
Evaluation of Olive Varieties Resistance for Sustainable Management of Verticillium Wilt

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Pathogen Preparation
2.3. Plant Inoculation
2.4. Disease Assessment and Plant Growth
2.5. DNA Extraction
2.6. Pathogen qPCR Quantification
2.7. Cumulative Stress Response Index (CSRI)
- CSRI = cumulative stress response index,
- PH = plant height,
- PFW = plant fresh weight,
- PS = plant survival,
- t = treatment,
- c = control.
2.8. Statistics
3. Results
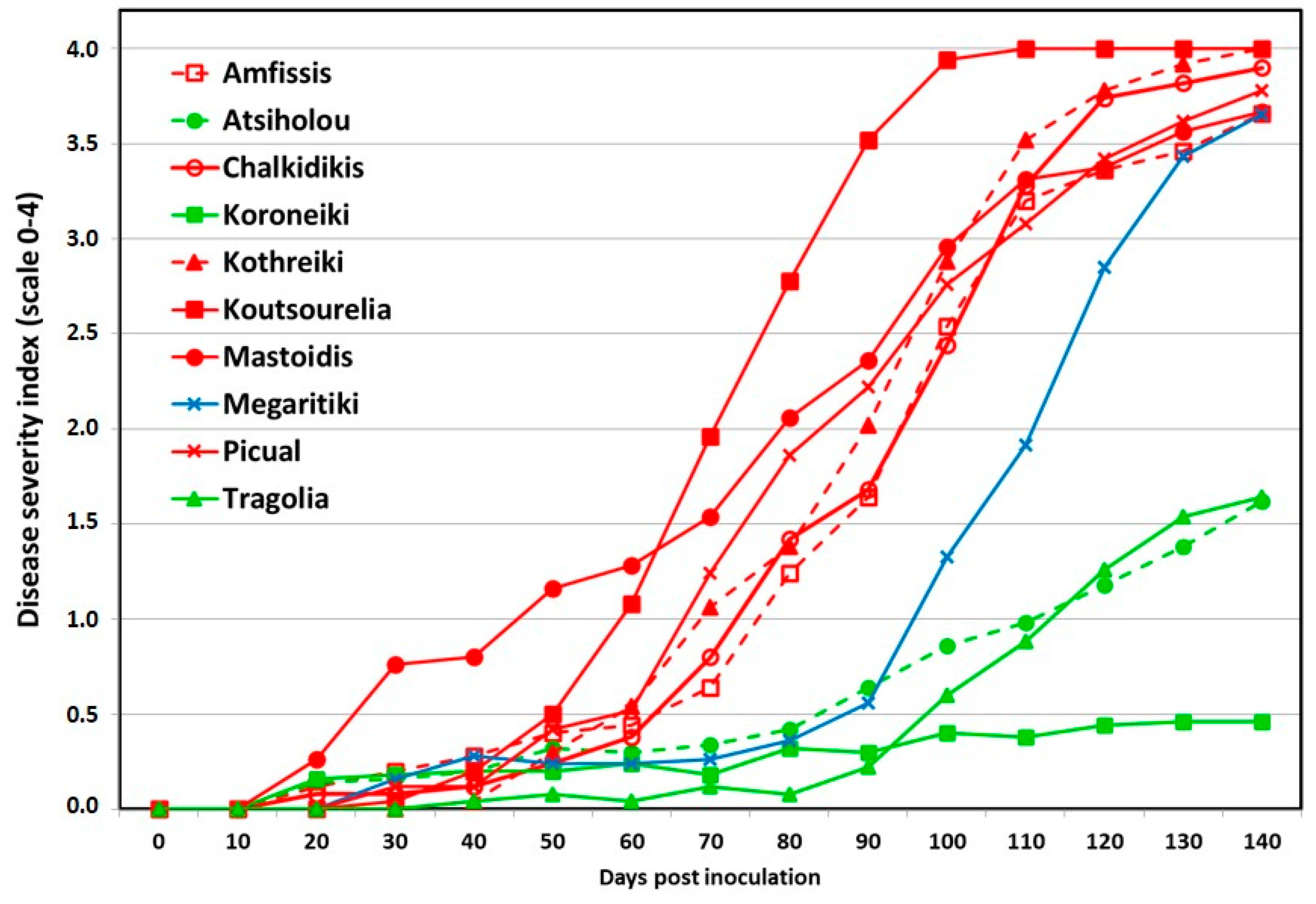
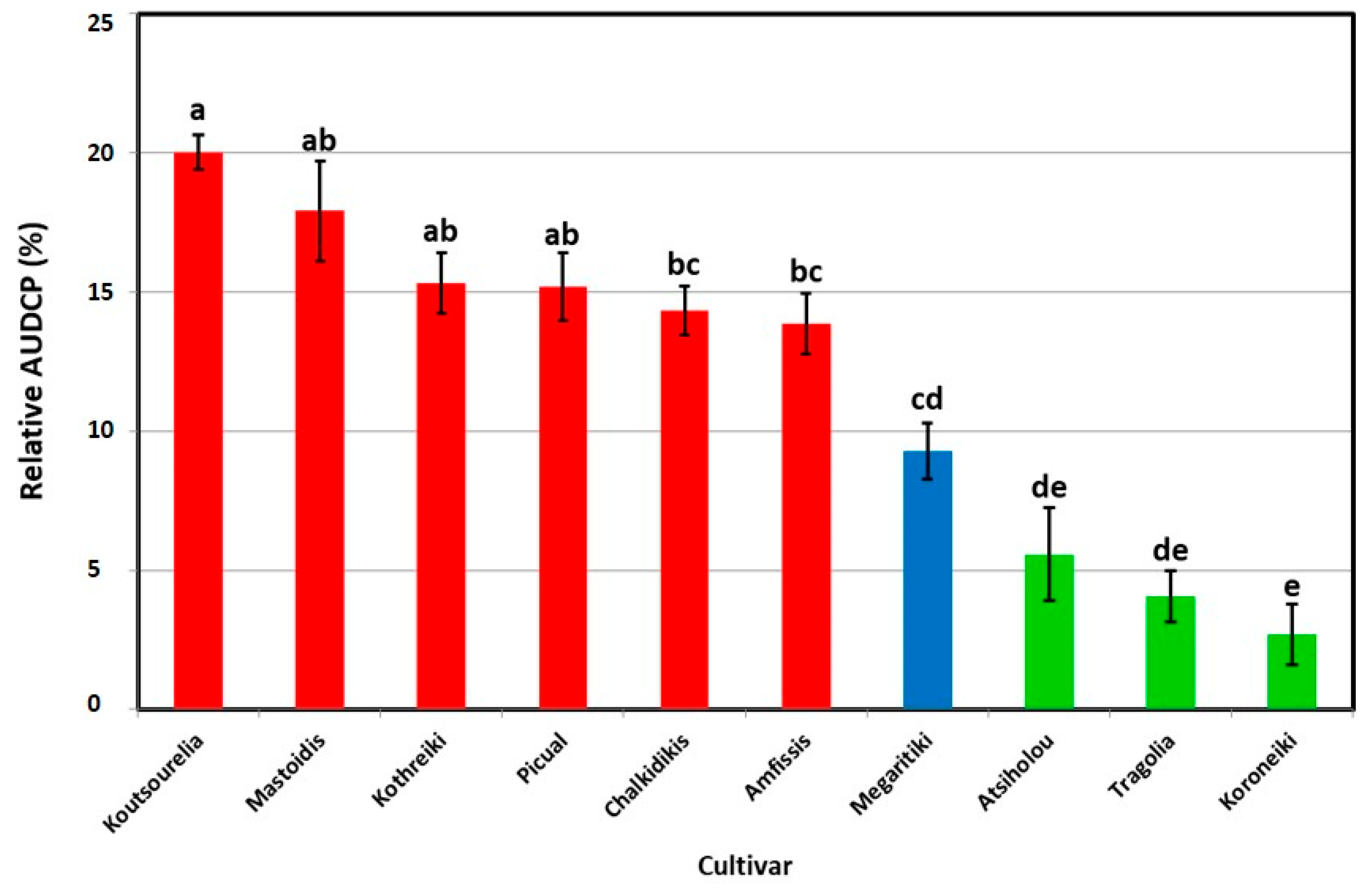
3.1. Symptom Development
3.2. Effects of V. dahliae Inoculation on Plant Growth
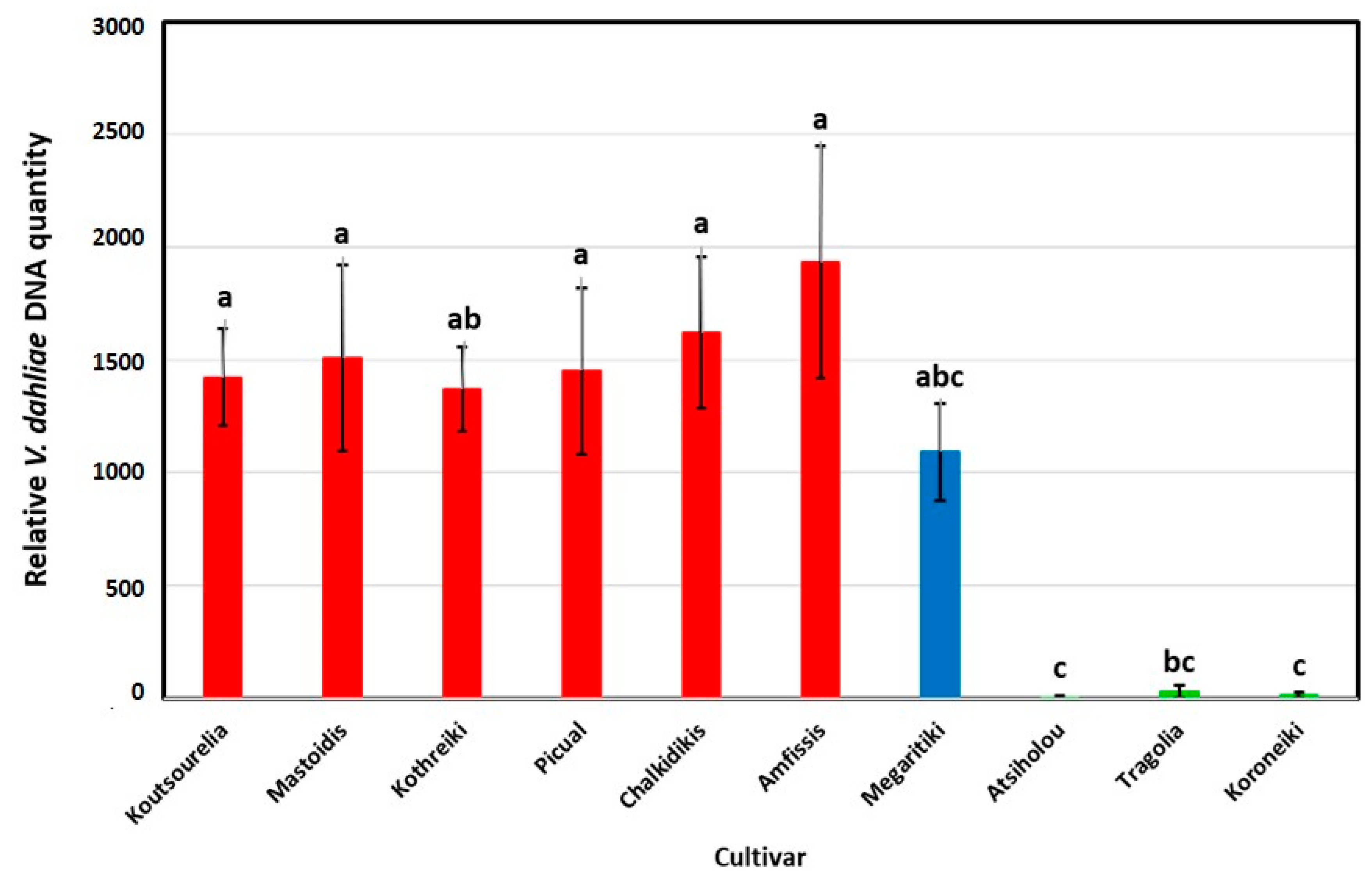
3.3. Verticillium Dahliae qPCR Quantification
3.4. Cumulative Stress Response Index (CSRI)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trapero, A.; Blanco, M.A. Diseases. In Olive Growing; Barranco, D., Fernández-Escobar, R., Rallo, L., Eds.; Junta de Andalucía/Mundi-Prensa/RIRDC/AOA: Pendle Hill, NSW, Australia, 2010; pp. 521–578. [Google Scholar]
- Jiménez-Díaz, R.M.; Cirulli, M.; Bubici, G.; Jiménez-Gasco, L.M.; Antoniou, P.P.; Tjamos, E.C. Verticillium wilt, a major threat to olive production: Current status and future prospects for its management. Plant Dis. 2012, 96, 304–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soilborne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier Academic Press: Cambridge, CA, USA, 2005. [Google Scholar]
- Montes-Osuna, N.; Mercado-Blanco, J. Verticillium Wilt of Olive and Its Control: What Did We Learn during the Last Decade? Plants 2020, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Newton, A.C. Climate changes, plant diseases and food security: An overview. Plant Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Antoniou, P.P.; Markakis, E.A.; Tjamos, S.E.; Paplomatas, E.J.; Tjamos, E.C. Novel methodologies in screening and selecting olive varieties and rootstocks for resistance to Verticillium dahliae. Eur. J. Plant Pathol. 2008, 122, 549–560. [Google Scholar] [CrossRef]
- Bubici, G.; Cirulli, M. Control of Verticillium wilt of olive by resistant rootstocks. Plant Soil 2012, 352, 363–376. [Google Scholar] [CrossRef]
- Colella, C.; Miacola, C.; Amenduni, M.; D’Amico, M.; Bubici, G.; Cirulli, M. Sources of Verticillium wilt resistance in wild olive germplasm from the Mediterranean region. Plant Pathol. 2008, 57, 533–539. [Google Scholar] [CrossRef]
- Erten, L.; Yıldız, M. Screening for resistance of Turkish olive varieties and clonal rootstocks to Verticillium wilt. Phytoparasitica 2011, 39, 83–92. [Google Scholar] [CrossRef]
- Garcia-Ruiz, G.M.; Trapero, C.; Del Rio, C.; Lopez-Escudero, F.J. Evaluation of resistance of Spanish olive varieties to Verticillium dahliae in inoculations conducted in greenhouse. Phytoparasitica 2014, 42, 205–212. [Google Scholar] [CrossRef]
- Lopez-Escudero, F.J.; Del Rio, C.; Caballero, J.M.; Blanco-Lopez, M.A. Evaluation of olive varieties for resistance to Verticillium dahliae. Eur. J. Plant Pathol. 2004, 110, 79–85. [Google Scholar] [CrossRef]
- López-Escudero, F.J.; Blanco-López, M.A.; del Río, C.; Caballero, J.M. Response of olive varieties to stem puncture inoculation with a defoliating pathotype of Verticillium dahliae. HortScience 2007, 42, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Paplomatas, E.J.; Tjamos, E.C. Symptom development, pathogen isolation, and real-time qPCR quantification as factors for evaluating the resistance of olive varieties to Verticillium pathotypes. Eur. J. Plant Pathol. 2009, 124, 603–611. [Google Scholar] [CrossRef]
- Martos Moreno, C.; López-Escudero, F.J.; Blanco López, M.A. Resistance of olive varieties to the defoliating pathotype of Verticillium dahliae. HortScience 2006, 41, 1313–1316. [Google Scholar] [CrossRef] [Green Version]
- Sesli, M.; Onan, E.; Oden, S.; Yener, H.; Yegenoglu, E.D. Resistance of olive varieties to Verticillium dahliae. Sci. Res. Essays 2010, 5, 1561–1565. [Google Scholar]
- Valverde Caballero, P.; Trapero Ramírez, C.; Barranco Navero, D.; López-Escudero, F.J.; Gordon Bermúdez-Coronel, A.; Díez, C.M. Assessment of Maternal Effects and Genetic Variability in Resistance to Verticillium dahliae in Olive Progenies. Plants 2021, 10, 1534. [Google Scholar] [CrossRef]
- Song, R.; Li, J.; Xie, C.; Jian, W.; Yang, X. An Overview of the Molecular Genetics of Plant Resistance to the Verticillium Wilt Pathogen Verticillium dahliae. Int. J. Mol. Sci. 2020, 21, 1120. [Google Scholar] [CrossRef] [Green Version]
- Cardoni, M.; Mercado-Blanco, J.; Villar, R. Functional Traits of Olive Varieties and Their Relationship with the Tolerance Level towards Verticillium Wilt. Plants 2021, 10, 1079. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Available online: https://www.nationmaster.com/nmx/ranking/olives-production-fao (accessed on 15 December 2021).
- Tjamos, E.C.; Thanassoulopoulos, C.C.; Biris, D.A. Resistance evaluation to Verticillium dahliae of olive rootstocks. In Proceedings of the 3rd National Phytopathological Conference of the Hellenic Phytopathological Society, Volos, Greece, 16–18 October 1985. [Google Scholar]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Paplomatas, E.J.; Tjamos, E.C. Phenolic responses of resistant and susceptible olive varieties induced by defoliating and non-defoliating Verticillium dahliae pathotypes. Plant Dis. 2010, 94, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Pegg, G.F.; Brady, B.L. Verticillium Wilts, 1st ed.; CABI Publishing: Wallingford, UK, 2002. [Google Scholar]
- Markakis, E.A.; Koubouris, G.C.; Sergentani, C.K.; Ligoxigakis, E.K. Evaluation of Greek grapevine varieties for resistance to Phaeomoniella chlamydospora. Eur. J. Plant Pathol. 2017, 149, 277–283. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology, 1st ed.; Wiley-Interscience: New York, NY, USA, 1990. [Google Scholar]
- Korolev, N.; Perez-Artes, E.; Bejarano-Alcazar, J.; Rodrıguez-Jurado, D.; Katan, J.; Katan, T.; Jimenez-Diaz, R.M. Comparative study of genetic diversity and pathogenicity among populations of Verticillium dahliae from cotton in Spain and Israel. Eur. J. Plant Pathol. 2001, 107, 443–456. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Koubouris, G.C.; Metzidakis, I.T.; Vasilakakis, M.D. Impact of temperature on olive (Olea europaea L.) pollen performance in relation to relative humidity and genotype. Environ. Exp. Bot. 2009, 67, 209–214. [Google Scholar] [CrossRef]
- Markakis, E.A.; Ligoxigakis, E.K.; Roussos, P.A.; Sergentani, C.K.; Kavroulakis, N.; Roditakis, E.N.; Koubouris, G.C. Differential susceptibility responses of Greek olive cultivars to Fomitiporia mediterranea. Eur. J. Plant Pathol. 2019, 153, 1055–1066. [Google Scholar] [CrossRef]
- Tjamos, E.C. Prospects and strategies in controlling Verticillium wilt of olive. Bull. OEPP/EPPO Bull 1993, 23, 505–512. [Google Scholar] [CrossRef]
- Arias-Calderón, R.; León, L.; Bejarano-Alcázar, J.; Belaj, A.; de la Rosa, R.; Rodríguez-Jurado, D. Resistance to Verticillium wilt in olive progenies from open-pollination. Sci. Hortic. 2015, 185, 34–42. [Google Scholar] [CrossRef]
- Trapero, C.; Rallo, L.; López-Escudero, F.J.; Barranco, D.; Dıez, C.M. Variability and selection of verticillium wilt resistant genotypes in cultivated olive and in the Olea genus. Plant Pathol. 2015, 64, 890–900. [Google Scholar] [CrossRef]
- Trapero, C.; Díez, C.M.; Rallo, L.; Barranco, D.; López-Escudero, F.J. Effective inoculation methods to screen for resistance to Verticillium wilt in olive. Sci. Hortic. 2013, 162, 252–259. [Google Scholar] [CrossRef]
- Serrano, A.; Rodríguez-Jurado, D.; Román, B.; Bejarano-Alcázar, J.; De la Rosa, R.; León, L. Verticillium Wilt Evaluation of Olive Breeding Selections Under Semi-Controlled Conditions. Plant Dis. 2021, 105, 1781–1790. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | F Values a | ||||||
|---|---|---|---|---|---|---|---|
| df b | DI | FDS | M | RAUDPC | H | W | |
| Replication | 4 | 0.113 | 0.687 | 0.034 | 0.471 | 0.956 | 1.647 |
| Variety | 9 | 33.410 *** | 48.778 *** | 29.984 *** | 28.114 *** | 32.850 *** | 43.984 *** |
| Replication × Variety | 36 | - | 1.234 | - | 1.576 | 1.178 | 1.574 |
| Variety | Disease Parameters a | Growth Parameters x | |||
|---|---|---|---|---|---|
| Disease Incidence (%) b | Final Disease Severity (Scale 0–4) c | Mortality (%) d | Plant Height (cm) y | Plant Fresh Weight (gr) z | |
| Amfissis | 100.00 ± 0.00 a | 3.66 ± 0.18 a | 84.00 ± 9.80 a | 30.28 ± 0.72 c | 2.43 ± 0.35 b |
| Atsiholou | 60.00 ± 8.94 b | 1.62 ± 0.34 b | 32.00 ± 10.20 b | 39.28 ± 1.00 a | 8.09 ± 0.87 a |
| Chalkidikis | 100.00 ± 0.00 a | 3.90 ± 0.10 a | 96.00 ± 4.00 a | 28.68 ± 0.61 c | 2.13 ± 0.19 b |
| Koroneiki | 24.00 ± 7.48 c | 0.46 ± 0.18 c | 0.00 ± 0.00 c | 40.08 ± 0.92 a | 9.75 ± 0.51 a |
| Kothreiki | 100.00 ± 0.00 a | 4.00 ± 0.00 a | 100.00 ± 0.00 a | 29.52 ± 0.49 c | 2.46 ± 0.12 b |
| Koutsourelia | 100.00 ± 0.00 a | 4.00 ± 0.00 a | 100.00 ± 0.00 a | 36.68 ± 1.18 a,b | 3.09 ± 0.20 b |
| Mastoidis | 96.00 ± 4.00 a | 3.67 ± 0.18 a | 88.00 ± 8.00 a | 34.42 ± 0.65 b | 2.78 ± 0.41 b |
| Megaritiki | 100.00 ± 0.00 a | 3.65 ± 0.12 a | 80.00 ± 10.95 a | 29.00 ± 0.62 c | 3.62 ± 0.25 b |
| Picual | 100.00 ± 0.00 a | 3.78 ± 0.11 a | 88.00 ± 4.90 a | 36.80 ± 0.91 a,b | 2.46 ± 0.27 b |
| Tragolia | 64.00 ± 7.48 b | 1.64 ± 0.32 b | 24.00 ± 4.00 b,c | 38.96 ± 0.79 a | 8.50 ± 0.84 a |
| Genotypes | Cumulative Stress Response Index (CSRI) | Rank a | Characterization |
|---|---|---|---|
| Amfissis | −184.52 | 7 | Sensitive |
| Atsiholou | −79.66 | 3 | Tolerant |
| Chalkidikis | −219.02 | 10 | Sensitive |
| Koroneiki | −33.64 | 1 | Tolerant |
| Kothreiki | −108.95 | 5 | Intermediate |
| Koutsourelia | −94.41 | 4 | Intermediate |
| Mastoidis | −188.72 | 8 | Sensitive |
| Megaritiki | −144.54 | 6 | Intermediate |
| Picual | −207.51 | 9 | Sensitive |
| Tragolia | −69.37 | 2 | Tolerant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markakis, E.A.; Krasagakis, N.; Manolikaki, I.; Papadaki, A.A.; Kostelenos, G.; Koubouris, G. Evaluation of Olive Varieties Resistance for Sustainable Management of Verticillium Wilt. Sustainability 2022, 14, 9342. https://doi.org/10.3390/su14159342
Markakis EA, Krasagakis N, Manolikaki I, Papadaki AA, Kostelenos G, Koubouris G. Evaluation of Olive Varieties Resistance for Sustainable Management of Verticillium Wilt. Sustainability. 2022; 14(15):9342. https://doi.org/10.3390/su14159342
Chicago/Turabian StyleMarkakis, Emmanouil A., Nikolaos Krasagakis, Ioanna Manolikaki, Anastasia A. Papadaki, Georgios Kostelenos, and Georgios Koubouris. 2022. "Evaluation of Olive Varieties Resistance for Sustainable Management of Verticillium Wilt" Sustainability 14, no. 15: 9342. https://doi.org/10.3390/su14159342