
Microbial Melanin: Renewable Feedstock and Emerging Applications in Food-Related Systems


Abstract
:1. Introduction
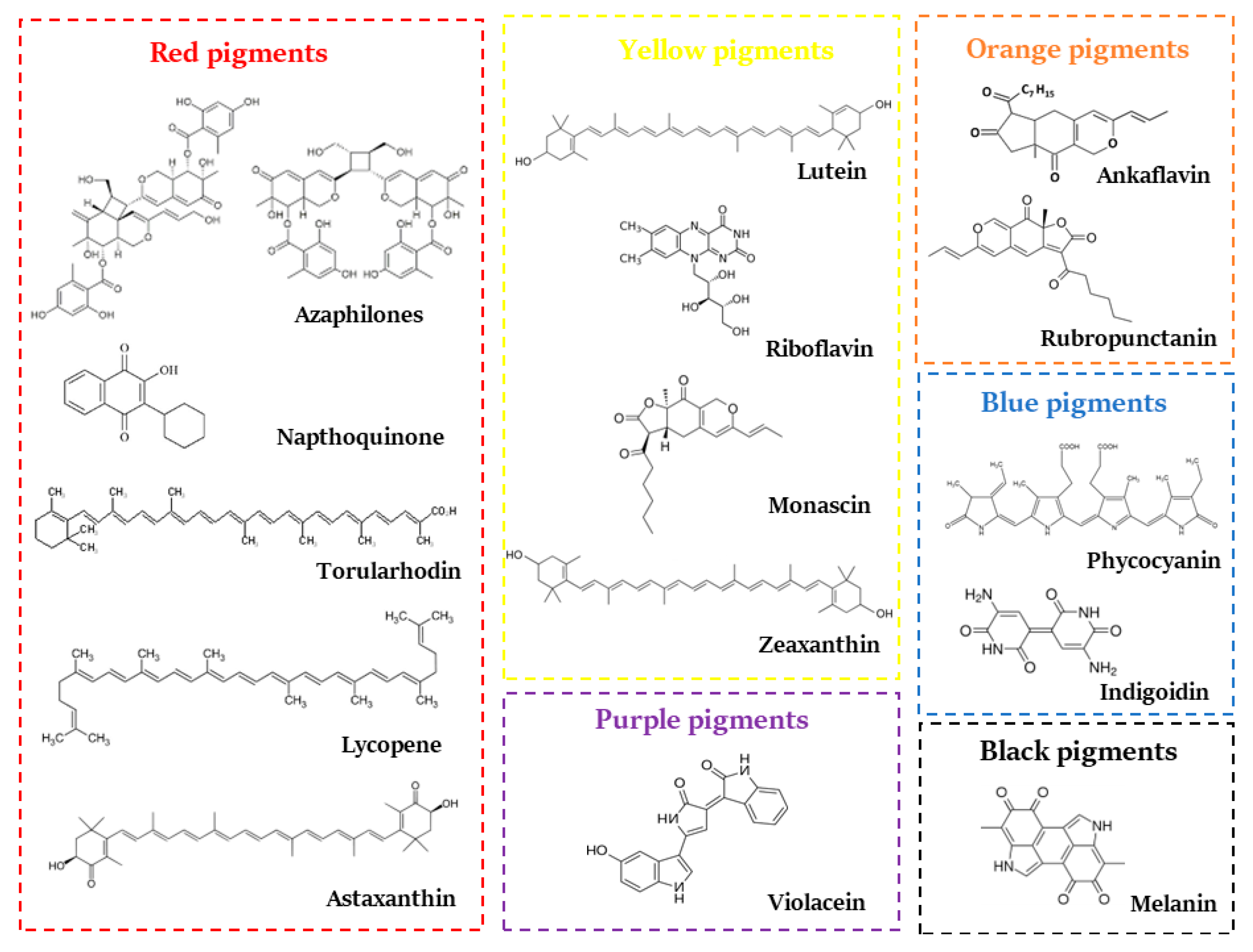
2. Microbial Pigments
3. Melanin Production via Fermentation
3.1. Melanin Production Using Commercial-Based Fermentation Media
3.2. Low-Cost and Renewable Agri-Food Waste and Byproduct Streams
3.3. Repurposing Agri-Food Waste via Melanin Production
3.4. Genetic Tools for Microbial Melanin Production
4. Melanin Polymers
4.1. Properties and Classification of Melanin
4.2. Structural Diversity of Melanin

4.3. Biosynthesis of Melanin from Microbes
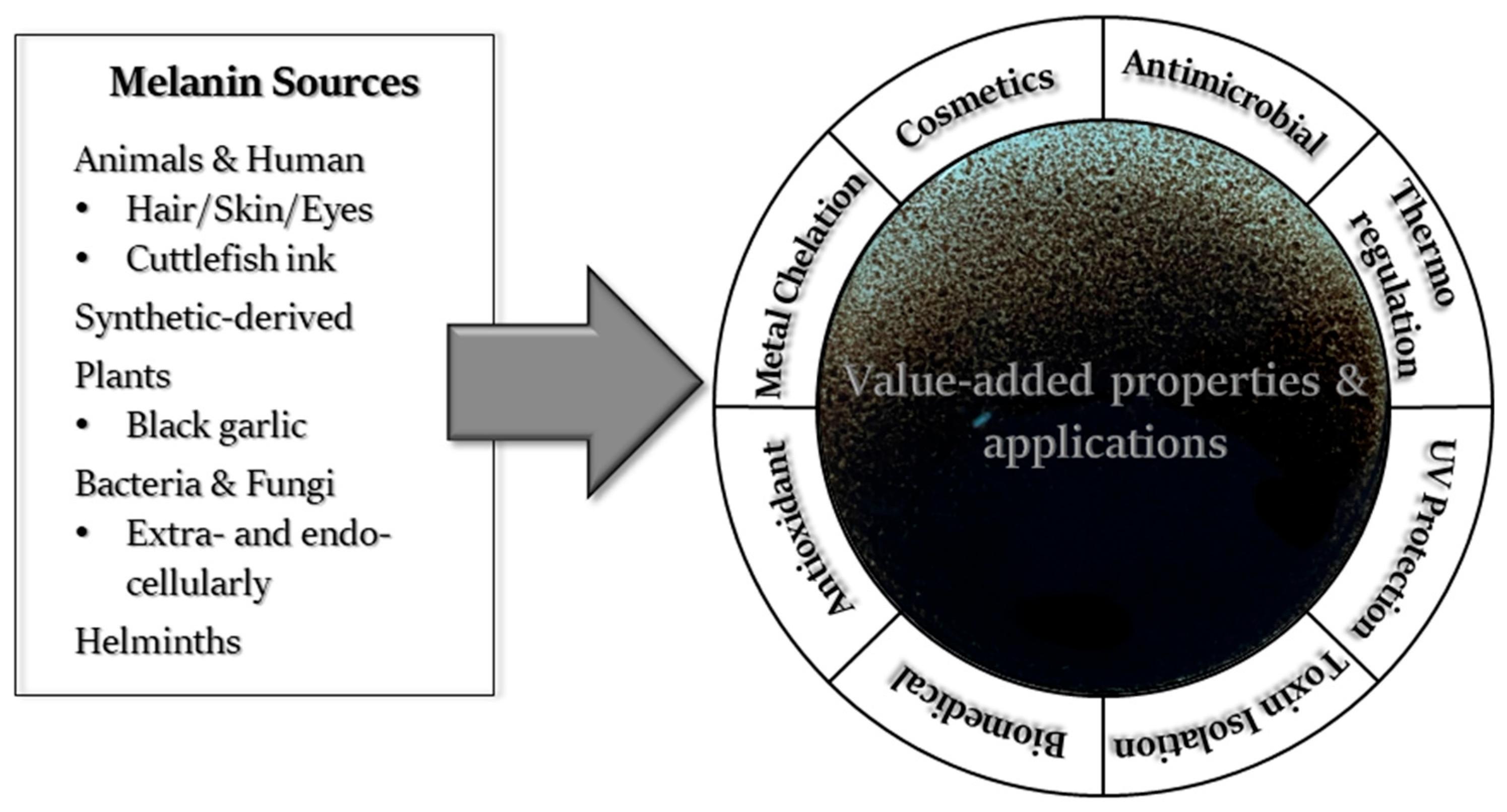
5. Property-Dependent Applications of Melanin
5.1. Absorption and Binding Capacities of Melanin and Related Properties
5.2. Antimicrobial Activity
5.3. Antioxidant Activity
5.4. The Case of Food Packaging Formulation
5.4.1. Biopolymers for Food Packaging Materials
5.4.2. Melanin Enhanced Biofilms
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tran-Ly, A.N.; Ribera, J.; Schwarze, F.W.M.R.; Brunelli, M.; Fortunato, G. Fungal Melanin-Based Electrospun Membranes for Heavy Metal Detoxification of Water. Sustain. Mater. Technol. 2020, 23, e00146. [Google Scholar] [CrossRef]
- Tran-Ly, A.N.; Reyes, C.; Schwarze, F.W.M.R.; Ribera, J. Microbial Production of Melanin and Its Various Applications. World J. Microbiol. Biotechnol. 2020, 36, 170. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.-A.; Saber, W.I.A. Natural Melanin: Current Trends, and Future Approaches, with Especial Reference to Microbial Source. Polymers 2022, 14, 1339. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Rhim, J.-W. New Insight into Melanin for Food Packaging and Biotechnology Applications. Crit. Rev. Food Sci. Nutr. 2022, 62, 4629–4655. [Google Scholar] [CrossRef]
- Ribera, J.; Panzarasa, G.; Stobbe, A.; Osypova, A.; Rupper, P.; Klose, D.; Schwarze, F.W.M.R. Scalable Biosynthesis of Melanin by the Basidiomycete Armillaria Cepistipes. J. Agric. Food Chem. 2019, 67, 132–139. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; El-Ewasy, S.M. Bioproduction, Characterization, Anticancer and Antioxidant Activities of Extracellular Melanin Pigment Produced by Newly Isolated Microbial Cell Factories Streptomyces Glaucescens NEAE-H. Sci. Rep. 2017, 7, 42129. [Google Scholar] [CrossRef]
- Pavan, M.E.; López, N.I.; Pettinari, M.J. Melanin Biosynthesis in Bacteria, Regulation and Production Perspectives. Appl. Microbiol. Biotechnol. 2020, 104, 1357–1370. [Google Scholar] [CrossRef]
- Cao, W.; Zhou, X.; McCallum, N.C.; Hu, Z.; Ni, Q.Z.; Kapoor, U.; Heil, C.M.; Cay, K.S.; Zand, T.; Mantanona, A.J.; et al. Unraveling the Structure and Function of Melanin through Synthesis. J. Am. Chem. Soc. 2021, 143, 2622–2637. [Google Scholar] [CrossRef]
- Singla, S.; Htut, K.Z.; Zhu, R.; Davis, A.; Ma, J.; Ni, Q.Z.; Burkart, M.D.; Maurer, C.; Miyoshi, T.; Dhinojwala, A. Isolation and Characterization of Allomelanin from Pathogenic Black Knot Fungus—A Sustainable Source of Melanin. ACS Omega 2021, 6, 35514–35522. [Google Scholar] [CrossRef]
- Market Watch 2023. Melanin Market Size 2023 Trend and Opportunities. Available online: https://www.marketwatch.com/press-release/melanin-cas-8049-97-6-market-size-2023-trend-and-opportunities-analysis-cagr-and-value-chain-study-business-growth-to-2028-2023-02-02 (accessed on 15 January 2023).
- Panesar, R.; Kaur, S.; Panesar, P.S. Production of Microbial Pigments Utilizing Agro-Industrial Waste: A Review. Curr. Opin. Food Sci. 2015, 1, 70–76. [Google Scholar] [CrossRef]
- Lopes, F.C.; Ligabue-Braun, R. Agro-Industrial Residues: Eco-Friendly and Inexpensive Substrates for Microbial Pigments Production. Front. Sustain. Food Syst. 2021, 5, 589414. [Google Scholar] [CrossRef]
- Mavria, A.; Tsouko, E.; Protonotariou, S.; Papagiannopoulos, A.; Georgiadou, M.; Selianitis, D.; Pispas, S.; Mandala, I.; Koutinas, A.A. Sustainable Production of Novel Oleogels Valorizing Microbial Oil Rich in Carotenoids Derived from Spent Coffee Grounds. J. Agric. Food Chem. 2022, 70, 10807–10817. [Google Scholar] [CrossRef]
- Tsouko, E.; Papadaki, A.; Papapostolou, H.; Ladakis, D.; Natsia, A.; Koutinas, A.; Kampioti, A.; Eriotou, E.; Kopsahelis, N. Valorization of Zante Currant Side-streams for the Production of Phenolic-rich Extract and Bacterial Cellulose: A Novel Biorefinery Concept. J. Chem. Technol. Biotechnol. 2020, 95, 427–438. [Google Scholar] [CrossRef]
- Tsouko, E.; Maina, S.; Alexandri, M.; Papapostolou, H.; Koutinas, A. Recovery of Value-Added Products and Biological Conversion of Coffee and Citrus Processing Waste Using Green Technologies. In Green Chemistry in Agriculture and Food Production; CRC Press: Boca Raton, FL, USA, 2023; pp. 111–149. [Google Scholar] [CrossRef]
- Alexandri, M.; Maina, S.; Tsouko, E.; Papapostolou, H.; Koutinas, A.; Kourmentza, K. Valorization of Fruit Processing By-Product Streams into Integrated Biorefinery Concepts: Extraction of Value-Added Compounds and Bioconversion to Chemicals. In Valorization of Agri-Food Wastes and By-Products; Elsevier: Amsterdam, The Netherlands, 2021; pp. 927–945. [Google Scholar] [CrossRef]
- Tsouko, E.; Papadaki, A.; Papanikolaou, S.; Danezis, G.P.; Georgiou, C.A.; Freire, D.M.G.; Koutinas, A. Enzymatic Production of Isopropyl and 2-Ethylhexyl Esters Using γ-Linolenic Acid Rich Fungal Oil Produced from Spent Sulphite Liquor. Biochem. Eng. J. 2021, 169, 107956. [Google Scholar] [CrossRef]
- Tsouko, E.; Papanikolaou, S.; Koutinas, A.A. Production of Fuels from Microbial Oil Using Oleaginous Microorganisms. In Handbook of Biofuels Production; Elsevier: Amsterdam, The Netherlands, 2016; pp. 201–236. [Google Scholar] [CrossRef]
- Pilafidis, S.; Diamantopoulou, P.; Gkatzionis, K.; Sarris, D. Valorization of Agro-Industrial Wastes and Residues through the Production of Bioactive Compounds by Macrofungi in Liquid State Cultures: Growing Circular Economy. Appl. Sci. 2022, 12, 11426. [Google Scholar] [CrossRef]
- Palacio-Barrera, A.M.; Areiza, D.; Zapata, P.; Atehortúa, L.; Correa, C.; Peñuela-Vásquez, M. Induction of Pigment Production through Media Composition, Abiotic and Biotic Factors in Two Filamentous Fungi. Biotechnol. Rep. 2019, 21, e00308. [Google Scholar] [CrossRef]
- Sánchez-Muñoz, S.; Mariano-Silva, G.; Leite, M.O.; Mura, F.B.; Verma, M.L.; da Silva, S.S.; Chandel, A.K. Production of Fungal and Bacterial Pigments and Their Applications. In Biotechnological Production of Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2020; pp. 327–361. [Google Scholar] [CrossRef]
- Venil, C.K.; Devi, P.R.; Ahmad, W.A. Agro-Industrial Waste as Substrates for the Production of Bacterial Pigment. In Valorisation of Agro-industrial Residues— Volume I: Biological Approaches; Applied Environmental Science and Engineering for a Sustainable Future; Zakaria, Z.A., Boopathy, R., Dib, J.R., Eds.; Springer International Publishing: Cham, Switzerland; Berlin, Germany, 2020; pp. 149–162. [Google Scholar] [CrossRef]
- Tuli, H.S.; Chaudhary, P.; Beniwal, V.; Sharma, A.K. Microbial Pigments as Natural Color Sources: Current Trends and Future Perspectives. J. Food Sci. Technol. 2015, 52, 4669–4678. [Google Scholar] [CrossRef]
- Dyaa, A.; Soliman, H.; Abdelrazak, A.; Samra, B.N.; Khojah, E.; Ahmed, A.F.; El-Esawi, M.A.; Elsayed, A. Optimization of Carotenoids Production from Rhodotorula Sp. Strain ATL72 for Enhancing Its Biotechnological Applications. J. Fungi 2022, 8, 160. [Google Scholar] [CrossRef]
- Chen, W.; Feng, Y.; Molnár, I.; Chen, F. Nature and Nurture: Confluence of Pathway Determinism with Metabolic and Chemical Serendipity Diversifies Monascus Azaphilone Pigments. Nat. Prod. Rep. 2019, 36, 561–572. [Google Scholar] [CrossRef]
- Romsdahl, J.; Schultzhaus, Z.; Cuomo, C.A.; Dong, H.; Abeyratne-Perera, H.; Hervey, W.J.; Wang, Z. Phenotypic Characterization and Comparative Genomics of the Melanin-Producing Yeast Exophiala Lecanii-Corni Reveals a Distinct Stress Tolerance Profile and Reduced Ribosomal Genetic Content. J. Fungi 2021, 7, 1078. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Ruiz-Sánchez, J.P.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Méndez-Zavala, A.; Giuffrida, D.; Dufossé, L.; Montañez, J. Biotechnological Approaches for the Production of Natural Colorants by Talaromyces/Penicillium: A Review. Biotechnol. Adv. 2020, 43, 107601. [Google Scholar] [CrossRef] [PubMed]
- Selianitis, D.; Efthymiou, M.-N.; Tsouko, E.; Papagiannopoulos, A.; Koutinas, A.; Pispas, S. Nanocellulose Production from Different Sources and Their Self-Assembly in Composite Materials. In Handbook of Nanocelluloses; Barhoum, A., Ed.; Springer International Publishing: Cham, Switzerland; Berlin, Germany, 2021; pp. 1–32. [Google Scholar] [CrossRef]
- Lopes, H.J.S.; Bonturi, N.; Miranda, E.A. Rhodotorula Toruloides Single Cell Oil Production Using Eucalyptus Urograndis Hemicellulose Hydrolysate as a Carbon Source. Energies 2020, 13, 795. [Google Scholar] [CrossRef]
- Martínez, L.M.; Martinez, A.; Gosset, G. Production of Melanins With Recombinant Microorganisms. Front. Bioeng. Biotechnol. 2019, 7, 285. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Nimse, S.B.; Mathew, D.E.; Dhimmar, A.; Sahastrabudhe, H.; Gajjar, A.; Ghadge, V.A.; Kumar, P.; Shinde, P.B. Microbial Melanin: Recent Advances in Biosynthesis, Extraction, Characterization, and Applications. Biotechnol. Adv. 2021, 53, 107773. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.J.B.; Casadevall, A. Functions of Fungal Melanin beyond Virulence. Fungal Biol. Rev. 2017, 31, 99–112. [Google Scholar] [CrossRef]
- Cordero, R.J.B.; Vij, R.; Casadevall, A. Microbial Melanins for Radioprotection and Bioremediation. Microb. Biotechnol. 2017, 10, 1186–1190. [Google Scholar] [CrossRef]
- Sindhu, R.; Binod, P.; Nair, R.B.; Varjani, S.; Pandey, A.; Gnansounou, E. Waste to Wealth. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2020; pp. 181–197. [Google Scholar] [CrossRef]
- Müjdeci, G.N. Experimental Modeling and Optimization of Melanin Production by Aureobasidium Pullulans NBRC 100716 in Carrot Peel Extract. Environ. Prog. Sustain. Energy 2022, 41, e13919. [Google Scholar] [CrossRef]
- Zou, Y.; Tian, M. Fermentative Production of Melanin by Auricularia Auricula. J. Food Process. Preserv. 2017, 41, e12909. [Google Scholar] [CrossRef]
- Tarangini, K.; Mishra, S. Production of Melanin by Soil Microbial Isolate on Fruit Waste Extract: Two Step Optimization of Key Parameters. Biotechnol. Rep. 2014, 4, 139–146. [Google Scholar] [CrossRef]
- Restaino, O.F.; Scognamiglio, M.; Mirpoor, S.F.; Cammarota, M.; Ventriglia, R.; Giosafatto, C.V.L.; Fiorentino, A.; Porta, R.; Schiraldi, C. Enhanced Streptomyces Roseochromogenes Melanin Production by Using the Marine Renewable Source Posidonia Oceanica Egagropili. Appl. Microbiol. Biotechnol. 2022, 106, 7265–7283. [Google Scholar] [CrossRef]
- Arikan, E.B.; Canli, O.; Caro, Y.; Dufossé, L.; Dizge, N. Production of Bio-Based Pigments from Food Processing Industry By-Products (Apple, Pomegranate, Black Carrot, Red Beet Pulps) Using Aspergillus Carbonarius. J. Fungi 2020, 6, 240. [Google Scholar] [CrossRef]
- Raman, N.M.; Shah, P.H.; Mohan, M.; Ramasamy, S. Improved Production of Melanin from Aspergillus Fumigatus AFGRD105 by Optimization of Media Factors. AMB Express 2015, 5, 72. [Google Scholar] [CrossRef]
- Oh, J.-J.; Kim, J.Y.; Kwon, S.L.; Hwang, D.-H.; Choi, Y.-E.; Kim, G.-H. Production and Characterization of Melanin Pigments Derived from Amorphotheca Resinae. J. Microbiol. 2020, 58, 648–656. [Google Scholar] [CrossRef]
- Elsayis, A.; Hassan, S.W.M.; Ghanem, K.M.; Khairy, H. Optimization of Melanin Pigment Production from the Halotolerant Black Yeast Hortaea Werneckii AS1 Isolated from Solar Salter in Alexandria. BMC Microbiol. 2022, 22, 92. [Google Scholar] [CrossRef]
- Ruan, L.; He, W.; He, J.; Sun, M.; Yu, Z. Cloning and Expression of Mel Gene from Bacillus Thuringiensis in Escherichia Coli. Antonie Van Leeuwenhoek 2005, 87, 283–288. [Google Scholar] [CrossRef]
- Guo, J.; Rao, Z.; Yang, T.; Man, Z.; Xu, M.; Zhang, X.; Yang, S.-T. Cloning and Identification of a Novel Tyrosinase and Its Overexpression in Streptomyces kathirae SC-1 for Enhancing Melanin Production. FEMS Microbiol. Lett. 2015, 362, fnv041. [Google Scholar] [CrossRef]
- Ahn, S.-Y.; Choi, M.; Jeong, D.; Park, S.; Park, H.; Jang, K.-S.; Choi, K.-Y. Synthesis and Chemical Composition Analysis of Protocatechualdehyde-Based Novel Melanin Dye by 15T FT-ICR: High Dyeing Performance on Soft Contact Lens. Dye. Pigment. 2019, 160, 546–554. [Google Scholar] [CrossRef]
- Chávez-Béjar, M.I.; Balderas-Hernandez, V.E.; Gutiérrez-Alejandre, A.; Martinez, A.; Bolívar, F.; Gosset, G. Metabolic Engineering of Escherichia Coli to Optimize Melanin Synthesis from Glucose. Microb. Cell Fact 2013, 12, 108. [Google Scholar] [CrossRef]
- Mejía-Caballero, A.; de Anda, R.; Hernández-Chávez, G.; Rogg, S.; Martinez, A.; Bolívar, F.; Castaño, V.M.; Gosset, G. Biosynthesis of Catechol Melanin from Glycerol Employing Metabolically Engineered Escherichia Coli. Microb. Cell Fact 2016, 15, 161. [Google Scholar] [CrossRef]
- Guo, J.; Rao, Z.; Yang, T.; Man, Z.; Xu, M.; Zhang, X. High-Level Production of Melanin by a Novel Isolate of Streptomyces Kathirae. FEMS Microbiol. Lett. 2014, 357, 85–91. [Google Scholar] [CrossRef]
- Ilakovac, B.; Voca, N.; Pezo, L.; Cerjak, M. Quantification and Determination of Household Food Waste and Its Relation to Sociodemographic Characteristics in Croatia. Waste Manag. 2020, 102, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ochoa, Á.; Leyva-Jiménez, F.J.; Pimentel-Moral, S.; del Carmen Villegas-Aguilar, M.; Alañón, M.E.; Segura-Carretero, A.; de la Luz Cádiz-Gurrea, M. Revalorisation of Agro-Industrial Wastes into High Value-Added Products. In Sustainable Bioconversion of Waste to Value Added Products, Advances in Science, Technology & Innovation; Inamuddin, K.A., Ed.; Springer International Publishing: Cham, Switzerland; Berlin, Germany, 2021; pp. 229–245. [Google Scholar] [CrossRef]
- Eurostat. Environmental Statistics and Accounts in Europe. Available online: https://ec.europa.eu/eurostat/documents/3217494/5723037/KS-32-10-283-EN.PDF.pdf/22a4889d-e6c9-4583-8d17-fb5104e7eec0 (accessed on 28 December 2022).
- Winkler, T.; Aschemann, R. Decreasing Greenhouse Gas Emissions of Meat Products Through Food Waste Reduction. A Framework for a Sustainability Assessment Approach. In Food Waste Reduction and Valorisation: Sustainability Assessment and Policy Analysis; Morone, P., Papendiek, F., Tartiu, V.E., Eds.; Springer International Publishing: Cham, Switzerland; Berlin, Germany, 2017; pp. 43–67. [Google Scholar] [CrossRef]
- FAO—News Article: Food Wastage: Key Facts and Figures. Available online: https://www.fao.org/news/story/en/item/196402/icode/ (accessed on 1 March 2023).
- Ritota, M.; Manzi, P. Pleurotus Spp. Cultivation on Different Agri-Food By-Products: Example of Biotechnological Application. Sustainability 2019, 11, 5049. [Google Scholar] [CrossRef]
- Torres-Valenzuela, L.S.; Ballesteros-Gómez, A.; Rubio, S. Green Solvents for the Extraction of High Added-Value Compounds from Agri-Food Waste. Food Eng. Rev. 2020, 12, 83–100. [Google Scholar] [CrossRef]
- Ben-Othman, S.; Jõudu, I.; Bhat, R. Bioactives from Agri-Food Wastes: Present Insights and Future Challenges. Molecules 2020, 25, 510. [Google Scholar] [CrossRef] [PubMed]
- Saadoun, J.H.; Bertani, G.; Levante, A.; Vezzosi, F.; Ricci, A.; Bernini, V.; Lazzi, C. Fermentation of Agri-Food Waste: A Promising Route for the Production of Aroma Compounds. Foods 2021, 10, 707. [Google Scholar] [CrossRef]
- Nikodinovic-Runic, J.; Martin, L.B.; Babu, R.; Blau, W.; O’Connor, K.E. Characterization of Melanin-Overproducing Transposon Mutants of Pseudomonas Putida F6. FEMS Microbiol. Lett. 2009, 298, 174–183. [Google Scholar] [CrossRef]
- Solano, F. Melanin and Melanin-Related Polymers as Materials with Biomedical and Biotechnological Applications—Cuttlefish Ink and Mussel Foot Proteins as Inspired Biomolecules. Int. J. Mol. Sci. 2017, 18, 1561. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Stark, R.E.; Casadevall, A. Fungal Melanin: What Do We Know About Structure? Front. Microbiol. 2015, 6, 1463. [Google Scholar] [CrossRef]
- Vahidzadeh, E.; Kalra, A.P.; Shankar, K. Melanin-Based Electronics: From Proton Conductors to Photovoltaics and Beyond. Biosens. Bioelectron. 2018, 122, 127–139. [Google Scholar] [CrossRef]
- Pralea, I.-E.; Moldovan, R.-C.; Petrache, A.-M.; Ilieș, M.; Hegheș, S.-C.; Ielciu, I.; Nicoară, R.; Moldovan, M.; Ene, M.; Radu, M.; et al. From Extraction to Advanced Analytical Methods: The Challenges of Melanin Analysis. Int. J. Mol. Sci. 2019, 20, 3943. [Google Scholar] [CrossRef]
- Camacho, E.; Vij, R.; Chrissian, C.; Prados-Rosales, R.; Gil, D.; O’Meally, R.N.; Cordero, R.J.B.; Cole, R.N.; McCaffery, J.M.; Stark, R.E.; et al. The Structural Unit of Melanin in the Cell Wall of the Fungal Pathogen Cryptococcus Neoformans. J. Biol. Chem. 2019, 294, 10471–10489. [Google Scholar] [CrossRef]
- El-Bialy, H.A.; El-Gamal, M.S.; Elsayed, M.A.; Saudi, H.A.; Khalifa, M.A. Microbial Melanin Physiology under Stress Conditions and Gamma Radiation Protection Studies. Radiat. Phys. Chem. 2019, 162, 178–186. [Google Scholar] [CrossRef]
- Bayram, S. Production, Purification, and Characterization of Streptomyces Sp. Strain MPPS2 Extracellular Pyomelanin Pigment. Arch. Microbiol. 2021, 203, 4419–4426. [Google Scholar] [CrossRef]
- Mattoon, E.R.; Cordero, R.J.B.; Casadevall, A. Fungal Melanins and Applications in Healthcare, Bioremediation and Industry. J. Fungi 2021, 7, 488. [Google Scholar] [CrossRef]
- Coelho, E.; Reis, T.A.; Cotrim, M.; Mullan, T.K.; Corrêa, B. Resistant Fungi Isolated from Contaminated Uranium Mine in Brazil Shows a High Capacity to Uptake Uranium from Water. Chemosphere 2020, 248, 126068. [Google Scholar] [CrossRef]
- Saini, A.S.; Melo, J.S. Biosorption of Uranium by Melanin: Kinetic, Equilibrium and Thermodynamic Studies. Bioresour. Technol. 2013, 149, 155–162. [Google Scholar] [CrossRef]
- Blasi, B.; Poyntner, C.; Rudavsky, T.; Prenafeta-Boldú, F.X.; de Hoog, S.; Tafer, H.; Sterflinger, K. Pathogenic Yet Environmentally Friendly? Black Fungal Candidates for Bioremediation of Pollutants. Geomicrobiol. J. 2016, 33, 308–317. [Google Scholar] [CrossRef]
- Xu, C.; Li, J.; Yang, L.; Shi, F.; Yang, L.; Ye, M. Antibacterial Activity and a Membrane Damage Mechanism of Lachnum YM30 Melanin against Vibrio Parahaemolyticus and Staphylococcus Aureus. Food Control. 2017, 73, 1445–1451. [Google Scholar] [CrossRef]
- Carocho, M.; Morales, P.; Ferreira, I.C.F.R. Antioxidants: Reviewing the Chemistry, Food Applications, Legislation and Role as Preservatives. Trends Food Sci. Technol. 2018, 71, 107–120. [Google Scholar] [CrossRef]
- Solano, F. Photoprotection versus Photodamage: Updating an Old but Still Unsolved Controversy about Melanin: Photoprotection versus Photodamage. Polym. Int. 2016, 65, 1276–1287. [Google Scholar] [CrossRef]
- Łopusiewicz, Ł.; Kwiatkowski, P.; Drozłowska, E.; Trocer, P.; Kostek, M.; Śliwiński, M.; Polak-Śliwińska, M.; Kowalczyk, E.; Sienkiewicz, M. Preparation and Characterization of Carboxymethyl Cellulose-Based Bioactive Composite Films Modified with Fungal Melanin and Carvacrol. Polymers 2021, 13, 499. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Rhim, J.-W. Preparation of Carrageenan-Based Functional Nanocomposite Films Incorporated with Melanin Nanoparticles. Colloids Surf. B Biointerfaces 2019, 176, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Łopusiewicz, Ł.; Jędra, F.; Mizielińska, M. New Poly(Lactic Acid) Active Packaging Composite Films Incorporated with Fungal Melanin. Polymers 2018, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Li, L.; Yu, S.; Liu, J.; Shi, J. High Performance of Alginate/Polyvinyl Alcohol Composite Film Based on Natural Original Melanin Nanoparticles Used as Food Thermal Insulating and UV–Vis Block. Carbohydr. Polym. 2020, 233, 115884. [Google Scholar] [CrossRef] [PubMed]
- Ezati, P.; Rhim, J.-W.; Moradi, M.; Tajik, H.; Molaei, R. CMC and CNF-Based Alizarin Incorporated Reversible PH-Responsive Color Indicator Films. Carbohydr. Polym. 2020, 246, 116614. [Google Scholar] [CrossRef]
- Wang, H.; Qian, J.; Ding, F. Emerging Chitosan-Based Films for Food Packaging Applications. J. Agric. Food Chem. 2018, 66, 395–413. [Google Scholar] [CrossRef]
- Motelica, L.; Ficai, D.; Ficai, A.; Truşcă, R.-D.; Ilie, C.-I.; Oprea, O.-C.; Andronescu, E. Innovative Antimicrobial Chitosan/ZnO/Ag NPs/Citronella Essential Oil Nanocomposite—Potential Coating for Grapes. Foods 2020, 9, 1801. [Google Scholar] [CrossRef]
- Motelica, L.; Ficai, D.; Oprea, O.; Ficai, A.; Trusca, R.-D.; Andronescu, E.; Holban, A.M. Biodegradable Alginate Films with ZnO Nanoparticles and Citronella Essential Oil—A Novel Antimicrobial Structure. Pharmaceutics 2021, 13, 1020. [Google Scholar] [CrossRef]
- Motelica, L.; Ficai, D.; Oprea, O.-C.; Ficai, A.; Ene, V.-L.; Vasile, B.-S.; Andronescu, E.; Holban, A.-M. Antibacterial Biodegradable Films Based on Alginate with Silver Nanoparticles and Lemongrass Essential Oil–Innovative Packaging for Cheese. Nanomaterials 2021, 11, 2377. [Google Scholar] [CrossRef]
- Garrison, T.F.; Murawski, A.; Quirino, R.L. Bio-Based Polymers with Potential for Biodegradability. Polymers 2016, 8, 262. [Google Scholar] [CrossRef]
- Bang, Y.; Shankar, S.; Rhim, J. Preparation of Polypropylene/Poly (Butylene Adipate-co-terephthalate) Composite Films Incorporated with Melanin for Prevention of Greening of Potatoes. Packag. Technol. Sci. 2020, 33, pts.2525. [Google Scholar] [CrossRef]
- Shankar, S.; Wang, L.-F.; Rhim, J.-W. Effect of Melanin Nanoparticles on the Mechanical, Water Vapor Barrier, and Antioxidant Properties of Gelatin-Based Films for Food Packaging Application. Food Packag. Shelf Life 2019, 21, 100363. [Google Scholar] [CrossRef]
- Kiran, G.S.; Jackson, S.A.; Priyadharsini, S.; Dobson, A.D.W.; Selvin, J. Synthesis of Nm-PHB (Nanomelanin-Polyhydroxy Butyrate) Nanocomposite Film and Its Protective Effect against Biofilm-Forming Multi Drug Resistant Staphylococcus Aureus. Sci. Rep. 2017, 7, 9167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Organisms | Color | Oxidating Substrate | References |
|---|---|---|---|---|
| Eumelanin | Animal, human, bacteria, fungi | Black—Brown | L-tyrosine | [6] |
| Pheomelanin | Animal (hair, feathers) | Yellow—Red | L-tyrosine and L-cysteine | [6,7] |
| Neuromelanin | Animal | Black—Brown | [4] | |
| Allomelanin | Plants, fungi, bacteria | Black—Brown | Catechol, caffeic acid, dihydroxynaphthalene, tetrahydroxynaphthalene, protocatechualdehyde, 4-hydroxyphenylacetic acid, γ-glutaminyl-4-hydroxybenzene | [5] |
| Catechol melanin | Plants | Black—Brown | ||
| DHN-melanin | Fungi, bacteria | Black—Brown | ||
| Pyomelanin | Fungi, bacteria | Black—Brown | Homogentisic acid | [8] |
| Feedstock | Microbial Strain | Fermentation Time (Days) | Melanin (g/L) | Melanin Type | References |
|---|---|---|---|---|---|
| Renewable waste and byproduct streams | |||||
| Rice husk, rice flour and banana | Streptomyces gresiorubens DKR4 | [34] | |||
| Carrot peel extract | Aureobasidium pullulans | 15 | 4.4 | [35] | |
| Wheat bran extract | Auricularia auricula | 5 | 0.519 | [36] | |
| Fruit waste extract | Bacillus safensis | [37] | |||
| Marine residues | Streptomyces roseochromogenes ATCC 13400 | 39.4 | [38] | ||
| Pomegranate pulp | Aspergillus carbonarius M333 | 15 | [39] | ||
| Vegetable cabbage waste | Pseudomonas sp. | 48–72 h | [12] | ||
| Commercial-based fermentation media | |||||
| Dextrose, yeast extract and peptone | Aspergillus fumigatus AFGRD105 | 0.007 | [40] | ||
| Glucose, yeast extract and peptone | Amorphotheca resinae KUC3009 | 14 | 4.5 | eumelanin | [41] |
| Voguels salt water | Hortaea werneckii | 7 | 0.938 | [42] | |
| Amylodextrine, yeast extract and NaCl | Streptomyces kathirae | 13.7 | |||
| Glucose, yeast extract and peptone | Armillaria cepistipes (Empa strain 655) | 161 | 27.98 | eumelanin | [5] |
| Genetically modified strains | |||||
| Bacillus thuringiensis 4D11 | [43] | ||||
| S. kathirae SC-1 | 28.8 | [44] | |||
| Caffeic acid | E. coli BL21(DE3) | 12 h | 0.17 | eumelanin | [45] |
| Glucose | E. coli | 120 h | 3.2 | eumelanin | [46] |
| Glycerol | E. coli W3110 | 1.2 | catechol-melanin | [47] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsouko, E.; Tolia, E.; Sarris, D. Microbial Melanin: Renewable Feedstock and Emerging Applications in Food-Related Systems. Sustainability 2023, 15, 7516. https://doi.org/10.3390/su15097516
Tsouko E, Tolia E, Sarris D. Microbial Melanin: Renewable Feedstock and Emerging Applications in Food-Related Systems. Sustainability. 2023; 15(9):7516. https://doi.org/10.3390/su15097516
Chicago/Turabian StyleTsouko, Erminta, Eirini Tolia, and Dimitris Sarris. 2023. "Microbial Melanin: Renewable Feedstock and Emerging Applications in Food-Related Systems" Sustainability 15, no. 9: 7516. https://doi.org/10.3390/su15097516