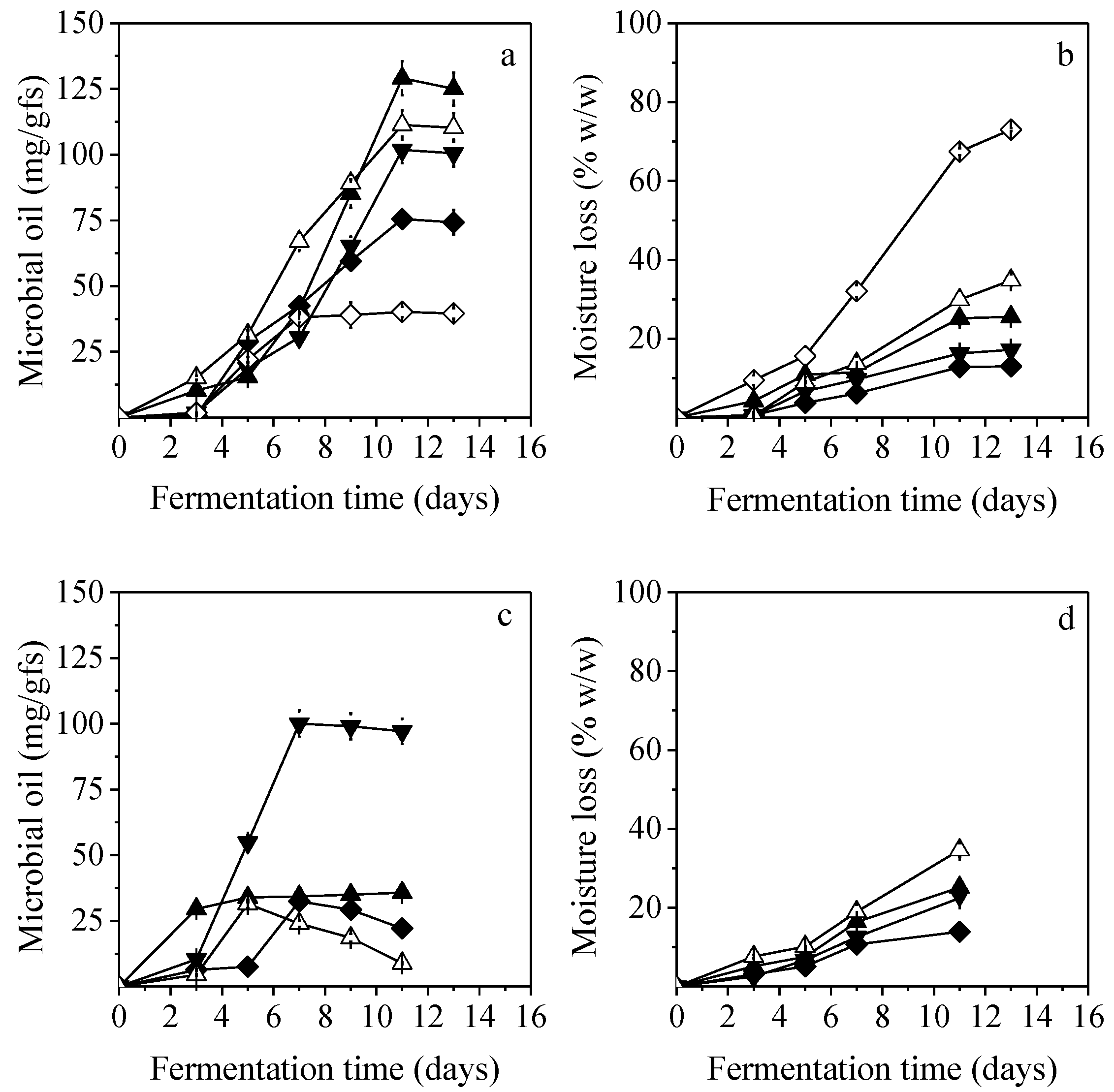
3.3. Effect of Temperature
C. echinulata and
M. ramanniana MUCL were the most efficient strains for lipid production among all fungal strains that were evaluated, and thus, they were selected for further investigation of the fermentation conditions (temperature). Results concerning the effect of temperature on lipid production are presented in
Figure 2. Different temperature regimes showed a significant effect (
p < 0.05) on lipids produced from
C. echinulata at 11-days of SSF. Tukey’s test demonstrated that solely data obtained from 27 °C and 30 °C did not differ significantly when compared between them. Low lipid production was achieved at 25 °C (75.5 mg/gfs) and 35 °C (40 mg/gfs) (after 11 days). The most effective temperature was 30 °C yielding a maximum lipid production of 129.1 mg/gfs after 11 days, while at 27 °C and 33 °C, lipid production was up to 111 mg/gfs (
Figure 2a).
In the case of
M. ramanniana MUCL, the most efficient temperature was 27 °C resulting in 100 mg/gfs of lipids after 7 days (
Figure 2c). Further increase or decrease in temperature resulted in a significant decrease of microbial lipids without significant differences (
p < 0.05) (regarding comparison of lipids production after 7 days of SSF at 25 °C, 30 °C, and 33 °C). Lipid production varied between 31.4 and 35.7 mg/g at 25 °C, 30 °C, and 33 °C without significant differences (
p < 0.05), whereas at 35 °C, only a 30.4% depletion of initial PKC oil content was observed. It should be stressed that an increase in temperature led eventually to increased moisture loss for both fungal strains (
Figure 2b,d).
Temperature constitutes an important variable for SSF performance as it directly affects the growth of microorganism and metabolites production. This is related to phenomena like protein denaturation, enzyme inhibition, and cell death [
13]. Temperature control is complicated when dealing with SSF due to its static nature and poor thermal conductivity of solids matrices [
13]. Although lipid production is a bioprocess that is affected by multiple factors, the results of this part of the study indicated that the temperature impact on lipid biosynthesis could be mostly related to the use of non-humidified chambers that led to fluctuations of the moisture content of the substrate.
Table 3 shows the effect of different temperature on the fatty acid profile of oil derived from fermented PKC after 11-days of SSF with
C. echinulata and
M. ramanniana MUCL. Temperatures of 25 °C and 35 °C did not favor the production of microbial oil when both fungal strains were applied, and thus, the ability of the fungi to biotransform the oil contained in the unfermented PKC was quite low. This was mainly reflected by the high content of C12:0 (40.1–42.9%) (
Table 3) that was detected in the fatty acids that was quite close to that of unfermented PKC (47.7%) (
Table 2).
Lipids derived from both fungal strains when fermentations were performed at 27 °C, 30 °C, and 33 °C demonstrated an increasing RSU as temperature was increased, while the lowest temperature regime of 27 °C displayed the lowest RSU (0.8–1.4). In the case of
C. echinulata, the increase in the unsaturation degree at 27 °C and 30 °C was mostly attributed to the increase of C18:1, C18:2, and C18:3 as well as the substantial decrease of C12:0 and C14:0 (compared to the oil of the fermented PKC and of the fermented PKC at 25 °C). A similar tendency was observed for lipids that derived from
M. ramanniana MUCL at a temperature regime of 27 °C. More specifically, in this case, the fatty acid profile of lipids was 2.7% GLA, 34.9% C18:1, and 18.0% C18:2, whereas C12:0 was reduced by 72.5%. The best performing fermentations in terms of total lipids production, resulted in the highest GLA production of 3.8–4.5% in the case of
C. echinulata and 2.7% in the case
M. ramanniana MUCL (
Table 3).
Fatty acids are regulators of membrane fluidity and permeability [
45]. Investigating the correlation between temperature and lipid saturation degree, it has been reported that increasing temperature values of fermentation tend to increase the saturation degree of fatty acids in microbial oil and decrease their average chain length [
46,
47]. Théberge et al. [
46] found that growth in temperature, obviously affected the fatty acid composition of free-living bacteria
Rhizobium leguminosarum bv.
viciae. More specifically, the percentage of saturated fatty acids, i.e., C16:0 and C18:0, increased with a decrease in temperature. In the case of unsaturated fatty acids, C18:l increased substantially by lowering the temperature. An unsaturated fatty acid (octadecenoic) varied from undetectable when 22 °C and 30 °C were applied to 14% at 10 °C and 15 °C. In another study,
Chlorella sorokiniana was cultivated at temperatures varying from 14 °C to 38 °C. Saturated fatty acids dominated at 38 °C with a percentage of 46%, while lower temperatures favored the formation of tri-unsaturated and di-unsaturated fatty acids [
47].
3.4. Effect of Initial Moisture Content
The effect of the initial moisture content of the substrate on lipid production by
C. echinulata (30 °C) and
M. ramanniana MUCL (27 °C) was studied in the range of 55–75% (
Figure 3). In the case of
C. echinulata, the ANOVA analysis (regarding the production of lipids at 11-days of SSF) showed that the initial moisture content of the solid substrate significantly affected lipids yield. Tukey test revealed significant differences at all possible combinations. The highest lipids production was achieved at 65% initial moisture content, followed by 108.8 mg/gfs at initial moisture content of 70%, after 11 days. Moisture levels lower than 65% were insufficient for the growth of
C. echinulata, resulting in the reduction of PKC oil content (of the initial substrate) by 71.9% and 79.8% at 55% and 60% initial moisture, respectively (data not presented in
Figure 3). In these cases, moisture losses were determined to be between 53 and 76% at the end of the fermentation. The highest initial moisture content of 75% led to the lowest microbial oil production and to the lowest moisture loss after 11 days of fermentation (
Figure 3a,b).
SSF of
M. ramanniana MUCL led to the maximum lipid production of 110.1 mg/gfs at 70% initial moisture content at 7 days of cultivation. Satisfactory lipid production (100.1 mg/gfs) was achieved at initial moisture contents of 65% while no statistically significant differences were detected. When the initial moisture content of 75% was applied, lipids production reached 93.8 mg/gfs at 7 days, with statistically significant differences, compared to data obtained from the initial moisture content of 70%. Initial moisture content of 60% resulted in a significant decrease (
p < 0.05) in lipid production (26.7 mg/gfs at 11 days of cultivation) when compared to data obtained from all the aforementioned moisture contents. The moisture level of 55% was insufficient for microbial lipids production, since only depletion of PKC oil content (27%) was determined. In all cases, moisture losses were found higher with decreased initial moisture contents (
Figure 3b,d).
The importance of water in SSF lies on the fact that most viable cells possess high moisture contents (70–80%). This study showed that a moisture content around 65–70%, depending on the microorganism, is required for maximum lipid production, since lower moisture levels are related to difficulties in nutrient diffusion, i.e., decreased the solubility of nutrients and microbial growth, while higher levels (>70%) led to substrate conglomeration, hindering of gas transfer phenomena, and bacterial contamination [
48].
Fatty acid composition of the 11-day fermented PKC (
Table 4) presented notable changes in comparison to the fatty acid composition of the unfermented PKC, for both fungal strains. A moisture level of 65% in the case of
C. echinulata enhanced the mono- and polyunsaturated fatty acid content of lipids. Particularly, fermented solids of
C. echinulata showed increased content of C18:1 (26.2%) and C18:2 (10.0%). C12:0 was decreased from 47.7% to 28.5% while GLA reached up to 4.5%. With respect to
M. ramanniana MUCL, initial moisture contents of 55% and 60% did not influence the fatty acid composition of oil (contained in the unfermented solids of PKC) while the produced GLA was rather low (0.5–0.7%). This indicated the very limited potential of the fungal strain to grow and produce lipids at relatively low moisture contents. At higher initial moisture levels, C16:0 and C18:1 contents were increased while C12:0 was significantly decreased. GLA formation reached its maximum value of 3.3% at initial moisture contents (70%) that favored lipids accumulation. Initial moisture contents within 70–75% have been reported to favor the production of ω−6 PUFAs [
49] while values within 65–75% have been proven optimal for total PUFAs production and C18:2 production. The initial moisture content of the solid matrix significantly affects the fatty acids proportion of lipids produced under SSF while the critical range that favors lipids production is narrow [
49,
50].
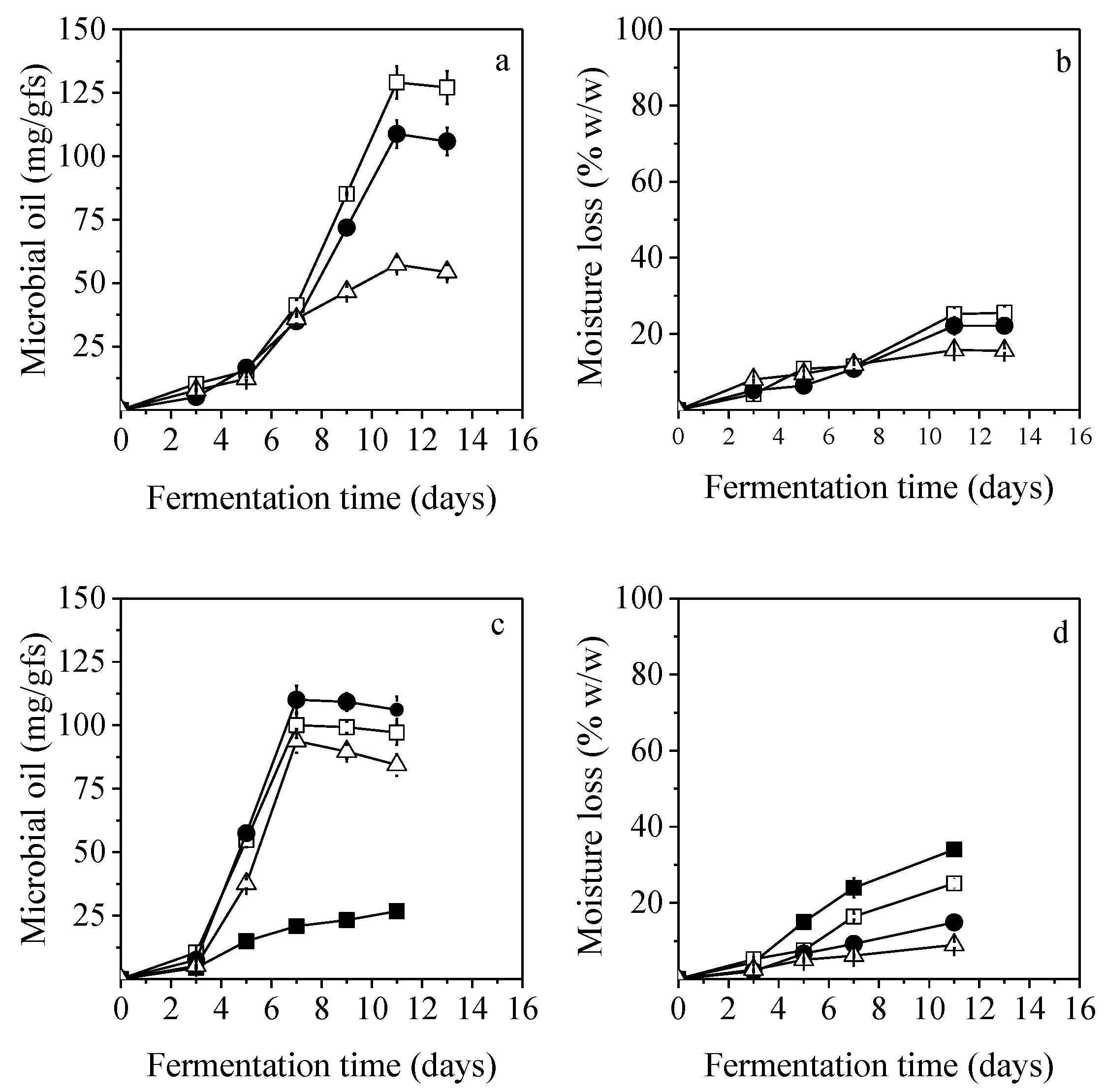
3.5. Effect of Different PKC to PPF Ratios
In an attempt to increase the surface area of SSF and its capacity to interchange with microbial growth and heat and mass transfer phenomena, two different ratios of PKC to PPF were studied (1:1 and 3:1,
w/
w). PPF is normally utilized for the prevention of the solids packing and porosity increase, serving mostly as a supporting matrix or a texturizer rather as a nutrient source [
51]. In the case of 1:1 ratio, the produced microbial oil did not exceed the initial oil content of the unfermented substrate. Specifically, it was observed a depletion of PKC oil by 62.9% and 23.9% for
C. echinulata and
M. ramanniana MUCL, respectively, after 11 days of fermentation. This was possibly related to the lignin-rich and protein-low content of PPF. More specifically, the protein content of PKC was found to be 12.8% while that of PPF was substantially lower (4.2%) (
Table 1). Rationally, the capacity of
C. echinulata to produce proteases (
Section 3.7) could lead to efficient enzymatic hydrolysis of rich in protein substrates, i.e., PKC, resulting in the release of amino acids and peptides, and thus favoring the fungal growth. Regarding lignin, PPF has been reported to possess a significant higher content (26.9%) compared to PKC (13.3%) [
35]. High lignin contents of lignocellulosic matrices have been related to the adsorption of enzymes onto the lignin structure, reducing the available amounts of the former for targeted action and thus limited nutrients uptake by the fungal strain for cell growth and metabolite synthesis [
52]. The moisture loss was 33.0% for
C. echinulata and 19.1% for
M. ramanniana MUCL.
C. echinulata showed similar behavior when cultivated at a PKC:PPF ration of 3:1. Considering
M. ramanniana MUCL, it produced 69.9 mg/gfs of lipids after 11 days of fermentation, when applying the 3:1 solid ratio with a moisture loss equal to 17.1%.
Fatty acid composition of lipids produced by
C. echinulata under the two different solid ratios was quite different compared to the initial fatty acid composition of the unfermented solids (
Table 5). C12:0 and C14:0 were decreased to 17.7–18.9% and 6.9%, respectively. C16:0 and C18:1 were increased, reaching 20.0–21.0% and 23.3–36.7%, respectively. When
C. echinulata was cultivated in the 1:1 ratio, the GLA content was slightly higher (3.2%) than in the 3:1 ratio (2.7%). The RSU was importantly decreased in both ratios. The ratio of C/N is the main factor of microbial growth and microbial oil production. Tailoring this ratio, the microbial metabolism can be shifted towards the biosynthesis of unsaturated fatty acids [
50]. Desirable C/N ratios can be obtained by mixing two solid substrates of different C and N concentrations, i.e., PKC (low C/N ratio = 25.2) and PPF (high C/N ratio = 69.5) (
Table 1). In the case of
M. ramanniana MUCL, the fatty acid profile of 1:1 ratio was similar to the initial unfermented substrate. At 3:1 ratio, a substantial increase of C18:1 (31.9%) and decrease of C12:0 (22.1%) was monitored. The GLA content of microbial oil was increased up to 1.9%.
3.6. Effect of Nutrient Supplementation
SSF with C. echinulata and M. ramanniana MUCL (under the best performing conditions of temperature and initial moisture content) were repeated in an air humidified chamber with 90% air saturation. Surprisingly, the lipids yields did not show considerable alterations compared to the results of SSF that was conducted in a non-humidified chamber. These results could be partially related to the fact that when C. echinulata was cultivated under the best conditions on non-humidified chambers, the moisture losses were quite low (~20%) and quite comparable with the values obtained when humidified chambers were used (~21.5%).
Consequently, experiments with
C. echinulata demonstrated the highest potential for lipid accumulation, and therefore, this fungal strain was selected for further evaluation. In this case, an attempt to monitor the biomass production was conducted via the estimation of fungal glucosamine content. Four sets of fermentations were carried out, with individual supplementation of PKC with 5% molasses, 5% crude glycerol, and a mixture of mineral solution or nitrogen sources. Except for nitrogen sources, the other supplements resulted in high moisture losses, up to 51.0% after 8 days of fermentation and low biomass production, varying between 54.0 and 83.5 mg/gfs. The produced intracellular lipids did not exceed the initial oil of the unfermented PKC. The initial oil content of the unfermented solids (
Table 1) was depleted by 64.4–80.8% after 8 days of fermentation. Enrichment of PKC with nitrogen sources seemed to trigger lipid production. Maximum lipid production of 91.0 ± 3.9 mg/gfs was achieved at 7 days in the presence of nitrogen source, indicating that PKC enrichment with additional nitrogen sources could enhance lipid productivity in SSF. Moreover, the high biomass formation in this case (144.3 mg/gfs) seemed to deteriorate moisture loss of the substrate to 8.6% after 8 days of fermentation, which is very favorable. In the study by Conti et al. [
20], mixtures of barley and spent malt grains were supplemented with yeast extract, peptone, Mg
2+, phosphates, and trace minerals to enhance lipids production by
C. elegans CCF. However, lipids production was not favored, indicating that the aforementioned compounds were present in the cereals, at adequate levels.
Fatty acid profile of oil that derived from all the different supplementation treatments showed alterations in comparison to the oil of the unfermented PKC (
Table 6). C12:0 was reduced in all cases varying between 24.8 and 39.0%, while C18:1 and C16:0 were increased in the range of 23.4–39.1% and 10.3–18.1%, respectively. C14:0 remained either almost stable (14.8%) as in the case of nitrogen sources enrichment, or it was decreased (7.7–10.4%). Moreover, GLA was not detected in any of the applied treatment.
3.7. Screening of Protein-Rich Agricultural Residues for Lipid Production
Based on the results of
Section 3.6, it was indicated that
C. echinulata may perform better in solid matrices with higher protein content than PKC. Thus, two agricultural residues, namely, CoSC and CaSC, which are very rich in protein (
Table 1), were selected for subsequent solid-state experiments.
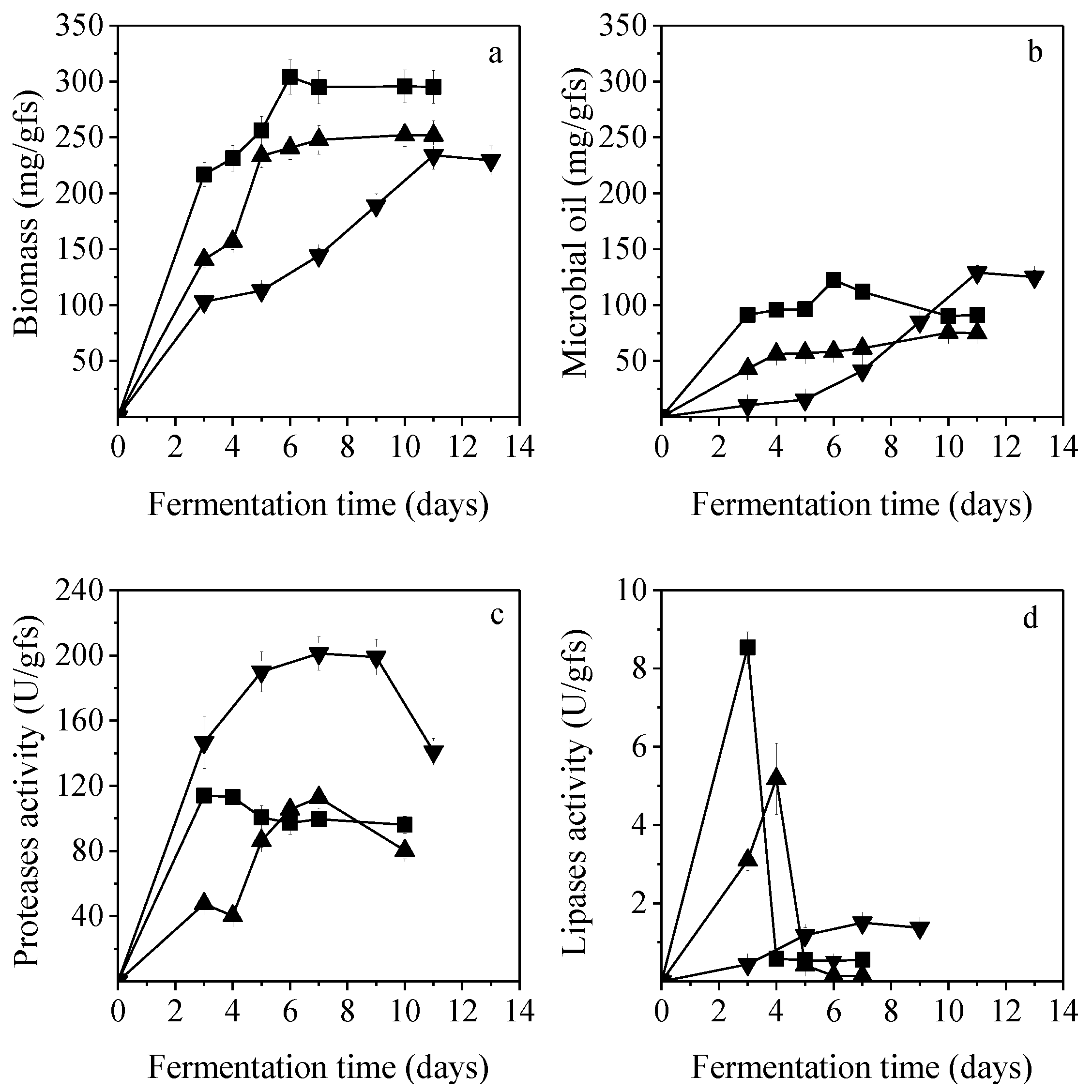
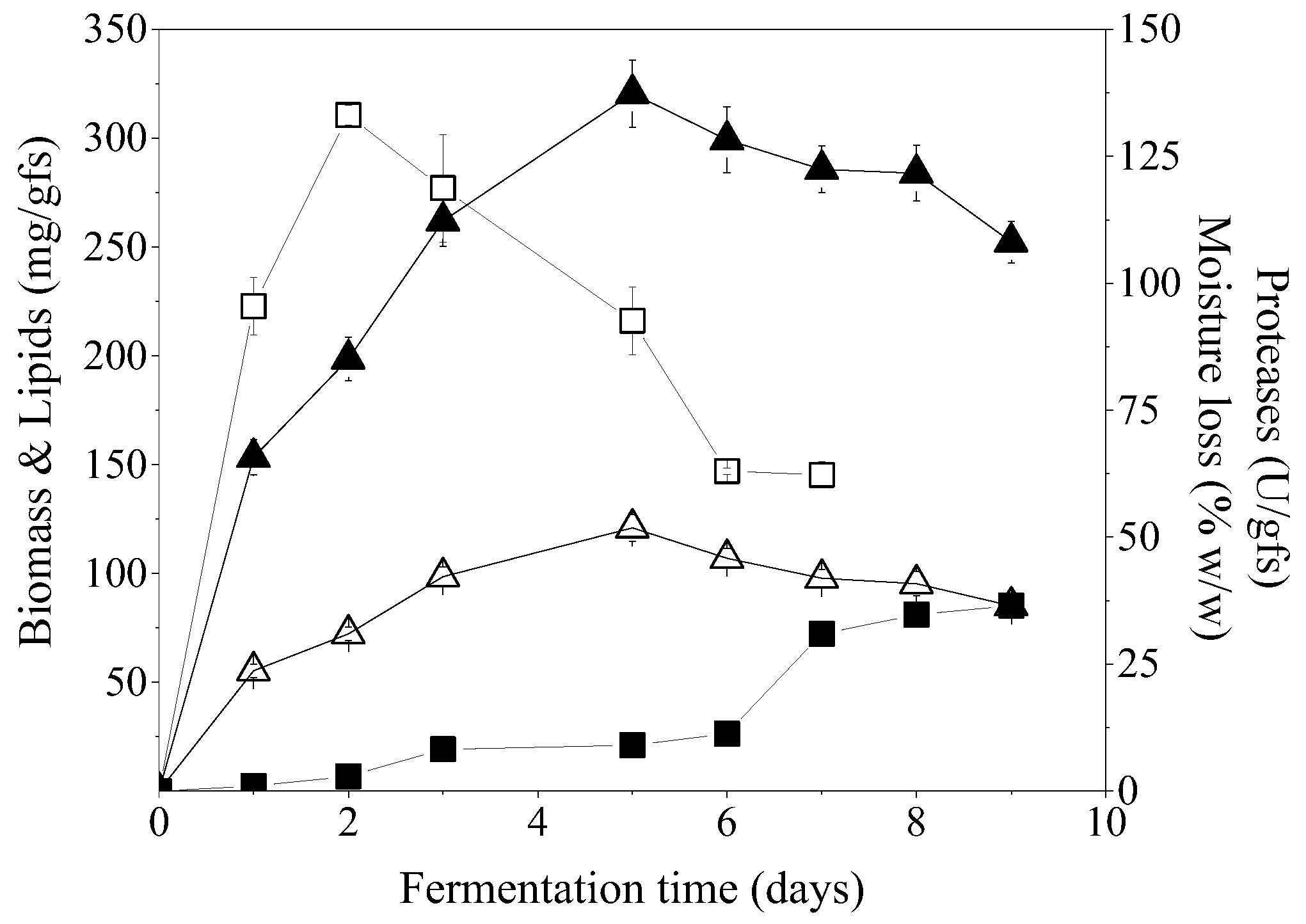
Figure 4 presents biomass growth, lipids accumulation, proteases activity, and lipases activity during SSF of PKC, CoSC, and CaSC.
C. echinulata was efficiently grown in all the renewable resources, with CoSC being the most efficient for biomass (304.1 mg/gfs) and lipid production (122.2 mg/gfs) with an intracellular lipid content of 40.2% at 6 days of SSF. Biomass production obtained on CoSC at 6-days of SSF showed significant differences (
p < 0.05) compared to those obtained using PKC and CaSC at 11 days and 10 days of SSF, respectively. The use of PKC and CaSC as SSF substrates did not reveal any statistical difference regarding biomass production.
The concertation of lipids production was not affected when SSF was implemented on PKC or CoSC. Thus, the productivity of the produced microbial oil in the case of PKC was two folds lower in comparison to CoSC while biomass production amounted to 234.1 mg/gfs, after 11 days of fermentation. SSF of CaSC led to a fungal biomass of 252.1 mg/gfs with low intracellular lipid content (29.9%) which was attributed to the low C/N ratio of the substrate (
Table 1). In this case, lipids concentration showed a statistically significant difference (
p < 0.05) compared to PKC and CoSC. Higher C/N ratios favor intracellular lipids production while lower ratios up to a particular threshold have been proven to be beneficial for biomass growth [
33]. As it is depicted in
Table 1, CaSC showed the lowest C/N ratio (8.0), followed by CoSC (11.5) and PKC (25.2). Moreover, the presence of a potent toxin (ricin) and an allergenic protein fraction (CB-1A or 2S albumin isoforms) may also have influenced the metabolism of lipogenesis since they are limiting factors for the assimilation of castor seed waste by living organisms [
53].
Proteases production was quite enhanced in the three solid matrices. In the case of CaSC and CoSC, maximum respective values of 112.8 U/gfs after 7 days of fermentation and 114.2 U/gfs after 3 days of fermentation were achieved. Proteolytic activity in PKC was 146.7 U/gfs after 3 days and reached the maximum value of 201.2 U/gfs after 7 days of fermentation. Lipases activity in all substrates was quite low. The highest value was observed on CoSC at the 3rd day (8.5 U/gfs) while it sharply decreased thereafter (0.5 U/gfs). Considering CaSC, lipases activity reached the maximum value of 5.2 U/gfs at the 4th day of fermentation while it decreased to 0.1–0.4 U/gfs thereafter, whereas in the case of PKC, it was in the range of 0.4–1.5 U/gfs throughout the fermentation. Vegetable oils, including castor, soybean, olive, corn, and palm oils, have been investigated as potential inducers of lipase production, with castor oil being the most efficient for maximal enzymatic activity [
18]. Rationally, castor oil and palm kernel oil contained in CaSC and PKC would lead to enhanced lipases production in the respective fermentations. Thus, the low lipases production that were determined in this study are attributed to the fact that lipases activity has been reported to be negatively affected by proteolysis [
33]. This has been validated via an unstructured mathematical model that describes time course variations of extracellular lipases and proteases activities for batch fermentations of the fungus
Penicillium restrictum [
54]. Due to the aforementioned, high proteases released in the solid medium in this particular study appear to be in detriment of lipases production, leading to their inactivation.
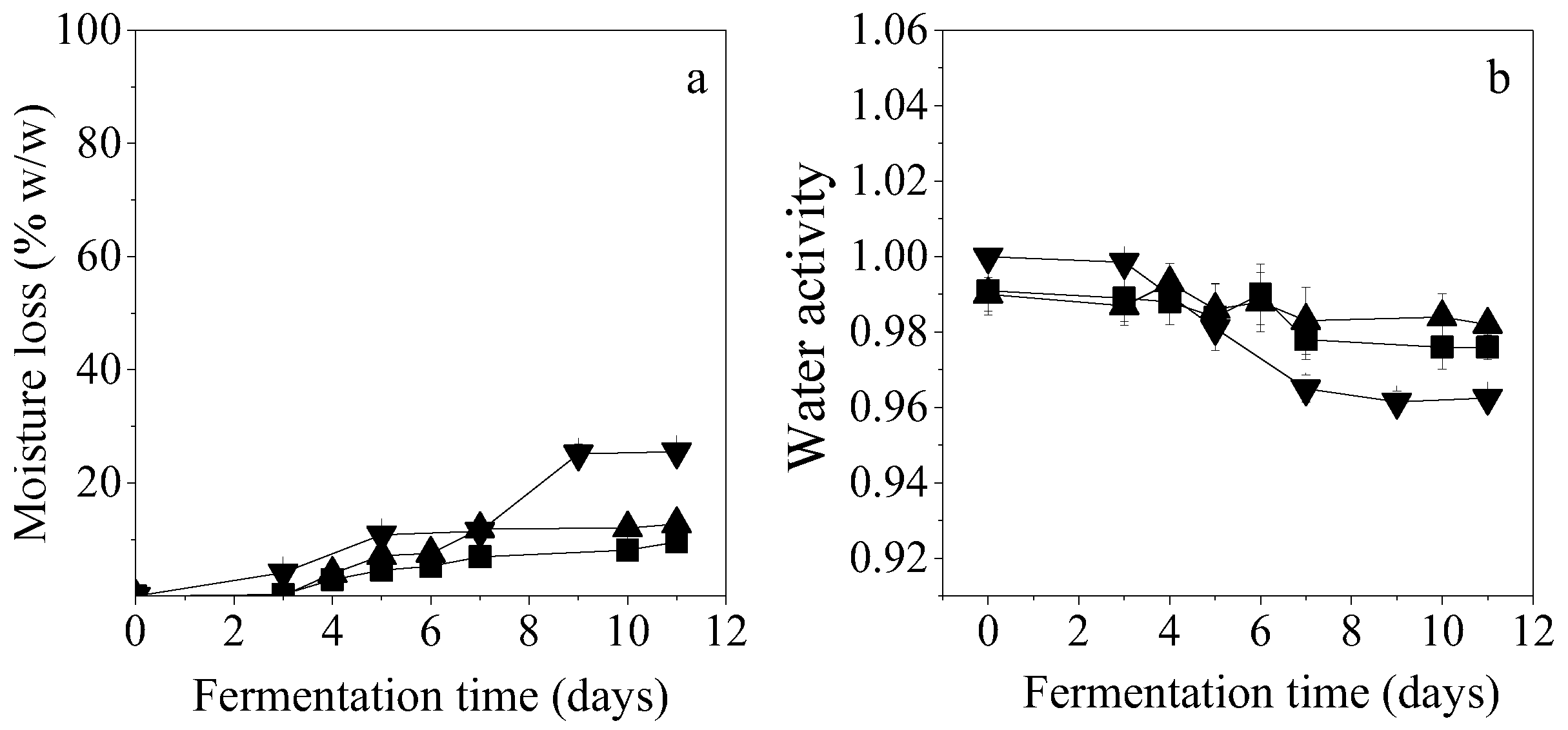
Moisture losses for CoSC and CaSC reached 9.6% and 12.8% (after 11 days of fermentation), respectively, while PKC resulted in 25.5% loss at the same fermentation time (
Figure 5a). Aw constitutes a thermodynamic factor that indicates the available water of the solid substrate, participating in several reactions that take place during the SSF procedure. Aw decreased during SSF as a result of moisture losses of the solid substrates. Decrease of initial moisture of PKC was higher than that of the other substrates, therefore leading to higher Aw changes and finally reduced to 0.96. For CoSC and CaSC, Aw was reduced up to 0.98 at the end of fermentation (
Figure 5b).
Table 7 shows the fatty acid profile of microbial lipids produced during the cultivation of
C. echinulata on CoSC and CaSC. The initial oil of CoSC and lipids extracted from the fermented CoSC presented similar profile, mainly consisting of C18:2 (49.7–51.9%) and C16:0 (27.1–27.5%) followed by C18:1 (16.6–17.1%). Lipids extracted from fermented CaSC showed alterations compared to the initial substrate’s oil. Ricinoleic acid ((12R) C18:1) was the predominant fatty acid, and it was decreased to 65.2%. C16:0 and C18:1 were increased to 9.5% and 10.3%, respectively. GLA content of the fermented CoSC and CaSC was quite low (0.8%).
3.8. SSF in PBR
Lipid production was scaled up from tray bioreactor to lab-scale PBR. Maximum biomass growth (320.5 mg/gfs) was observed after 5 days of fermentation with a lipid content of 37.7%, and a productivity of 0.024 g/gfs/d (
Figure 6). Although biomass and lipids production were lower than those achieved in tray bioreactors, the productivity was higher. Lipases activity was low (0.3–1.4 U/gfs) while maximum proteases activity was achieved after 2 days of fermentation (133.1 U/gfs) which then gradually decreased to 62.2 U/gfs at the 7th day of fermentation. Moisture losses were quite controllable until the 6th day of fermentation reaching up to 11.5% while they increased thereafter (30.9–36.5%). This is probably attributed to porosity reduction of the bed that was observed during the fermentation, creating a packed cotton matrix [
26]. The formation of the packed solid column can result in limited aeration of the fermentation and is consequently allied to deteriorations in its efficiency. Fatty acid analysis showed that C18:2 (45.8%) was the predominant fatty acid, followed by C16:0 (33.9%), C18:1 (16.4%), and C18:0 (3.9%).
Studies that deal with lipids production via SSF are rather limited. Several attempts have been reported in shake flasks, Petri dishes or plastic bags. Stredansky et al. [
19] demonstrated the efficient lipid production varying between 150 and 168 mg/gfs with productivities of 0.019–0.026 g/gfs/d, when
Cunninghamella and
Mortierella strains were cultivated on apple pomace and spent malt grain (SMG) impregnated with a nutrient solution. GLA content fluctuated between 4.2 and 7.4%. In the same study,
T. elegans resulted in 212 mg/gfs of lipids with a GLA content of 9.1% when it was cultivated on a mixture of apple pomace and SMG impregnated with peanut oil and a nutrient solution under forced aeration [
19]. Conti et al. [
20] screened several fungal strains of the order
Mucorales for lipid production.
C. elegans was the most efficient strain resulting in 172 mg/gfs of lipids, productivity of 0.024 g/gfs/d, and 11.6% GLA when cultivated in a 15:5:1 barley–SMG–peanut oil and nutrient solution. In the study by Dulf et al. [
22],
Actinomucor elegans and
Umbelopsis isabellina were evaluated for their ability to produce lipids on grape pomace.
U. isabelline yielded the highest amount of microbial oil (160 mg/gfs) with a GLA content of 3.8 mg/gfs while
A. elegans produced around 120 mg/gfs of microbial oil with a lower GLA content (1.9 mg/gfs). Lower lipids production has been reported by Peng and Chen [
55] (23–42 mg/gfs), Peng and Chen [
56] (79.0 mg/gfs), Hui et al. [
57] (62.9 mg/gfs), and Zhang and Hu [
21] (47.9 mg/gfs) with productivities ranging between 0.002 and 0.01 mg/gfs/d using wheat straw and bran and soybean hulls as substrates. This indicates that the composition of substrate plays a key role in lipid production and fatty acid composition.
To the best of our knowledge, this is the first study that reports microbial oil production in PBR. Overall, the microbial oil production that was achieved (121.0 mg/gfs) and the productivity (0.024 g/gfs/d) were quite comparable with most of the aforementioned studies or even higher. Experiments conducted on tray reactors led to the production of microbial oil varying between 75.4 and 129.1 mg/gfs when PKC, CoSC, and CaBC were employed as the fermentative substrate while GLA was in the range of 0.8–4.5% and productivities of 0.008–0.02 mg/gfs/d. Further investigation on SSF supplementation, pretreatment, and aeration of substrate need to be investigated in order to achieve microbial oil concentrations and GLA content efficient for industrial implementation.
3.9. Polyol Esters Production and Characterization
Microbial lipids with fatty acid composition similar to vegetable oils are suitable for the production of oleochemicals [
43], eliminating thus the utilization of food-grade vegetable oils for this purpose. The microbial oil derived from
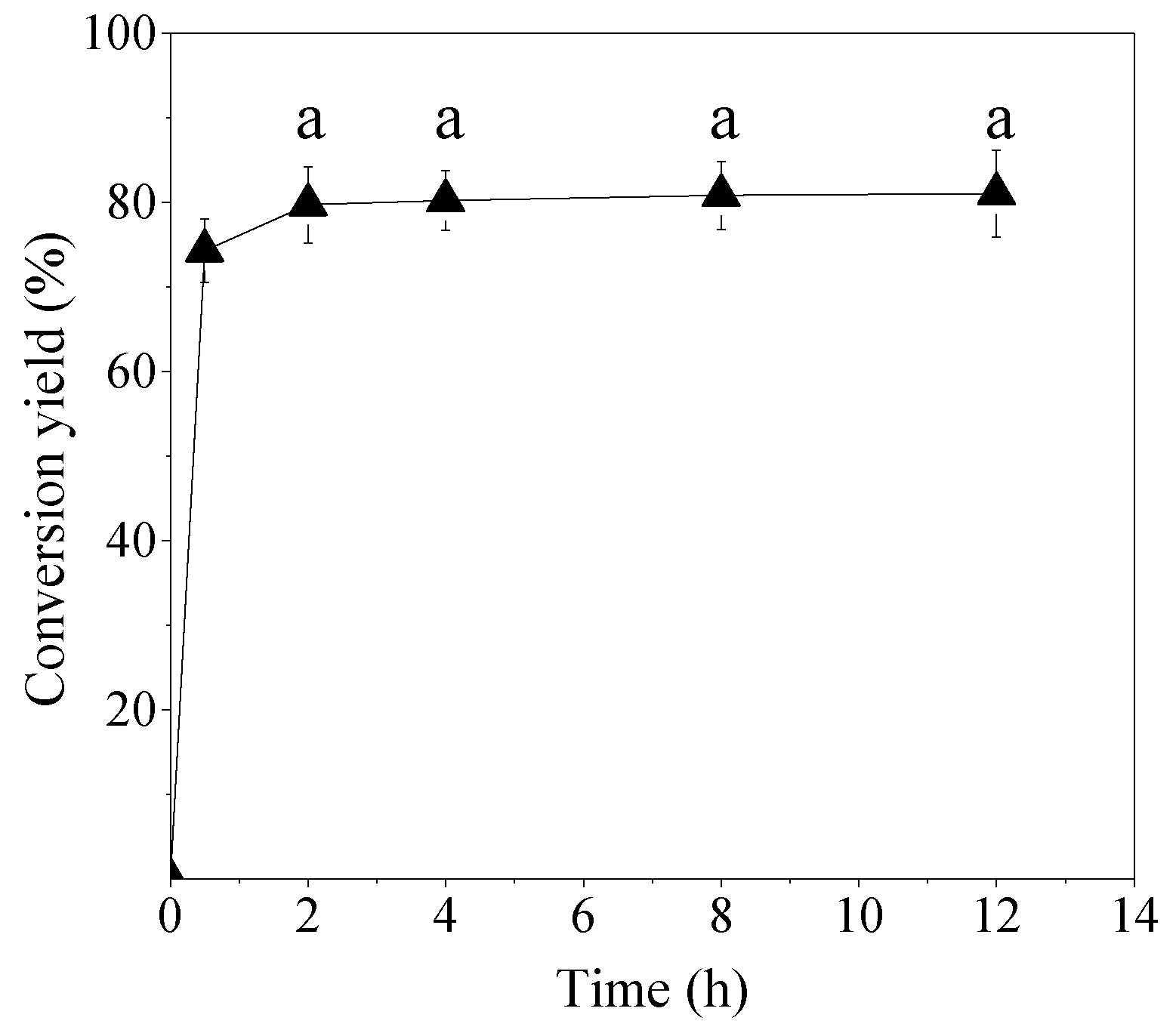
C. echinulata, which was extracted from fermented CoSC, was initially hydrolyzed to obtain a free fatty acid-rich fraction. The enzymatic bioconversion of free fatty acids to polyol esters was monitored via the reduction of the acidity, since analytical standards of polyols esters are not commercially available. Initial acidity of microbial oil was 18.0% while after 24 h of enzymatic hydrolysis, it was increased to 87.7%. Conversion yield reached up to 74.3% after 30 min of reaction while a conversion yield higher than 80% was achieved after 2 h, remaining almost stable thereafter (
Figure 7).
Vegetable oils have been reported to have great potential to be applied as lubricants, while biocatalysts, i.e., lipases are very efficient to bioconvert vegetable oils into fatty acid methyl esters with high conversion rates (up to 99%) [
58]. The main disadvantage of vegetable oils is the fact that their availability highly depends on the climatic conditions; they compete with food, and they require agricultural land [
59]. Thus, alternative oil sources are of outmost importance. Research dealing with microbial oil as the initiating material for biolubricants production is limited in the cited publication. Papadaki et al. [
27] reported the successful utilization of microbial oil produced from oleaginous yeasts to produce biolubricants with a conversion yield of 88% (NPG esters at 72 h). In another study, microbial oil derived from
Rhodotorula glutinis was enzymatically converted to trimethylolpropane fatty acid triesters with a yield of 89.5%. The produced biolubricants showed better tribological properties compared to fossil-based lubricants of the same viscosity [
60].
The composition of NPG esters concerning the content of di- and mono-esters was determined via NMR analysis (
Table 8). According to the data obtained, monoester content (29.6%,
w/
w) was quite lower than the diesters content (56.1%,
w/
w). The non-esterified content of NPG was 3.5% while fatty acids content was 10.8%. This could be improved by using surplus free fatty acid content during esterification. For instance, Cavalcanti et al. [
61] reported that enzymatic esterification of free fatty acids of soybean oil with NPG resulted in 100% esterification of the OH groups when applying a molar ratio of free fatty acids to polyol equal to 3.75:1.
Desirable viscosity values and indices are related to high working efficiencies of mechanical devices and low energy requirements, as a result of favorable friction forces that lead to prevention of collision and rubbing between two contacting surfaces. High viscosity indices ensure that lubricants remain effective even at high temperatures by maintaining the thickness of the oil film. The polyol esters were characterized in terms of acid number (23.2 mg KOH/g), kinematic viscosity (30.1 mm
2/s at 40 °C and 6.2 mm
2/s at 100 °C) and viscosity index (161). The viscosity index is quite comparable with esters produced via enzymatic esterification of NPG with free fatty acids derived from yeast lipids (181–183) or soybean oil (214) [
61]. Since microbial oil consists of triglycerides that maintain intermolecular interactions in elevated temperatures, the viscosity index of microbial oil-based lubricants is normally higher than that of mineral oils [
62].


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}



