
Semi-Automated Recording of Facial Sensitivity in Rat Demonstrates Antinociceptive Effects of the Anti-CGRP Antibody Fremanezumab
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
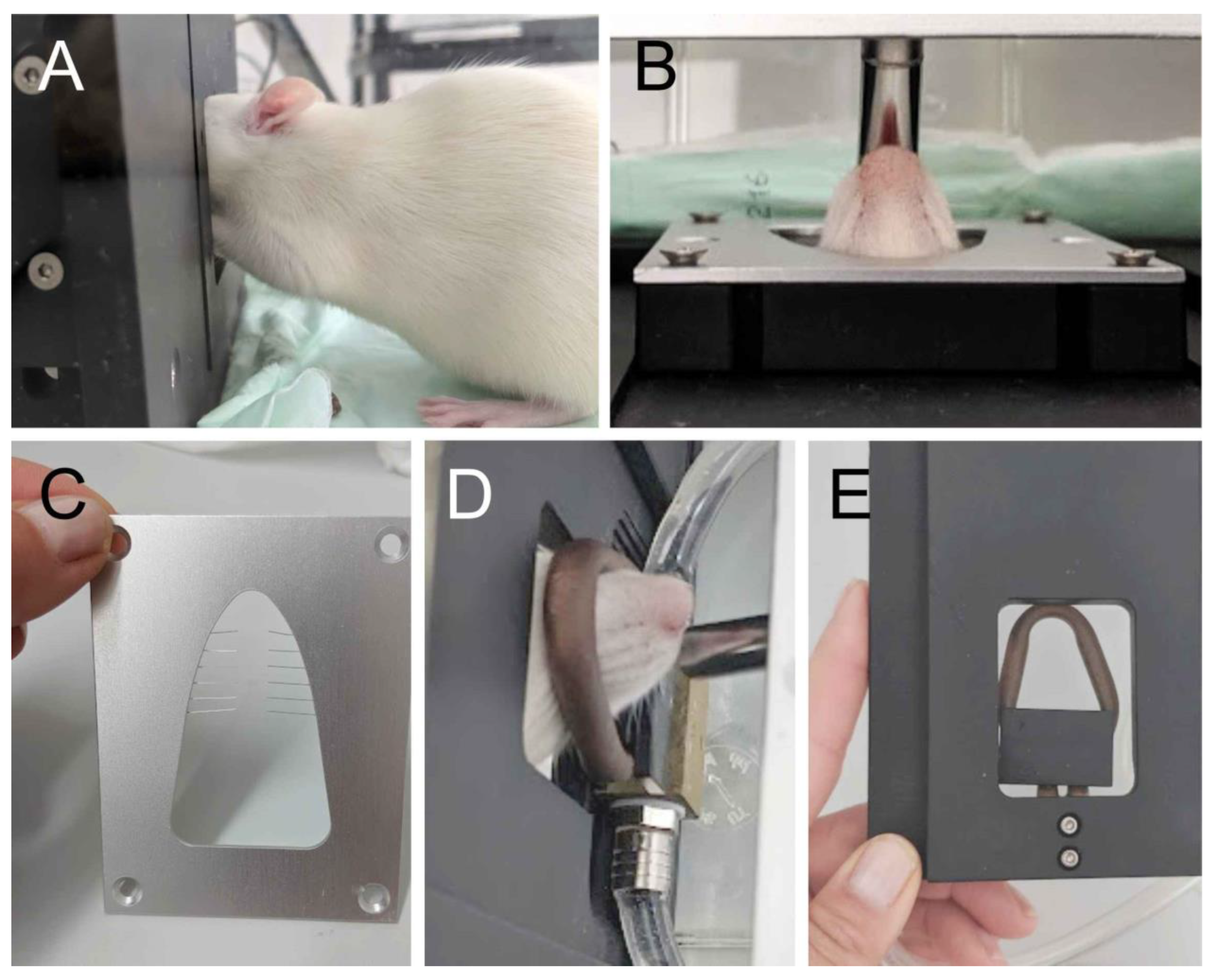
2.2. Preparation of Animals for Behavioral Tests
2.3. Test Cage and Recording Device
2.4. Mechanical and Thermal Stimulation Device
2.5. Test Procedures
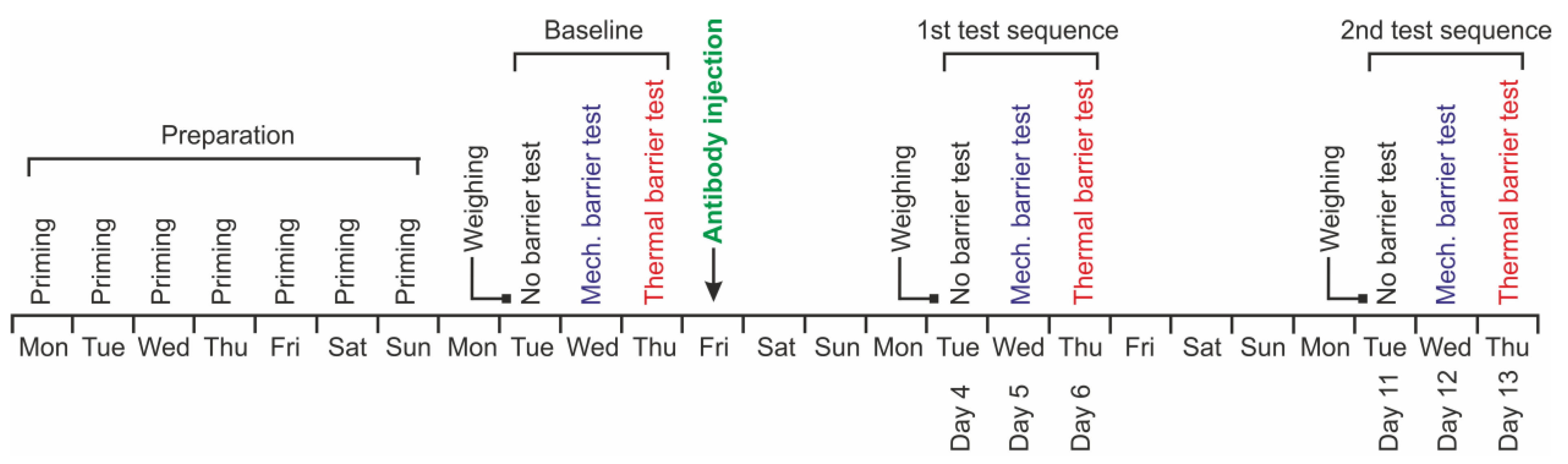
2.6. Test Sequence
2.7. Injection of Antibodies
2.8. Data Calculation and Statistics
3. Results
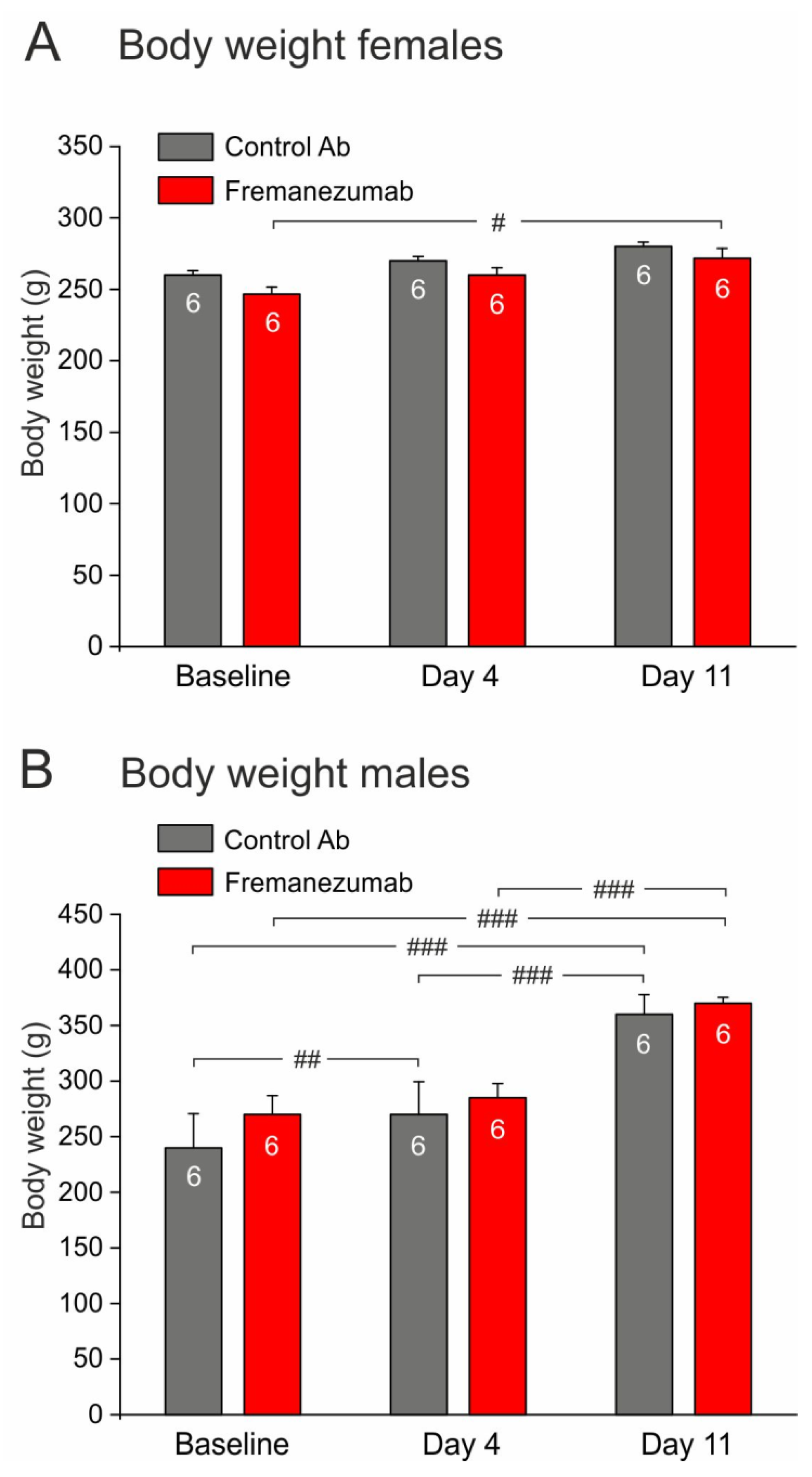
3.1. Allocation of Animals and Tolerability of Treatments
3.2. Baseline Experiments
3.3. Experiments after Antibody Injection
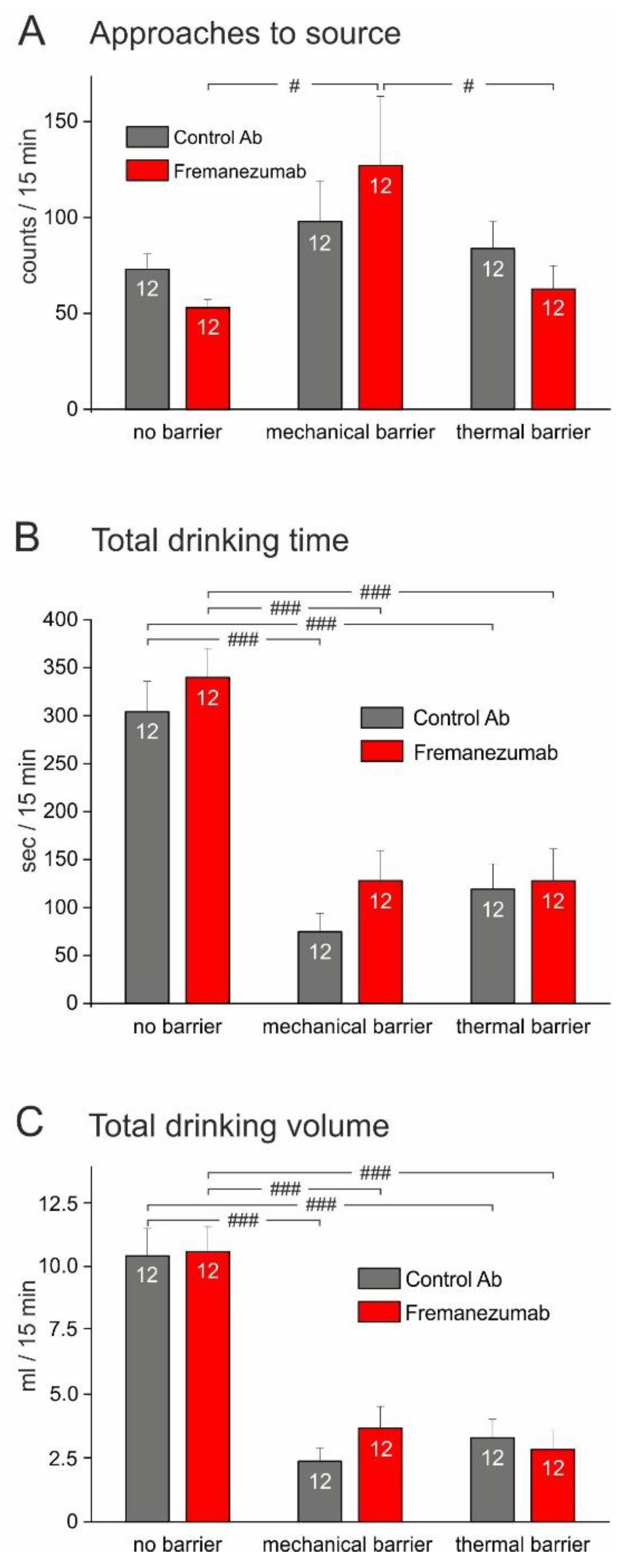
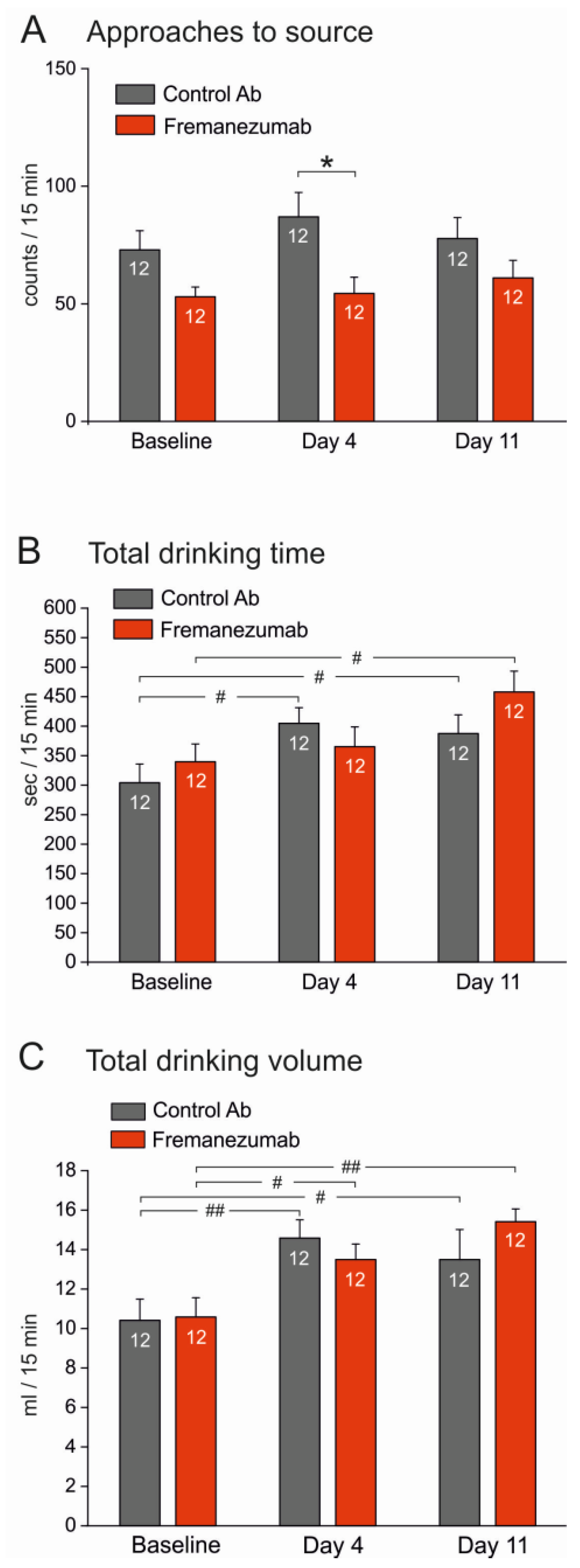
3.3.1. Experiments without Barrier (Figure 5)
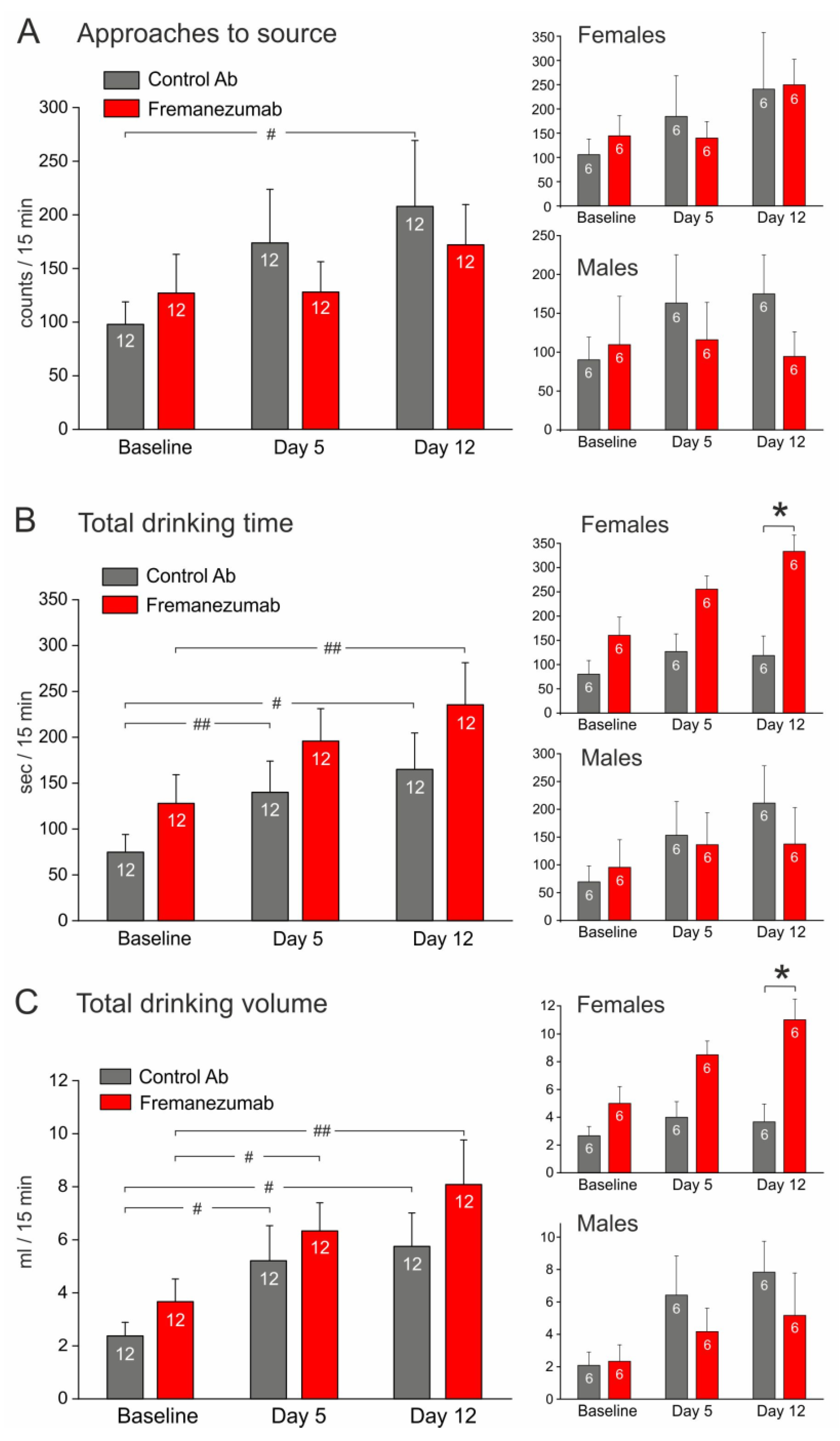
3.3.2. Experiments with Mechanical Barrier (Figure 6)
3.3.3. Experiments with Thermal Barrier (Figure 7)
4. Discussion
4.1. Test Device
4.2. Proposed Mechanisms of Anti-CGRP Antibody Effects
4.3. Sex Differences
4.4. Clinical Relevance and Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Edvinsson, L.; Ekman, R.; Jansen, I.; McCulloch, J.; Uddman, R. Calcitonin Gene-Related Peptide and Cerebral Blood Vessels: Distribution and Vasomotor Effects. J. Cereb. Blood Flow Metab. 1987, 7, 720–728. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Goadsby, P.J. Neuropeptides in Migraine and Cluster Headache. Cephalalgia 1994, 14, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Lassen, L.H.; Haderslev, P.A.; Jacobsen, V.B.; Iversen, H.K.; Sperling, B.; Olesen, J. Cgrp May Play A Causative Role in Migraine. Cephalalgia Int. J. Headache 2002, 22, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.M.; Hauge, A.W.; Olesen, J.; Ashina, M. Calcitonin gene-related peptide triggers migraine-like attacks in patients with migraine with aura. Cephalalgia 2010, 30, 1179–1186. [Google Scholar] [CrossRef]
- Benemei, S.; Cortese, F.; Labastida-Ramírez, A.; Marchese, F.; Pellesi, L.; Romoli, M.; Vollesen, A.L.; Lampl, C.; Ashina, M.; School of Advanced Studies of the European Headache Federation (EHF-SAS). Triptans and CGRP blockade—Impact on the cranial vasculature. J. Headache Pain 2017, 18, 103. [Google Scholar] [CrossRef]
- Charles, A.; Pozo-Rosich, P. Targeting calcitonin gene-related peptide: A new era in migraine therapy. Lancet 2019, 394, 1765–1774. [Google Scholar] [CrossRef]
- Melo-Carrillo, A.; Strassman, A.M.; Nir, R.-R.; Schain, A.J.; Noseda, R.; Stratton, J.; Burstein, R. Fremanezumab—A Humanized Monoclonal Anti-CGRP Antibody—Inhibits Thinly Myelinated (Aδ) But Not Unmyelinated (C) Meningeal Nociceptors. J. Neurosci. 2017, 37, 10587–10596. [Google Scholar] [CrossRef]
- Melo-Carrillo, A.; Noseda, R.; Nir, R.-R.; Schain, A.J.; Stratton, J.; Strassman, A.M.; Burstein, R. Selective Inhibition of Trigeminovascular Neurons by Fremanezumab: A Humanized Monoclonal Anti-CGRP Antibody. J. Neurosci. 2017, 37, 7149–7163. [Google Scholar] [CrossRef]
- Burstein, R.; Yamamura, H.; Malick, A.; Strassman, A.M. Chemical Stimulation of the Intracranial Dura Induces Enhanced Responses to Facial Stimulation in Brain Stem Trigeminal Neurons. J. Neurophysiol. 1998, 79, 964–982. [Google Scholar] [CrossRef]
- Edelmayer, R.M.; Ossipov, M.H.; Porreca, F. An Experimental Model of Headache-Related Pain. Methods Mol. Biol. 2012, 851, 109–120. [Google Scholar] [CrossRef]
- Zhang, Z.; Winborn, C.S.; de Prado, B.M.; Russo, A.F. Sensitization of Calcitonin Gene-Related Peptide Receptors by Receptor Activity-Modifying Protein-1 in the Trigeminal Ganglion. J. Neurosci. 2007, 27, 2693–2703. [Google Scholar] [CrossRef]
- De Prado, B.M.; Hammond, D.; Russo, A.F. Genetic Enhancement of Calcitonin Gene-Related Peptide-Induced Central Sensitization to Mechanical Stimuli in Mice. J. Pain 2009, 10, 992–1000. [Google Scholar] [CrossRef]
- Rea, B.J.; Wattiez, A.-S.; Waite, J.S.; Castonguay, W.C.; Schmidt, C.M.; Fairbanks, A.M.; Robertson, B.R.; Brown, C.J.; Mason, B.N.; Moldovan-Loomis, M.-C.; et al. Peripherally administered calcitonin gene–related peptide induces spontaneous pain in mice: Implications for migraine. Pain 2018, 159, 2306–2317. [Google Scholar] [CrossRef]
- Mogil, J.S.; Miermeister, F.; Seifert, F.; Strasburg, K.; Zimmermann, K.; Reinold, H.; Austin, J.-S.; Bernardini, N.; Chesler, E.J.; Hofmann, H.A.; et al. Variable sensitivity to noxious heat is mediated by differential expression of the CGRP gene. Proc. Natl. Acad. Sci. USA 2005, 102, 12938–12943. [Google Scholar] [CrossRef]
- Landini, L.; Janal, M.N.; Simone, L.P.; Pierangelo, G.; Romina, N. Migraine-provoking substances evoke periorbital allodynia in mice. J. Headache Pain 2019, 20, 18. [Google Scholar] [CrossRef]
- Capuano, A.; Greco, M.C.; Navarra, P.; Tringali, G. Correlation between algogenic effects of calcitonin-gene-related peptide (CGRP) and activation of trigeminal vascular system, in an in vivo experimental model of nitroglycerin-induced sensitization. Eur. J. Pharmacol. 2014, 740, 97–102. [Google Scholar] [CrossRef]
- Avona, A.; Burgos-Vega, C.; Burton, M.D.; Akopian, A.N.; Price, T.J.; Dussor, G. Dural Calcitonin Gene-Related Peptide Produces Female-Specific Responses in Rodent Migraine Models. J. Neurosci. 2019, 39, 4323–4331. [Google Scholar] [CrossRef]
- Wattiez, A.-S.; Wang, M.; Russo, A.F. CGRP in Animal Models of Migraine. In Calcitonin Gene-Related Peptide (CGRP) Mechanisms; Brain, S.D., Geppetti, P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 85–107. [Google Scholar]
- Gazerani, P.; Dong, X.; Wang, M.; Kumar, U.; Cairns, B.E. Sensitization of rat facial cutaneous mechanoreceptors by activation of peripheral N-methyl-d-aspartate receptors. Brain Res. 2010, 1319, 70–82. [Google Scholar] [CrossRef]
- Wei, X.; Edelmayer, R.M.; Yan, J.; Dussor, G. Activation of TRPV4 on dural afferents produces headache-related behavior in a preclinical rat model. Cephalalgia 2011, 31, 1595–1600. [Google Scholar] [CrossRef]
- Garrett, F.G.; Hawkins, J.L.; Overmyer, A.E.; Hayden, J.B.; Durham, P.L. Validation of a novel rat-holding device for studying heat- and mechanical-evoked trigeminal nocifensive behavioral responses. J. Orofac. Pain 2012, 26, 337–344. [Google Scholar]
- Wang, S.; Wang, J.; Liu, K.; Bai, W.; Cui, X.; Han, S.; Gao, X.; Zhu, B. Signaling Interaction between Facial and Meningeal Inputs of the Trigeminal System Mediates Peripheral Neurostimulation Analgesia in a Rat Model of Migraine. Neuroscience 2020, 433, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Neubert, J.K.; Rossi, H.L.; Malphurs, W.; Vierck, C.J.; Caudle, R.M. Differentiation between capsaicin-induced allodynia and hyperalgesia using a thermal operant assay. Behav. Brain Res. 2006, 170, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.A.; Hester, J.; Bokrand-Donatelli, Y.; Caudle, R.M.; Neubert, J.K. Adaptation of a novel operant orofacial testing system to characterize both mechanical and thermal pain. Behav. Brain Res. 2011, 217, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Shinoda, M.; Sessle, B.J.; Honda, K.; Imamura, Y.; Hitomi, S.; Tsuboi, Y.; Okada-Ogawa, A.; Iwata, K. Mechanisms Involved in Extraterritorial Facial Pain following Cervical Spinal Nerve Injury in Rats. Mol. Pain 2011, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Shinoda, M.; Kondo, M.; Shimizu, K.; Yonemoto, H.; Otsuki, K.; Akasaka, R.; Furukawa, A.; Iwata, K. Sensitization of TRPV1 and TRPA1 via peripheral mGluR5 signaling contributes to thermal and mechanical hypersensitivity. Pain 2017, 158, 1754–1764. [Google Scholar] [CrossRef]
- Kitagawa, S.; Tang, C.; Unekawa, M.; Kayama, Y.; Nakahara, J.; Shibata, M. Sustained Effects of CGRP Blockade on Cortical Spreading Depolarization-Induced Alterations in Facial Heat Pain Threshold, Light Aversiveness, and Locomotive Activity in the Light Environment. Int. J. Mol. Sci. 2022, 23, 13807. [Google Scholar] [CrossRef]
- Strassman, A.M.; Vos, B.P.; Mineta, Y.; Naderi, S.; Borsook, D.; Burstein, R. Fos-like immunoreactivity in the superficial medullary dorsal horn induced by noxious and innocuous thermal stimulation of facial skin in the rat. J. Neurophysiol. 1993, 70, 1811–1821. [Google Scholar] [CrossRef]
- Bhakta, M.; Vuong, T.; Taura, T.; Wilson, D.S.; Stratton, J.R.; Mackenzie, K.D. Migraine therapeutics differentially modulate the CGRP pathway. Cephalalgia 2021, 41, 499–514. [Google Scholar] [CrossRef]
- Grell, A.-S.; Haanes, K.A.; Johansson, S.E.; Edvinsson, L.; Sams, A. Fremanezumab inhibits vasodilatory effects of CGRP and capsaicin in rat cerebral artery—Potential role in conditions of severe vasoconstriction. Eur. J. Pharmacol. 2019, 864, 172726. [Google Scholar] [CrossRef]
- Dux, M.; Vogler, B.; Kuhn, A.; Mackenzie, K.D.; Stratton, J.; Messlinger, K. The Anti-CGRP Antibody Fremanezumab Lowers CGRP Release from Rat Dura Mater and Meningeal Blood Flow. Cells 2022, 11, 1768. [Google Scholar] [CrossRef]
- Hay, D.L.; Garelja, M.L.; Poyner, D.R.; Walker, C.S. Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR Review 25. Br. J. Pharmacol. 2018, 175, 3–17. [Google Scholar] [CrossRef]
- Zhang, Q.; Han, X.; Wu, H.; Zhang, M.; Hu, G.; Dong, Z.; Yu, S. Dynamic changes in CGRP, PACAP, and PACAP receptors in the trigeminovascular system of a novel repetitive electrical stimulation rat model: Relevant to migraine. Mol. Pain 2019, 15, 1744806918820452. [Google Scholar] [CrossRef]
- Araya, E.I.; Turnes, J.D.M.; Barroso, A.R.; Chichorro, J.G. Contribution of intraganglionic CGRP to migraine-like responses in male and female rats. Cephalalgia 2020, 40, 689–700. [Google Scholar] [CrossRef]
- Caronna, E.; Gallardo, V.J.; Alpuente, A.; Torres-Ferrus, M.; Pozo-Rosich, P. Anti-CGRP monoclonal antibodies in chronic migraine with medication overuse: Real-life effectiveness and predictors of response at 6 months. J. Headache Pain 2021, 22, 120. [Google Scholar] [CrossRef]
- Blumenfeld, A.; Durham, P.L.; Feoktistov, A.; Hay, D.L.; Russo, A.F.; Turner, I. Hypervigilance, Allostatic Load, and Migraine Prevention: Antibodies to CGRP or Receptor. Neurol. Ther. 2021, 10, 469–497. [Google Scholar] [CrossRef]
- Benbow, T.; Ekbatan, M.R.; Wang, G.H.Y.; Teja, F.; Exposto, F.G.; Svensson, P.; Cairns, B.E. Systemic administration of monosodium glutamate induces sexually dimorphic headache- and nausea-like behaviours in rats. Pain 2022, 163, 1838–1853. [Google Scholar] [CrossRef]
- Stucky, N.L.; Gregory, E.; Winter, M.K.; He, Y.-Y.; Hamilton, E.S.; McCarson, K.; Berman, N.E. Sex Differences in Behavior and Expression of CGRP-Related Genes in a Rodent Model of Chronic Migraine. Headache 2011, 51, 674–692. [Google Scholar] [CrossRef]
- Bree, D.; Mackenzie, K.; Stratton, J.; Levy, D. Enhanced post-traumatic headache-like behaviors and diminished contribution of peripheral CGRP in female rats following a mild closed head injury. Cephalalgia 2020, 40, 748–760. [Google Scholar] [CrossRef]
- De Logu, F.; Nassini, R.; Hegron, A.; Landini, L.; Jensen, D.D.; Latorre, R.; Ding, J.; Marini, M.; de Araujo, D.S.M.; Ramírez-Garcia, P.; et al. Schwann cell endosome CGRP signals elicit periorbital mechanical allodynia in mice. Nat. Commun. 2022, 13, 646. [Google Scholar] [CrossRef]
- Majmudar, N.G.; Robson, S.C.; Ford, G.A. Effects of the Menopause, Gender, and Estrogen Replacement Therapy on Vascular Nitric Oxide Activity. J. Clin. Endocrinol. Metab. 2000, 85, 1577–1583. [Google Scholar] [CrossRef]
- Burch, R.; Rizzoli, P.; Loder, E.; Loder, E. The prevalence and impact of migraine and severe headache in the United States: Updated age, sex, and socioeconomic-specific estimates from government health surveys. Headache 2021, 61, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Burstein, R.; Noseda, R.; Borsook, D. Migraine: Multiple Processes, Complex Pathophysiology. J. Neurosci. 2015, 35, 6619–6629. [Google Scholar] [CrossRef] [PubMed]
- Goadsby, P.J. Migraine, Allodynia, Sensitisation and All of That…. Eur. Neurol. 2005, 53 (Suppl. S1), 10–16. [Google Scholar] [CrossRef] [PubMed]
- Silvestro, M.; Tessitore, A.; Orologio, I.; De Micco, R.; Tartaglione, L.; Trojsi, F.; Tedeschi, G.; Russo, A. Galcanezumab effect on “whole pain burden” and multidimensional outcomes in migraine patients with previous unsuccessful treatments: A real-world experience. J. Headache Pain 2022, 23, 69. [Google Scholar] [CrossRef]
- Barbanti, P.; Aurilia, C.; Egeo, G.; Torelli, P.; Proietti, S.; Cevoli, S.; Bonassi, S. For the Italian Migraine Registry Study Group. Late Response to Anti-CGRP Monoclonal Antibodies in Migraine: A Multicenter, Prospective, Observational Study. Neurology 2023. [Google Scholar] [CrossRef]
- Sand, T.; Zhitniy, N.; Nilsen, K.B.; Helde, G.; Hagen, K.; Stovner, L.J. Thermal pain thresholds are decreased in the migraine preattack phase. Eur. J. Neurol. 2008, 15, 1199–1205. [Google Scholar] [CrossRef]
- Kopruszinski, C.M.; Xie, J.Y.; Eyde, N.M.; Remeniuk, B.; Walter, S.; Stratton, J.; Bigal, M.; Chichorro, J.; Dodick, D.; Porreca, F. Prevention of stress- or nitric oxide donor-induced medication overuse headache by a calcitonin gene-related peptide antibody in rodents. Cephalalgia 2017, 37, 560–570. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benedicter, N.; Messlinger, K.; Vogler, B.; Mackenzie, K.D.; Stratton, J.; Friedrich, N.; Dux, M. Semi-Automated Recording of Facial Sensitivity in Rat Demonstrates Antinociceptive Effects of the Anti-CGRP Antibody Fremanezumab. Neurol. Int. 2023, 15, 622-637. https://doi.org/10.3390/neurolint15020039
Benedicter N, Messlinger K, Vogler B, Mackenzie KD, Stratton J, Friedrich N, Dux M. Semi-Automated Recording of Facial Sensitivity in Rat Demonstrates Antinociceptive Effects of the Anti-CGRP Antibody Fremanezumab. Neurology International. 2023; 15(2):622-637. https://doi.org/10.3390/neurolint15020039
Chicago/Turabian StyleBenedicter, Nicola, Karl Messlinger, Birgit Vogler, Kimberly D. Mackenzie, Jennifer Stratton, Nadine Friedrich, and Mária Dux. 2023. "Semi-Automated Recording of Facial Sensitivity in Rat Demonstrates Antinociceptive Effects of the Anti-CGRP Antibody Fremanezumab" Neurology International 15, no. 2: 622-637. https://doi.org/10.3390/neurolint15020039