

The Impact of Pigment-Epithelium-Derived Factor on MCF-7 Cell Metabolism in the Context of Glycaemic Condition



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Cell Cultivation
2.3. Sample Preparation for Metabolomics Analysis
2.4. Metabolome Profiling Using GC/Q-MS
2.5. Data Processing and Statistical Analysis
3. Results
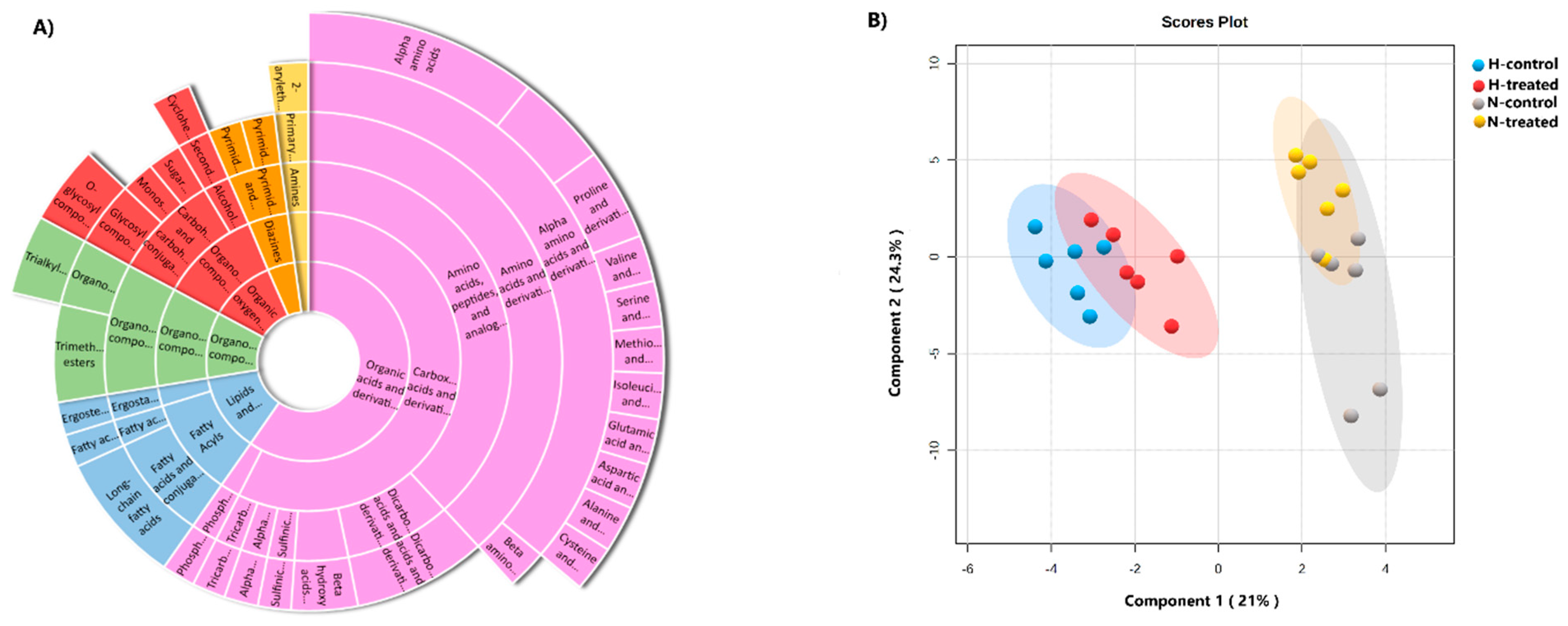
3.1. Metabolic Profile among the Groups
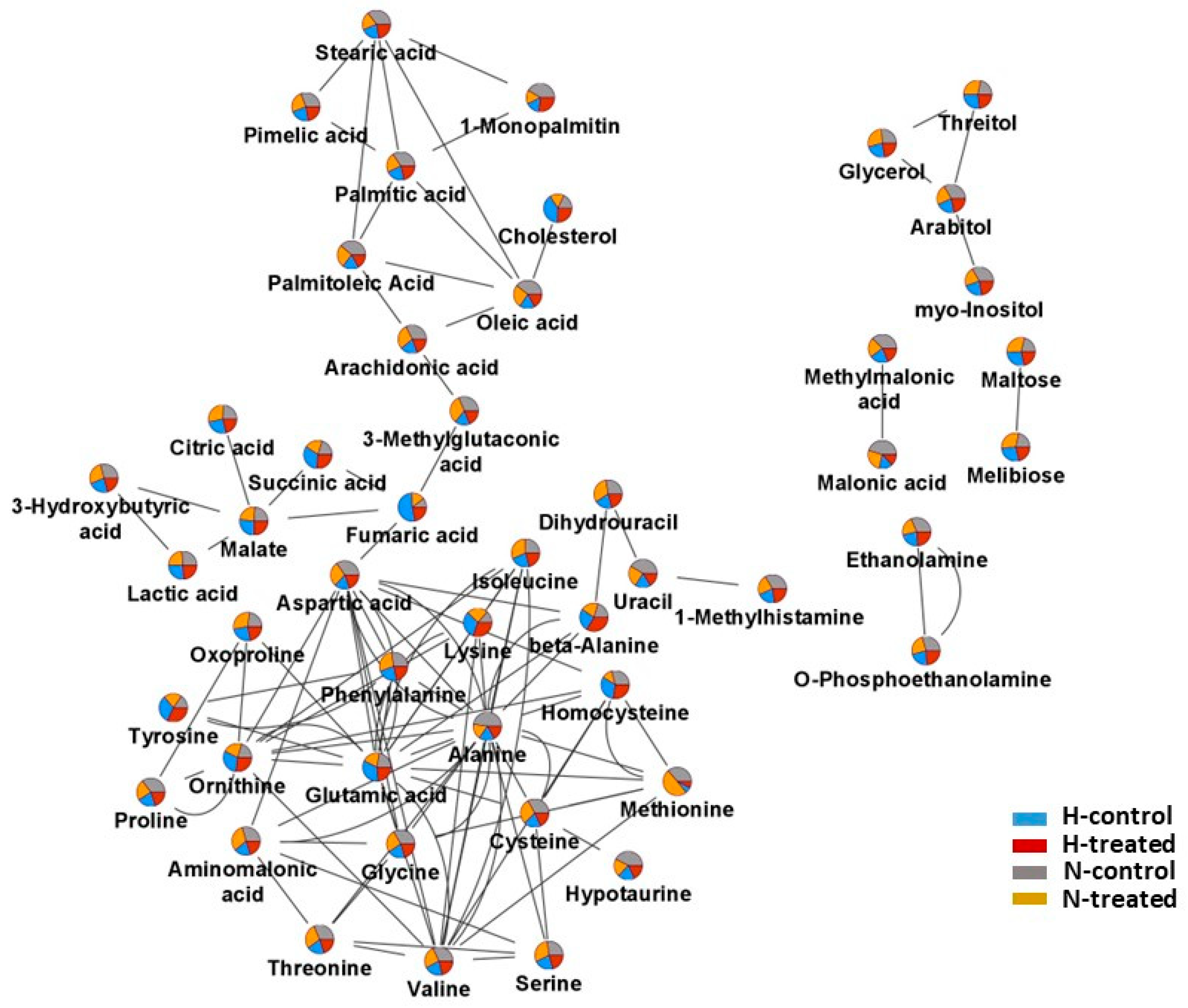
3.2. Response of MCF-7 Metabolomes to PEDF under Glycaemic Loading
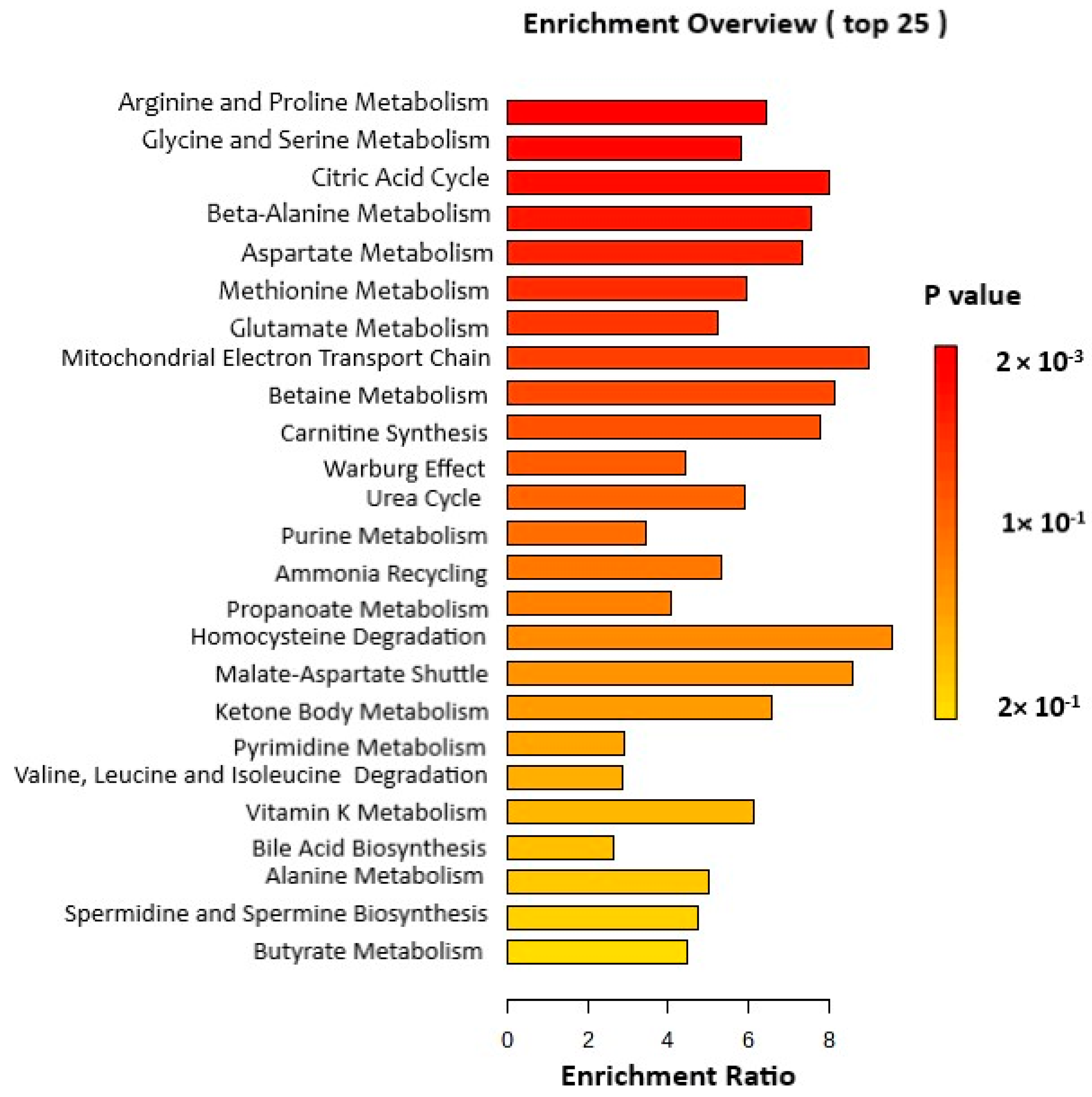
3.3. Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Network. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arruebo, M.; Vilaboa, N.; Sáez-Gutierrez, B.; Lambea, J.; Tres, A.; Valladares, M.; González-Fernández, Á. Assessment of the evolution of cancer treatment therapies. Cancers 2011, 3, 3279–3330. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, N.N.; Thompson, C.B. The emerging hallmarks of cancer metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of cancer metabolism. Sci. Adv. 2016, 2, e1600200. [Google Scholar] [CrossRef] [Green Version]
- Cantor, J.R.; Sabatini, D.M. Cancer Cell Metabolism: One Hallmark, Many FacesCancer Cell Metabolism: One Hallmark, Many Faces. Cancer Discov. 2012, 2, 881–898. [Google Scholar] [CrossRef] [Green Version]
- Vigneri, P.; Frasca, F.; Sciacca, L.; Pandini, G.; Vigneri, R. Diabetes and cancer. Endocr.-Relat. Cancer 2009, 16, 1103–1123. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Zheng, Q.; Meng, X. Hyperglycemia and chemoresistance in breast cancer: From cellular mechanisms to treatment response. Front. Oncol. 2021, 11, 628359. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Sang, H.; Zhou, Y.; Shang, C.; Wang, Y.; Zhu, H. Effects of hyperglycemia on the progression of tumor diseases. J. Exp. Clin. Cancer Res. 2019, 38, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, S.P.; Sagasti, A.; Spinella, P.; Notario, V. Pigment Epithelium-derived Factor Behaves Like a Noninhibitory Serpin: Neurotrophic Activity Does Not Require The Serpin Reactive Loop (∗). J. Biol. Chem. 1995, 270, 25992–25999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abooshahab, R.; Al-Salami, H.; Dass, C.R. The increasing role of pigment epithelium-derived factor in metastasis: From biological importance to a promising target. Biochem. Pharmacol. 2021, 193, 114787. [Google Scholar] [CrossRef]
- Abooshahab, R.; Dass, C.R. The biological relevance of pigment epithelium-derived factor on the path from aging to age-related disease. Mech. Ageing Dev. 2021, 196, 111478. [Google Scholar] [CrossRef] [PubMed]
- Bernard, A.; Gao-Li, J.; Franco, C.A.; Bouceba, T.; Huet, A.; Li, Z. Laminin Receptor Involvement in the Anti-angiogenic Activity of Pigment Epithelium-derived Factor. J. Biol. Chem. 2009, 284, 10480–10490. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, E.; Aymerich, M.S.; Becerra, S.P. Binding of pigment epithelium-derived factor (PEDF) to retinoblastoma cells and cerebellar granule neurons: Evidence for a PEDF receptor. J. Biol. Chem. 1999, 274, 31605–31612. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.T.; Lin, C.C.; Tsai, M.C.; Chen, K.D.; Chiu, K.W. Pigment epithelium-derived factor in lipid metabolic disorders. Biomed. J. 2018, 41, 102–108. [Google Scholar] [CrossRef]
- Carnagarin, R.; Dharmarajan, A.M.; Dass, C.R. PEDF-induced alteration of metabolism leading to insulin resistance. Mol. Cell. Endocrinol. 2015, 401, 98–104. [Google Scholar] [CrossRef]
- Zhou, D.; Cheng, S.Q.; Ji, H.F.; Wang, J.S.; Xu, H.T.; Zhang, G.Q.; Pang, D. Evaluation of protein pigment epithelium-derived factor (PEDF) and microvessel density (MVD) as prognostic indicators in breast cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 1719–1727. [Google Scholar] [CrossRef]
- Danzi, F.; Pacchiana, R.; Mafficini, A.; Scupoli, M.T.; Scarpa, A.; Donadelli, M.; Fiore, A. To metabolomics and beyond: A technological portfolio to investigate cancer metabolism. Signal Transduct. Target. Ther. 2023, 8, 137. [Google Scholar] [CrossRef]
- Ribbenstedt, A.; Ziarrusta, H.; Benskin, J.P. Development, characterization and comparisons of targeted and non-targeted metabolomics methods. PLoS ONE 2018, 13, e0207082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abooshahab, R.; Hooshmand, K.; Luna, G.; Al-Salami, H.; Dass, C.R. Metabolomics Profiling Reveals the Role of PEDF in Triple-Negative Breast Cancer Cell MDA-MB-231 under Glycaemic Loading. Pharmaceutics 2023, 15, 543. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.V.; Valnickova, Z.; Enghild, J.J. Pigment-epithelium-derived factor (PEDF) occurs at a physiologically relevant concentration in human blood: Purification and characterization. Biochem. J. 2003, 374, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djoumbou Feunang, Y.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated chemical classification with a comprehensive, computable taxonomy. J. Cheminform. 2016, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Zhong, M.; Xiong, Y.; Gao, Z.; Wu, Z.; Liu, Y.; Hong, X. Emerging roles of nucleotide metabolism in cancer development: Progress and prospect. Aging 2021, 13, 13349. [Google Scholar] [CrossRef]
- Cao, D.; Pizzorno, G. Uridine Phosphorylase. Drugs Today 2004, 40, 431–443. [Google Scholar]
- Matchett, E.C.; Ambrose, E.C.; Kornbluth, J. Characterization of uridine-cytidine kinase like-1 nucleoside kinase activity and its role in tumor growth. Biochem. J. 2022, 479, 1149–1164. [Google Scholar] [CrossRef]
- Li, J.; Huang, S.; Chen, J.; Yang, Z.; Fei, X.; Zheng, M.; Ji, C.; Xie, Y.; Mao, Y. Identification and characterization of human uracil phosphoribosyltransferase (UPRTase). J. Hum. Genet. 2007, 52, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.H.; Pittman, D.L.; Wyatt, M.D. Uracil in DNA: Consequences for carcinogenesis and chemotherapy. Biochem. Pharmacol. 2008, 76, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of amino acids in cancer. Front. Cell Dev. Biol. 2021, 8, 603837. [Google Scholar] [CrossRef] [PubMed]
- Razak, M.A. Multifarious beneficial effect of nonessential amino acid, glycine: A review. Oxid. Med. Cell. Longev. 2017, 2017, 1716701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Glycine suppresses AGE/RAGE signaling pathway and subsequent oxidative stress by restoring Glo1 function in the aorta of diabetic rats and in HUVECs. Oxid. Med. Cell. Longev. 2019, 2019, 4628962. [Google Scholar]
- Sun, K.; Wu, Z.; Ji, Y.; Wu, G. Glycine regulates protein turnover by activating protein kinase B/mammalian target of rapamycin and by inhibiting MuRF1 and atrogin-1 gene expression in C2C12 myoblasts. J. Nutr. 2016, 146, 2461–2467. [Google Scholar] [CrossRef] [Green Version]
- Chiang, E.P.I.; Wang, Y.C.; Chen, W.W.; Tang, F.Y. Effects of insulin and glucose on cellular metabolic fluxes in homocysteine transsulfuration, remethylation, S-adenosylmethionine synthesis, and global deoxyribonucleic acid methylation. J. Clin. Endocrinol. Metab. 2009, 94, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Sbodio, J.I.; Snyder, S.H.; Paul, B.D. Regulators of the transsulfuration pathway. Br. J. Pharmacol. 2019, 176, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Steegborn, C.; Clausen, T.; Sondermann, P.; Jacob, U.; Worbs, M.; Marinkovic, S.; Huber, R.; Wahl, M.C. Kinetics and inhibition of recombinant human cystathionine γ-lyase: Toward the rational control of transsulfuration. J. Biol. Chem. 1999, 274, 12675–12684. [Google Scholar] [CrossRef] [Green Version]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [Green Version]
- Caruso, J.; Charles, J.; Unruh, K.; Giebel, R.; Learmonth, L.; Potter, W. Ergogenic effects of β-alanine and carnosine: Proposed future research to quantify their efficacy. Nutrients 2012, 4, 585–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derave, W.; Everaert, I.; Beeckman, S.; Baguet, A. Muscle carnosine metabolism and β-alanine supplementation in relation to exercise and training. Sports Med. 2010, 40, 247–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiedje, K.E.; Stevens, K.; Barnes, S.; Weaver, D.F. β-Alanine as a small molecule neurotransmitter. Neurochem. Int. 2010, 57, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.D.; Sale, C.; Garner, A.C.; Hipkiss, A.R. Anti-cancer actions of carnosine and the restoration of normal cellular homeostasis. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2021, 1868, 119117. [Google Scholar] [CrossRef] [PubMed]
- Billacura, M.P.; Cripps, M.J.; Hanna, K.; Sale, C.; Turner, M.D. β-alanine scavenging of free radicals protects mitochondrial function and enhances both insulin secretion and glucose uptake in cells under metabolic stress. Adv. Redox Res. 2022, 6, 100050. [Google Scholar] [CrossRef]
- Prakash, M.D.; Fraser, S.; Boer, J.C.; Plebanski, M.; de Courten, B. Anti-cancer effects of carnosine—A dipeptide molecule. Molecules 2021, 26, 1644. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.; Son, H.; Baek, J.-H. Tricarboxylic acid (Tca) cycle intermediates: Regulators of immune responses. Life 2021, 11, 69. [Google Scholar] [CrossRef]
- Martínez-Reyes, I.; Chandel, N.S. Mitochondrial TCA cycle metabolites control physiology and disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2–Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [Green Version]
- Selak, M.A.; Armour, S.M.; MacKenzie, E.D.; Boulahbel, H.; Watson, D.G.; Mansfield, K.D.; Pan, Y.; Simon, M.C.; Thompson, C.B.; Gottlieb, E. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-α prolyl hydroxylase. Cancer Cell 2005, 7, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Mansouri, S.; Shahriari, A.; Kalantar, H.; Moini Zanjani, T.; Haghi Karamallah, M. Role of malate dehydrogenase in facilitating lactate dehydrogenase to support the glycolysis pathway in tumors. Biomed. Rep. 2017, 6, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Hanse, E.A.; Ruan, C.; Kachman, M.; Wang, D.; Lowman, X.H.; Kelekar, A. Cytosolic malate dehydrogenase activity helps support glycolysis in actively proliferating cells and cancer. Oncogene 2017, 36, 3915–3924. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.T.; Kreutzberger, A.J.; Lee, J.; Kiessling, V.; Tamm, L.K. The role of cholesterol in membrane fusion. Chem. Phys. Lipids 2016, 199, 136–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, M.; Yarrarapu, S.N.S.; Dimri, M. Biochemistry, Cholesterol; StatPearls Publishing LLC: St. Petersburg, FL, USA, 2018. [Google Scholar]
- Ding, X.; Zhang, W.; Li, S.; Yang, H. The role of cholesterol metabolism in cancer. Am. J. Cancer Res. 2019, 9, 219. [Google Scholar] [PubMed]
- Borg, M.L.; Andrews, Z.B.; Duh, E.J.; Zechner, R.; Meikle, P.J.; Watt, M.J. Pigment epithelium–derived factor regulates lipid metabolism via adipose triglyceride lipase. Diabetes 2011, 60, 1458–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubuis, S.; Baenke, F.; Scherbichler, N.; Alexander, L.T.; Schulze, A.; Zamboni, N. Metabotypes of breast cancer cell lines revealed by non-targeted metabolomics. Metab. Eng. 2017, 43, 173–186. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | VIP | One-Way ANOVA | Multiple Comparisons Tukey HSD (a p-Value) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| a p-Value | FDR | HT vs. HC | NC vs. HC | NT vs. HC | NC vs. HT | NT vs. HT | NC vs. NT | ||
| Uracil | 1.0007 | 1.07 × 10−7 | 5.14 × 10−6 | 0.982 | 0.000 | 0.525 | 0.000 | 0.325 | 0.000 |
| beta-Alanine | 1.587 | 1.39 × 10−6 | 3.33 × 10−5 | 0.012 | 0.012 | 0.011 | 0.000 | 0.000 | 1.000 |
| Methionine | 2.1201 | 2.56 × 10−5 | 0.000409 | 0.999 | 0.011 | 0.000 | 0.015 | 0.000 | 0.188 |
| Glycine | 0.97332 | 0.000313 | 0.003589 | 0.851 | 0.000 | 0.573 | 0.002 | 0.959 | 0.007 |
| Succinic acid | 1.643 | 0.000448 | 0.003589 | 0.008 | 0.000 | 0.003 | 0.584 | 0.970 | 0.836 |
| Cholesterol | 1.8464 | 0.000449 | 0.003589 | 0.014 | 0.001 | 0.001 | 0.715 | 0.568 | 0.995 |
| Homocysteine | 1.9154 | 0.000654 | 0.004486 | 0.946 | 0.246 | 0.001 | 0.479 | 0.002 | 0.059 |
| Methylmalonic acid | 1.5986 | 0.002992 | 0.015957 | 0.966 | 0.010 | 0.833 | 0.003 | 0.981 | 0.001 |
| Oleic acid | 1.0777 | 0.006105 | 0.041381 | 0.994 | 0.004 | 0.490 | 0.007 | 0.644 | 0.089 |
| Fumaric acid | 1.4726 | 0.004588 | 0.022023 | 0.037 | 0.005 | 0.019 | 0.793 | 0.989 | 0.929 |
| Threonine | 1.4385 | 0.005339 | 0.023298 | 0.931 | 0.007 | 0.104 | 0.027 | 0.290 | 0.596 |
| Malate | 1.4995 | 0.010921 | 0.040484 | 0.999 | 0.036 | 0.084 | 0.049 | 0.112 | 0.974 |
| Aminomalonic acid | 1.7133 | 0.010964 | 0.040484 | 0.631 | 0.052 | 0.013 | 0.414 | 0.148 | 0.911 |
| Palmitoleic Acid | 0.96085 | 0.012301 | 0.042175 | 1.000 | 0.019 | 0.675 | 0.020 | 0.685 | 0.180 |
| Aspartic acid | 1.2957 | 0.014993 | 0.047976 | 0.673 | 0.011 | 0.168 | 0.116 | 0.737 | 0.548 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abooshahab, R.; Hooshmand, K.; Salami, H.-A.; Dass, C.R. The Impact of Pigment-Epithelium-Derived Factor on MCF-7 Cell Metabolism in the Context of Glycaemic Condition. Pharmaceutics 2023, 15, 2140. https://doi.org/10.3390/pharmaceutics15082140
Abooshahab R, Hooshmand K, Salami H-A, Dass CR. The Impact of Pigment-Epithelium-Derived Factor on MCF-7 Cell Metabolism in the Context of Glycaemic Condition. Pharmaceutics. 2023; 15(8):2140. https://doi.org/10.3390/pharmaceutics15082140
Chicago/Turabian StyleAbooshahab, Raziyeh, Kourosh Hooshmand, Hani-Al Salami, and Crispin R. Dass. 2023. "The Impact of Pigment-Epithelium-Derived Factor on MCF-7 Cell Metabolism in the Context of Glycaemic Condition" Pharmaceutics 15, no. 8: 2140. https://doi.org/10.3390/pharmaceutics15082140