
Sex and Cross-Sex Testosterone Treatment Alters Gamma-Hydroxybutyrate Acid Toxicokinetics and Toxicodynamics in Rats
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals, Estrus Staging, and Hormone Treatment
2.3. Toxicokinetic Study
2.4. Analysis of GHB in Plasma and Urine
2.5. Quantification of Plasma Testosterone
2.6. Data Analysis
3. Results
3.1. Plasma Testosterone Levels
3.2. Toxicokinetic Analysis
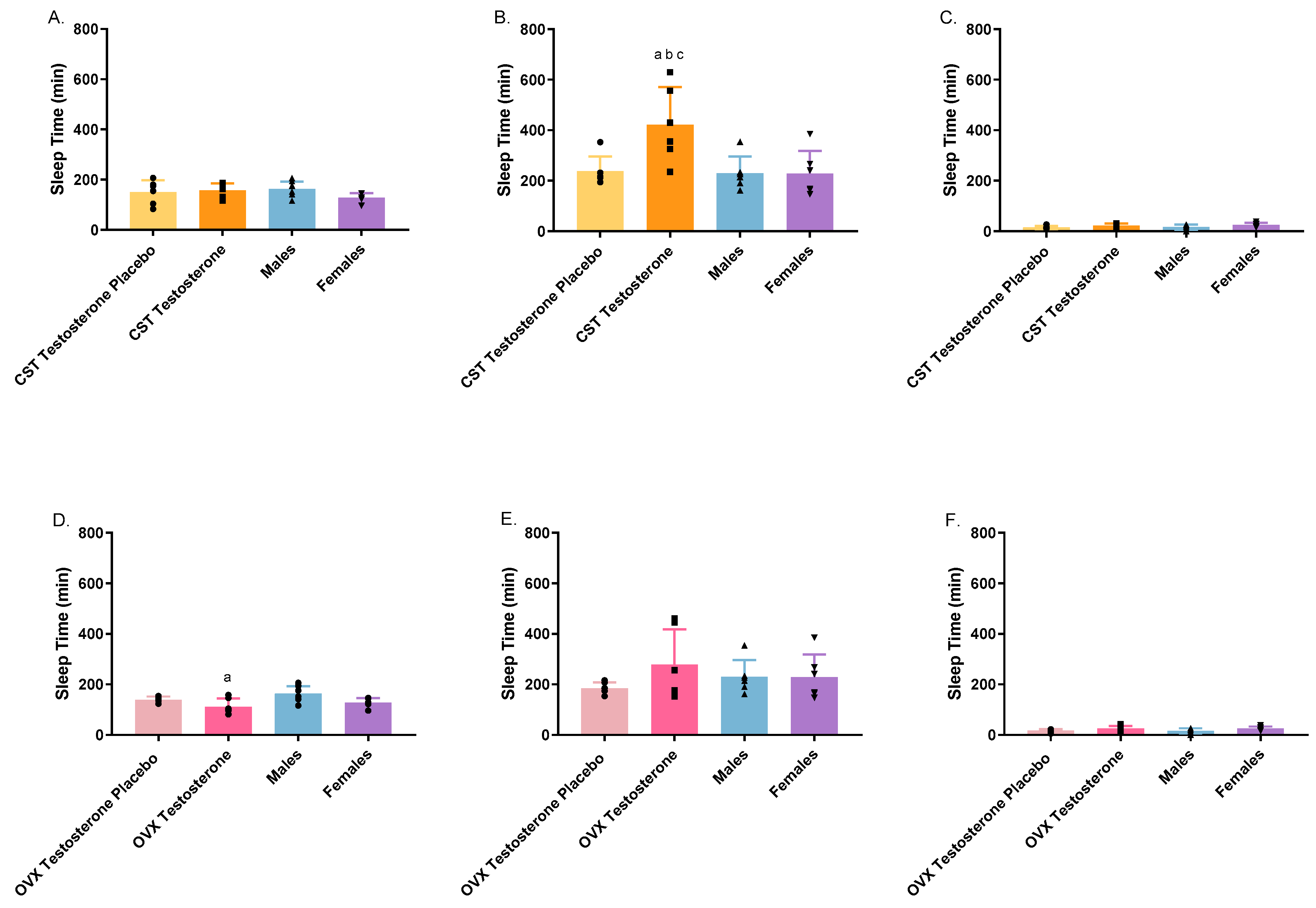
3.3. Toxicodynamic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, C.M. Pharmacologic, Pharmacokinetic, and Clinical Assessment of Illicitly Used γ-Hydroxybutyrate. J. Clin. Pharmacol. 2017, 57, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Felmlee, M.A.; Morse, B.L.; Follman, K.E.; Morris, M.E. The drug of abuse gamma-hydroxybutyric acid exhibits tissue-specific nonlinear distribution. AAPS J. 2018, 20, 21. [Google Scholar] [CrossRef] [PubMed]
- Busardò, F.P.; Jones, A.W. GHB pharmacology and toxicology: Acute intoxication, concentrations in blood and urine in forensic cases and treatment of the withdrawal syndrome. Curr. Neuropharmacol. 2015, 13, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Schep, L.J.; Knudsen, K.; Slaughter, R.J.; Vale, J.A.; Mégarbane, B. The clinical toxicology of gamma-hydroxybutyrate, gamma-butyrolactone and 1, 4-butanediol. Clin. Toxicol. 2012, 50, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Tay, E.; Lo, W.K.W.; Murnion, B. Current insights on the impact of gamma-hydroxybutyrate (GHB) abuse. Subst. Abus. Rehabil. 2022, 13, 13–23. [Google Scholar] [CrossRef]
- Busardò, F.P.; Gottardi, M.; Tini, A.; Minutillo, A.; Sirignano, A.; Marinelli, E.; Zaami, S. Replacing GHB with GBL in recreational settings: A new trend in chemsex. Curr. Drug Metab. 2018, 19, 1080–1085. [Google Scholar] [CrossRef]
- Edmundson, C.; Heinsbroek, E.; Glass, R.; Hope, V.; Mohammed, H.; White, M.; Desai, M. Sexualised drug use in the United Kingdom (UK): A review of the literature. Int. J. Drug Policy 2018, 55, 131–148. [Google Scholar] [CrossRef]
- Dijkstra, B.A.G.; Beurmanjer, H.; Goudriaan, A.; Schellekens, A.; Joosten, E. Unity in diversity: A systematic review on the GHB using population. Int. J. Drug Policy 2021, 94, 103230. [Google Scholar] [CrossRef]
- Wang, Q.; Morris, M.E. The role of monocarboxylate transporter 2 and 4 in the transport of γ-hydroxybutyric acid in mammalian cells. Drug Metab. Dispos. 2007, 35, 1393–1399. [Google Scholar] [CrossRef]
- Morris, M.E.; Hu, K.; Wang, Q. Renal clearance of γ-hydroxybutyric acid in rats: Increasing renal elimination as a detoxification strategy. J. Pharmacol. Exp. Ther. 2005, 313, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Q.; Darling, I.M.; Morris, M.E. Transport of γ-hydroxybutyrate in rat kidney membrane vesicles: Role of monocarboxylate transporters. J. Pharmacol. Exp. Ther. 2006, 318, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Morse, B.L.; Felmlee, M.A.; Morris, M.E. γ-Hydroxybutyrate blood/plasma partitioning: Effect of physiologic pH on transport by monocarboxylate transporters. Drug Metab. Dispos. 2012, 40, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Morris, M.E. The drug of abuse γ-hydroxybutyrate is a substrate for sodium-coupled monocarboxylate transporter (SMCT) 1 (SLC5A8): Characterization of SMCT-mediated uptake and inhibition. Drug Metab. Dispos. 2009, 37, 1404–1410. [Google Scholar] [CrossRef] [PubMed]
- Felmlee, M.A.; Jones, R.S.; Rodriguez-Cruz, V.; Follman, K.E.; Morris, M.E. Monocarboxylate transporters (SLC16): Function, regulation, and role in health and disease. Pharmacol. Rev. 2020, 72, 466–485. [Google Scholar] [CrossRef] [PubMed]
- Gopal, E.; Fei, Y.-J.; Sugawara, M.; Miyauchi, S.; Zhuang, L.; Martin, P.; Smith, S.B.; Prasad, P.D.; Ganapathy, V. Expression of slc5a8 in kidney and its role in Na+-coupled transport of lactate. J. Biol. Chem. 2004, 279, 44522–44532. [Google Scholar] [CrossRef] [PubMed]
- Felmlee, M.A.; Dave, R.A.; Morris, M.E. Mechanistic models describing active renal reabsorption and secretion: A simulation-based study. AAPS J. 2013, 15, 278–287. [Google Scholar] [CrossRef]
- Enoki, T.; Yoshida, Y.; Lally, J.; Hatta, H.; Bonen, A. Testosterone increases lactate transport, monocarboxylate transporter (MCT) 1 and MCT4 in rat skeletal muscle. J. Physiol. 2006, 577, 433–443. [Google Scholar] [CrossRef]
- Cupeiro, R.; González-Lamuño, D.; Amigo, T.; Peinado, A.B.; Ruiz, J.R.; Ortega, F.B.; Benito, P.J. Influence of the MCT1-T1470A polymorphism (rs1049434) on blood lactate accumulation during different circuit weight trainings in men and women. J. Sci. Med. Sport 2012, 15, 541–547. [Google Scholar] [CrossRef]
- Ibarra, M.; Vázquez, M.; Fagiolino, P.; Derendorf, H. Sex related differences on valproic acid pharmacokinetics after oral single dose. J. Pharmacokinet. Pharmacodyn. 2013, 40, 479–486. [Google Scholar] [CrossRef]
- Cao, J.; Ng, M.; Felmlee, M.A. Sex hormones regulate rat hepatic monocarboxylate transporter expression and membrane trafficking. J. Pharm. Pharm. Sci. 2017, 20, 435–444. [Google Scholar] [CrossRef]
- Wei, H.; Cao, J.; Fallert, T.; Yeo, S.; Felmlee, M.A. GHB toxicokinetics and renal monocarboxylate transporter expression are influenced by the estrus cycle in rats. BMC Pharmacol. Toxicol. 2023, 24, 58. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Lee, A.; Zhang, Q.; Felmlee, M.A. Effect of Sex and Cross-Sex Hormone Treatment on Renal Monocarboxylate-Transporter Expression in Rats. Pharmaceutics 2023, 15, 2404. [Google Scholar] [CrossRef] [PubMed]
- Felmlee, M.A.; Morse, B.L.; Morris, M.E. γ-Hydroxybutyric acid: Pharmacokinetics, pharmacodynamics, and toxicology. AAPS J. 2021, 23, 22. [Google Scholar] [CrossRef] [PubMed]
- Brenneisen, R.; ElSohly, M.A.; Murphy, T.P.; Passarelli, J.; Russmann, S.; Salamone, S.J.; Watson, D.E. Pharmacokinetics and excretion of gamma-hydroxybutyrate (GHB) in healthy subjects. J. Anal. Toxicol. 2004, 28, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, X.; Morris, M.E. Effects of l-lactate and d-mannitol on γ-hydroxybutyrate toxicokinetics and toxicodynamics in rats. Drug Metab. Dispos. 2008, 36, 2244–2251. [Google Scholar] [CrossRef]
- Morse, B.L.; Vijay, N.; Morris, M.E. γ-Hydroxybutyrate (GHB)-induced respiratory depression: Combined receptor-transporter inhibition therapy for treatment in GHB overdose. Mol. Pharmacol. 2012, 82, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Ovens, M.J.; Davies, A.J.; Wilson, M.C.; Murray, C.M.; Halestrap, A.P. AR-C155858 is a potent inhibitor of monocarboxylate transporters MCT1 and MCT2 that binds to an intracellular site involving transmembrane helices 7–10. Biochem. J. 2010, 425, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Vijay, N.; Morse, B.L.; Morris, M.E. A novel monocarboxylate transporter inhibitor as a potential treatment strategy for γ-hydroxybutyric acid overdose. Pharm. Res. 2015, 32, 1894–1906. [Google Scholar] [CrossRef]
- Follman, K.E.; Morris, M.E. Treatment of γ-hydroxybutyric acid and γ-butyrolactone overdose with two potent monocarboxylate transporter 1 inhibitors, AZD3965 and AR-C155858. J. Pharmacol. Exp. Ther. 2019, 370, 84–91. [Google Scholar] [CrossRef]
- Cora, M.C.; Kooistra, L.; Travlos, G. Vaginal cytology of the laboratory rat and mouse: Review and criteria for the staging of the estrous cycle using stained vaginal smears. Toxicol. Pathol. 2015, 43, 776–793. [Google Scholar] [CrossRef]
- McLean, A.C.; Valenzuela, N.; Fai, S.; Bennett, S.A.L. Performing vaginal lavage, crystal violet staining, and vaginal cytological evaluation for mouse estrous cycle staging identification. JoVE (J. Vis. Exp.) 2012, 67, e4389. [Google Scholar]
- Felmlee, M.A.; Roiko, S.A.; Morse, B.L.; Morris, M.E. Concentration-effect relationships for the drug of abuse γ-hydroxybutyric acid. J. Pharmacol. Exp. Ther. 2010, 333, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Damassa, D.A.; Smith, E.R.; Tennent, B.; Davidson, J.M. The relationship between circulating testosterone levels and male sexual behavior in rats. Horm. Behav. 1977, 8, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Shulman, L.M.; Spritzer, M.D. Changes in the sexual behavior and testosterone levels of male rats in response to daily interactions with estrus females. Physiol. Behav. 2014, 133, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.M.; Moreira, A.C. Daily variation of plasma testosterone, androstenedione, and corticosterone in rats under food restriction. Horm. Behav. 1997, 31, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Mock, E.J.; Norton, H.W.; Frankel, A.I. Daily rhythmicity of serum testosterone concentration in the male laboratory rat. Endocrinology 1978, 103, 1111–1121. [Google Scholar] [CrossRef]
- Falvo, R.; Kaltenbach, C.; Pancoe, W. Determination of testosterone concentration in the plasma of normal and androgen-sterilized female rats, using a competitive protein binding technique. Neuroendocrinology 1972, 10, 229–234. [Google Scholar] [CrossRef]
- Rush, M.E.; Blake, C.A. Serum testosterone concentrations during the 4-day estrous cycle in normal and adrenalectomized rats. Proc. Soc. Exp. Biol. Med. 1982, 169, 216–221. [Google Scholar] [CrossRef]
- Carai, M.A.; Colombo, G.; Brunetti, G.; Melis, S.; Serra, S.; Vacca, G.; Mastinu, S.; Pistuddi, A.M.; Solinas, C.; Cignarella, G.; et al. Role of GABAB receptors in the sedative/hypnotic effect of γ-hydroxybutyric acid. Eur. J. Pharmacol. 2001, 428, 315–321. [Google Scholar] [CrossRef]
- Vijay, N.; Morris, M.E. Role of monocarboxylate transporters in drug delivery to the brain. Curr. Pharm. Des. 2014, 20, 1487–1498. [Google Scholar] [CrossRef]
- Bhattacharya, I.; Boje, K.M. GHB (γ-hydroxybutyrate) carrier-mediated transport across the blood-brain barrier. J. Pharmacol. Exp. Ther. 2004, 311, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Kwatra, N.V.; Morris, M.E. Toxicokinetic/toxicodynamic interaction studies in rats between the drugs of abuse γ-Hydroxybutyric acid and ketamine and treatment strategies for overdose. Pharmaceutics 2021, 13, 741. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Cruz, V.; Morris, M.E. γ-Hydroxybutyric Acid–Ethanol Drug-Drug Interaction: Reversal of Toxicity with Monocarboxylate Transporter 1 Inhibitors. J. Pharmacol. Exp. Ther. 2021, 378, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Alluri, R.V.; Li, R.; Varma, M.V. Transporter–enzyme interplay and the hepatic drug clearance: What have we learned so far? Expert Opin. Drug Metab. Toxicol. 2020, 16, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Longcope, C.; Kato, T.; Horton, R. Conversion of blood androgens to estrogens in normal adult men and women. J. Clin. Investig. 1969, 48, 2191–2201. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.R. Aromatization of androgens in women: Current concepts and findings. Fertil. Steril. 2002, 77, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Lakshman, K.M.; Kaplan, B.; Travison, T.G.; Basaria, S.; Knapp, P.E.; Singh, A.B.; LaValley, M.P.; Mazer, N.A.; Bhasin, S. The effects of injected testosterone dose and age on the conversion of testosterone to estradiol and dihydrotestosterone in young and older men. J. Clin. Endocrinol. Metab. 2010, 95, 3955–3964. [Google Scholar] [CrossRef]
- Chan, K.J.; Jolly, D.; Liang, J.J.; Weinand, J.D.; Safer, J.D. Estrogen levels do not rise with testosterone treatment for transgender men. Endocr. Pract. 2018, 24, 329–333. [Google Scholar] [CrossRef]
- Snyder, E.M.; Small, C.L.; Li, Y.; Griswold, M.D. Regulation of gene expression by estrogen and testosterone in the proximal mouse reproductive tract. Biol. Reprod. 2009, 81, 707–716. [Google Scholar] [CrossRef]
- Sever, R.; Glass, C.K. Signaling by nuclear receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a016709. [Google Scholar] [CrossRef]
- Weikum, E.R.; Liu, X.; Ortlund, E.A. The nuclear receptor superfamily: A structural perspective. Protein Sci. 2018, 27, 1876–1892. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma Hormone Level | Testosterone (ng/mL) | Testosterone Non-Detected |
|---|---|---|
| OVX Testosterone Placebo | -- | 18 out of 18 |
| OVX Testosterone | 6.220 ± 2.627 | 0 out of 18 |
| CST Testosterone Placebo | -- | 18 out of 18 |
| CST Testosterone | 5.440 ± 1.955 | 0 out of 17 |
| Females (estrus) | 0.481 | 16 out of 17 |
| Males | 0.786 ± 0.462 | 2 out of 21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Wei, H.; Lee, A.; Felmlee, M.A. Sex and Cross-Sex Testosterone Treatment Alters Gamma-Hydroxybutyrate Acid Toxicokinetics and Toxicodynamics in Rats. Pharmaceutics 2024, 16, 143. https://doi.org/10.3390/pharmaceutics16010143
Zhang Q, Wei H, Lee A, Felmlee MA. Sex and Cross-Sex Testosterone Treatment Alters Gamma-Hydroxybutyrate Acid Toxicokinetics and Toxicodynamics in Rats. Pharmaceutics. 2024; 16(1):143. https://doi.org/10.3390/pharmaceutics16010143
Chicago/Turabian StyleZhang, Qing, Hao Wei, Annie Lee, and Melanie A. Felmlee. 2024. "Sex and Cross-Sex Testosterone Treatment Alters Gamma-Hydroxybutyrate Acid Toxicokinetics and Toxicodynamics in Rats" Pharmaceutics 16, no. 1: 143. https://doi.org/10.3390/pharmaceutics16010143