
Pigment Epithelium-Derived Factor: Inhibition of Phosphorylation of Insulin Receptor (IR)/IR Substrate (IRS), Osteogeneration from Adipocytes, and Increased Levels Due to Doxorubicin Exposure
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Ethics
2.3. Mesenchymal Stem Cell (MSC) Isolation and Culture
2.4. Viability Assay
2.5. Immunoblotting
2.6. Fluorescence Immunocytochemistry
2.7. Von Kossa Staining
2.8. Gelfoam Binding and Release
2.9. Gelfoam Implantation Study
2.10. PEDF Detection in MDA-MB-231 Cells Exposed to Doxorubicin (Dox)
2.11. PEDF Detection in Mouse Cardiac Tissue
2.12. Measurement of ROS and GSH
2.13. Statistical Analysis
3. Results
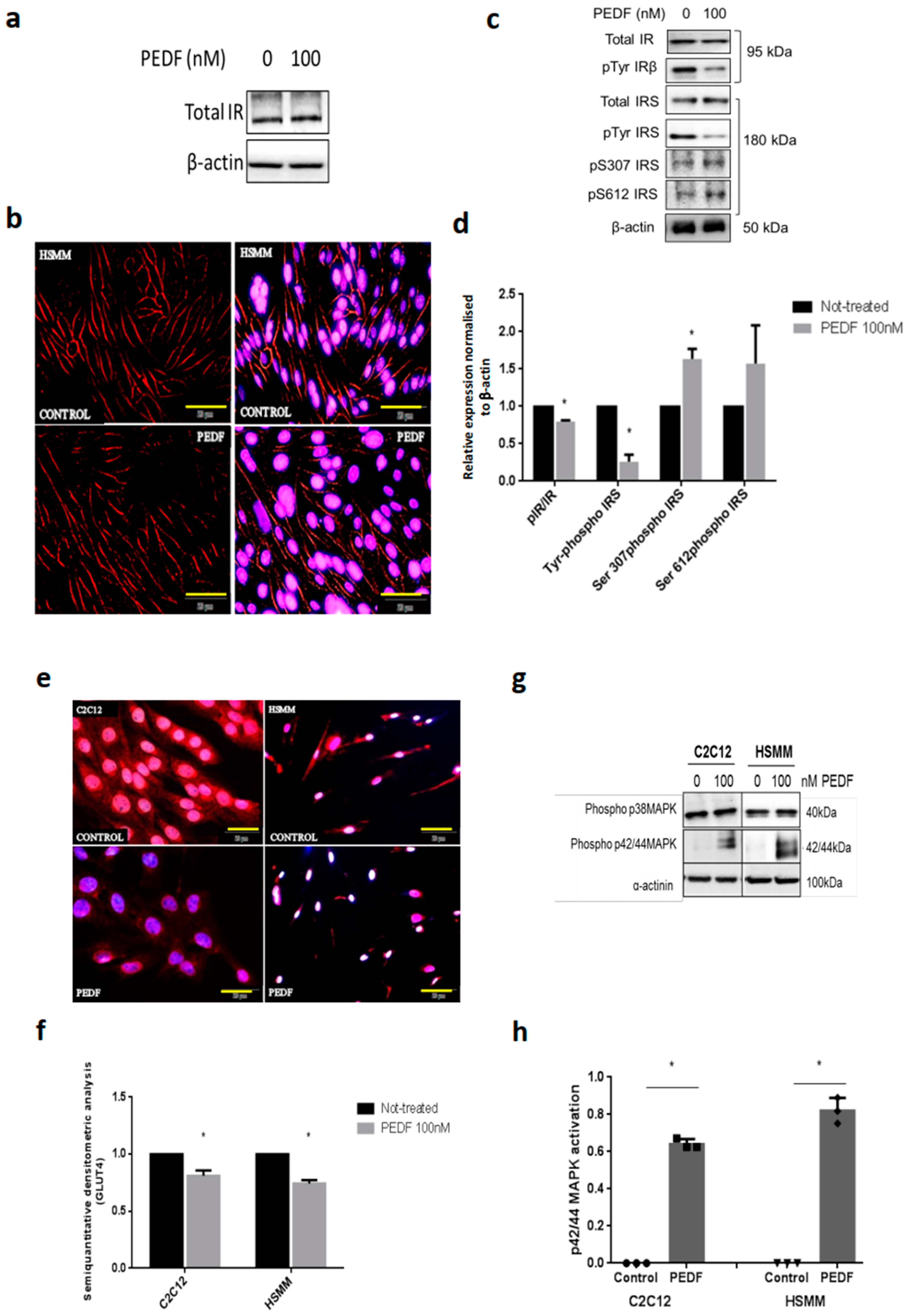
3.1. PEDF Impairs Metabolic Signalling via IRS-1 and Constitutively Activates p42/44 MAPK/Erk Kinase
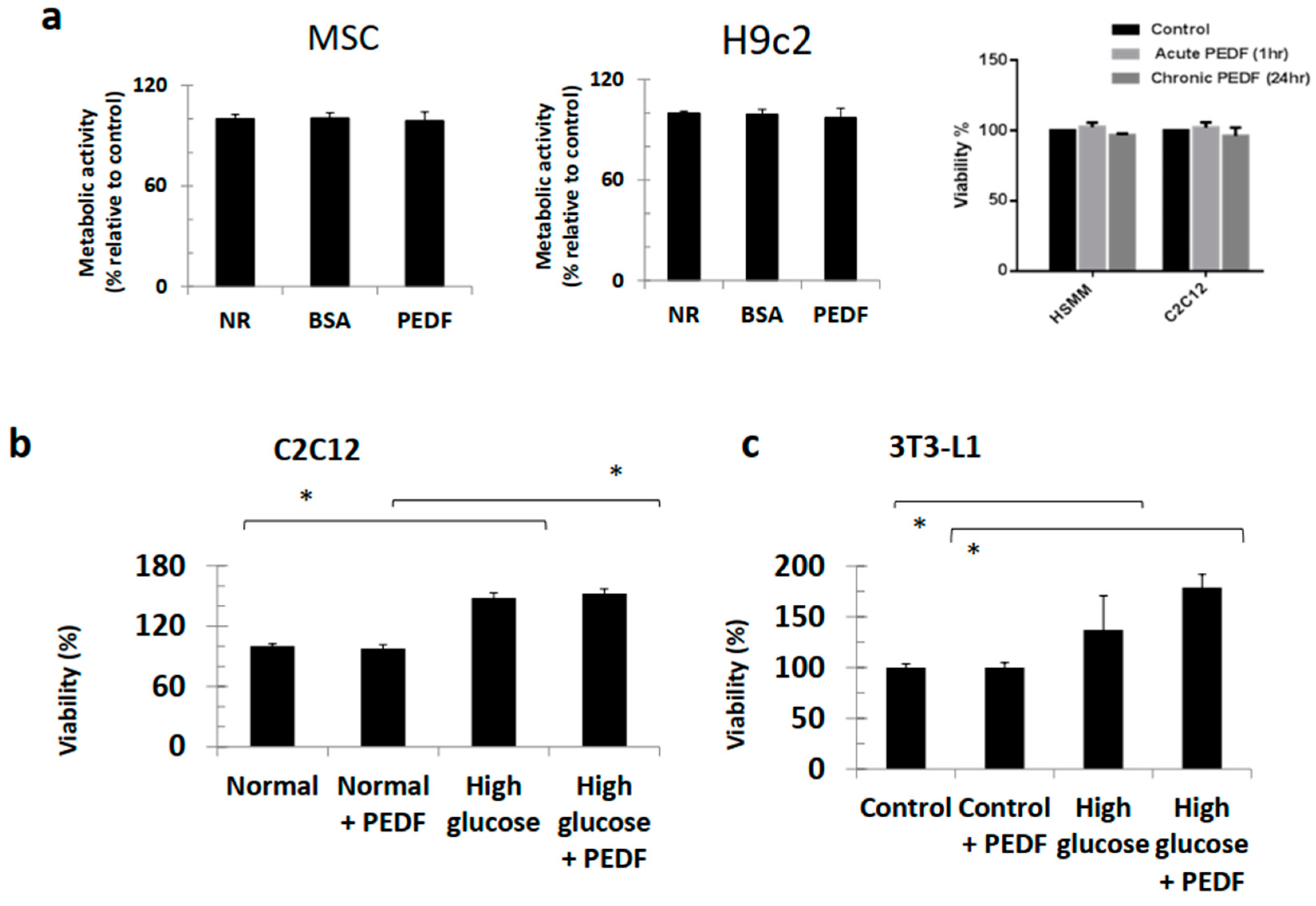
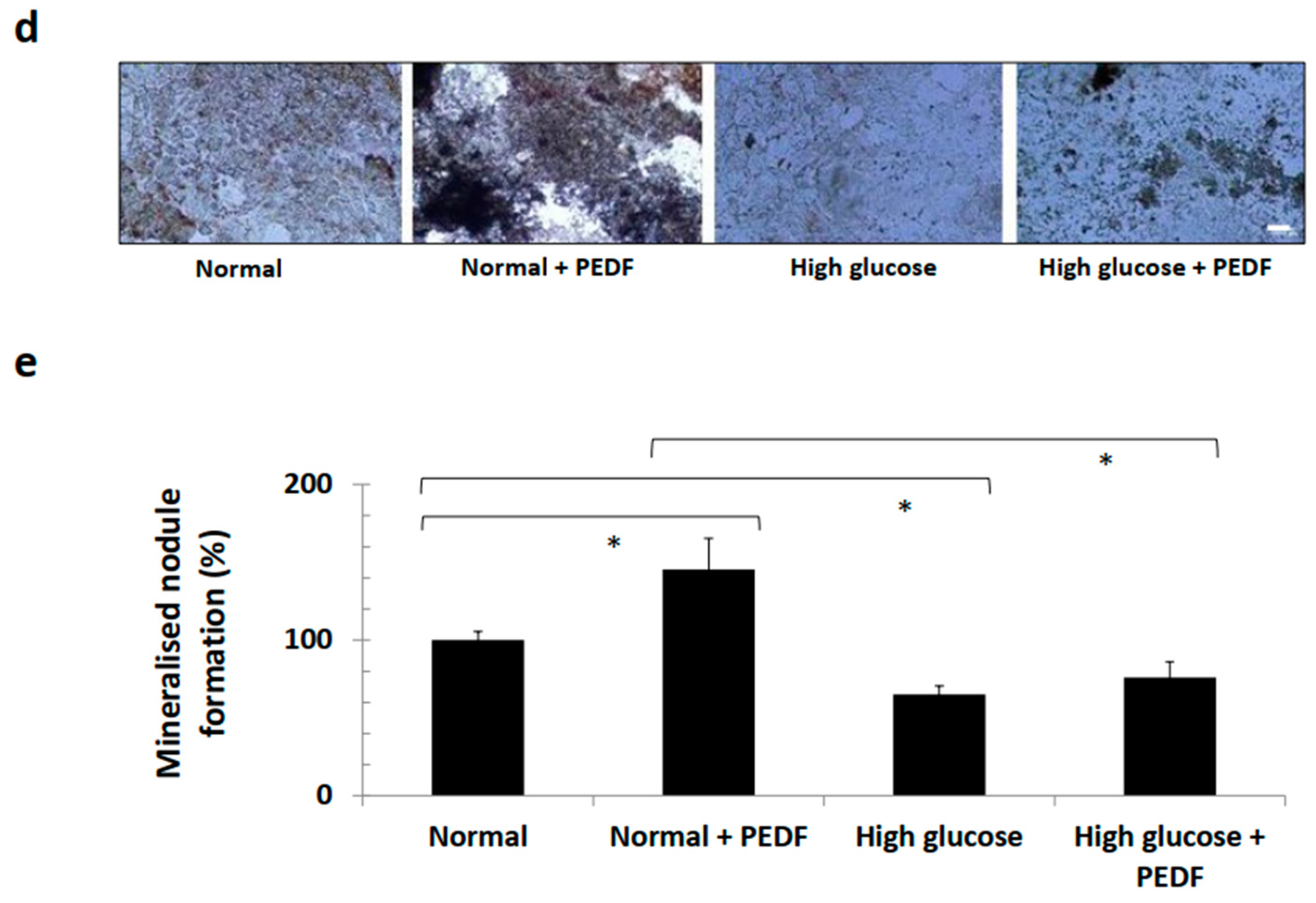
3.2. PEDF Did Not Perturb Viability of Various Cell Lines and Continued to Induce Bone Formation under Hyperglycaemic Conditions
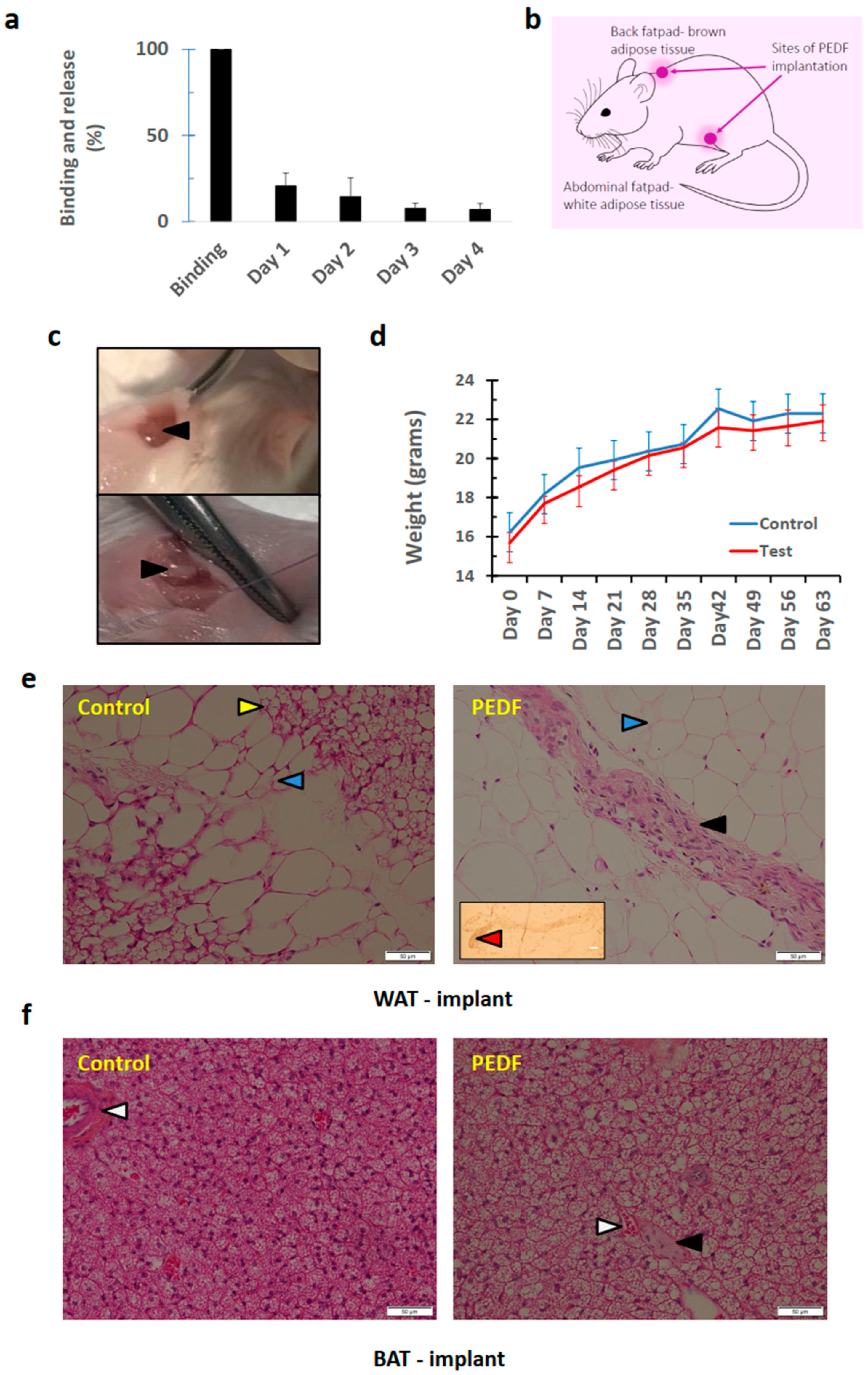
3.3. PEDF Implanted in Adipose Tissue (Brown and White) can Induce De Novo Osteogeneration, with Lack of Harmful Effects to Surrounding Tissues
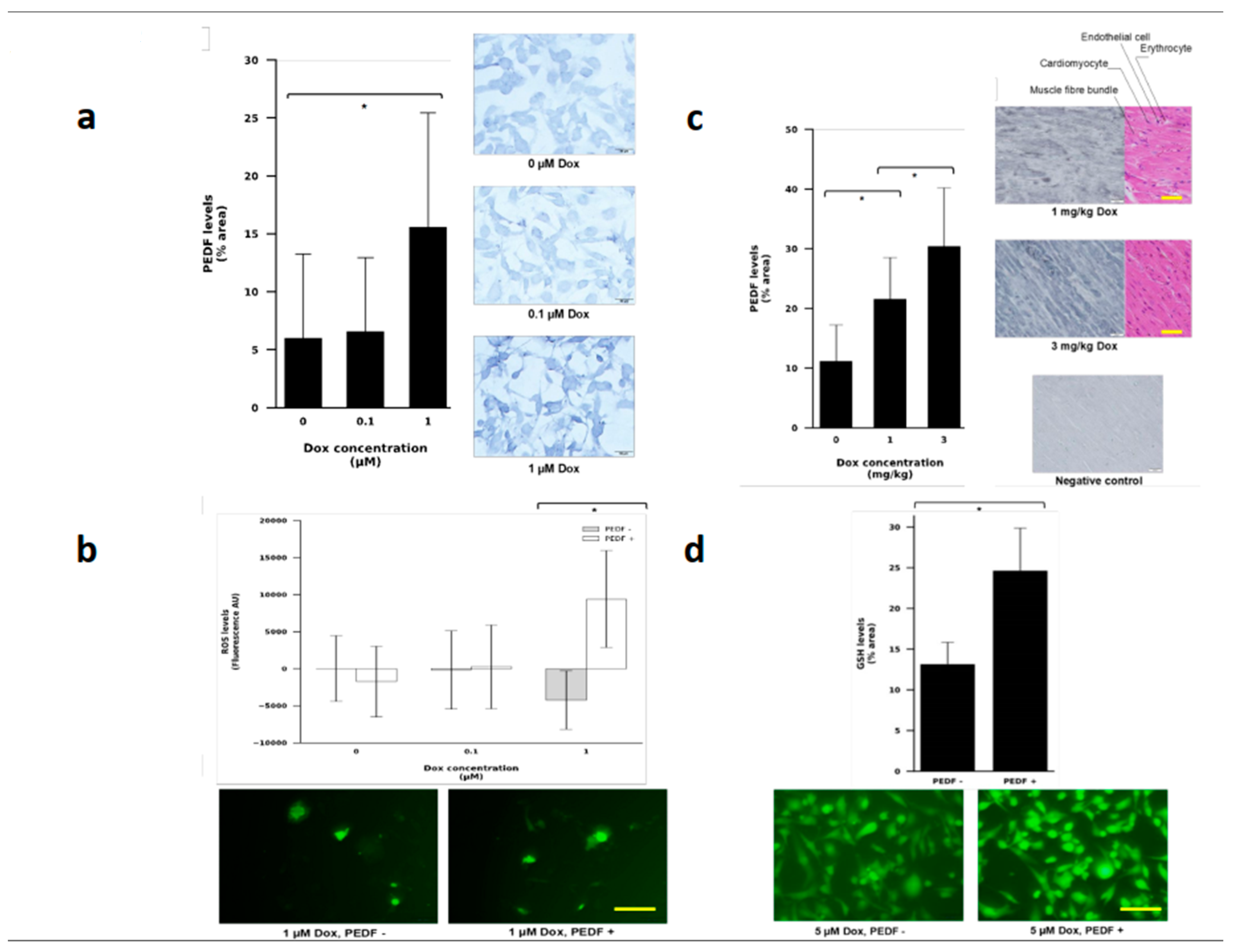
3.4. PEDF Levels Increased in Human Breast Cancer Cell Line Exposed to Doxorubicin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crowe, S.; Wu, L.E.; Economou, C.; Turpin, S.M.; Matzaris, M.; Hoehn, K.L.; Hevener, A.L.; James, D.E.; Duh, E.J.; Watt, M.J. Pigment epithelium-derived factor contributes to insulin resistance in obesity. Cell Metab. 2009, 10, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg, M.L.; Andrews, Z.B.; Duh, E.J.; Meikle, P.J.; Batt, M.J. Pigment epithelium-derived factor regulates lipid metabolism via adipose triglyceride lipase. Diabetes 2011, 60, 1458–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnagarin, R.; Dharmarajan, A.; Dass, C.R. PEDF attenuates insulin-dependent molecular pathways of glucose homeostasis in skeletal myocytes. Mol. Cell Endocrinol. 2016, 422, 115–124. [Google Scholar] [CrossRef]
- Ek, E.T.; Contreras, K.G.; Dass, C.R.; Choong, P.F.M. Inhibition of orthotopic osteosarcoma growth and metastasis by multitargeted antitumor activities of pigment epithelium-derived factor. Clin. Exp. Metastasis 2007, 24, 93–106. [Google Scholar] [CrossRef]
- Lee, P.X.; Martinez, J.; Dass, C.R. Stimulation of bone regeneration with pigment epithelium-derived factor microparticles: Evidence in silico, in vitro and in vivo. Pharmazie 2016, 71, 382–389. [Google Scholar]
- Carnagarin, R.; Dharmarajan, A.; Dass, C.R. Insulin antagonises pigment epithelium-derived factor (PEDF)-induced modulation of lineage commitment of myocytes and heterotrophic ossification. Mol. Cell Endocrinol. 2018, 472, 159–166. [Google Scholar] [CrossRef]
- Elahy, M.; Doschak, M.R.; Hughes, J.D.; Baindur-Hudson, S.; Dass, C.R. Alginate bead-encapsulated PEDF induces ectopic bone formation in vivo in the absence of co-administered mesenchymal stem cells. Curr. Drug Targets 2018, 17, 467–478. [Google Scholar] [CrossRef]
- Chen, W.; Mao, M.; Fang, M.J.; Xie, Y.; Rui, Y. Fracture risk assessment in diabetes mellitus. Front. Endocrinol. 2022, 13, 961761. [Google Scholar] [CrossRef]
- Tacar, O.; Indumuthy, S.; Tan, M.L.; Baindur, S.; Friedhuber, A.M.; Dass, C.R. Cardiomyocyte apoptosis vs autophagy with prolonged doxorubicin treatment: Comparison with osteosarcoma cells. J. Pharm. Pharmacol. 2015, 67, 231–243. [Google Scholar] [CrossRef]
- Jones, I.C.; Dass, C.R. Doxorubicin-induced cardiotoxicity: Causative factors and possible interventions. J. Pharm. Pharmacol. 2022, 74, 1677–1688. [Google Scholar] [CrossRef]
- Tan, M.L.; Choong, P.F.M.; Dass, C.R. Anti-chondrosarcoma effects of PEDF mediated via molecules important to apoptosis, cell cycling, adhesion and invasion. Biochem. Biophys. Res. Commun. 2010, 398, 613–618. [Google Scholar] [CrossRef]
- Stone, R.L.; Bernlohr, D.A. The molecular basis for inhibition of adipose conversion of murine 3T3-L1 cells by retinoic acid. Differentiation 1990, 45, 119–127. [Google Scholar] [CrossRef]
- Ek, E.T.; Contreras, K.G.; Dass, C.R.; Choong, P.F.M. PEDF-derived synthetic peptides exhibit antitumor activity in an orthotopic model of human osteosarcoma. J. Orthop. Res. 2007, 25, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.L.; Choong, P.F.M.; Dass, C.R. The potential role of free chitosan in bone trauma and bone cancer management. Biomaterials 2014, 35, 7828–7838. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Tobe, K.; Tamemoto, H.; Yazaki, Y.; Kadowaki, T. Insulin signalling and insulin actions in the muscles and livers of insulin-resistant, insulin receptor substrate 1-deficient mice. Mol. Cell Biol. 1996, 16, 3074–3084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, K.; Liu, Y.F.; Shorer, H.; Hemi, R.; LeRoith, D.; Quan, M.; Kanety, H.; Seger, R.; Zick, Y. Phosphorylation of insulin receptor substrate-1 (IRS-1) by protein kinase B positively regulates IRS-1 function. J. Biol. Chem. 1997, 272, 29911–29918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.J.; Goldberg, J.L.; Qiao, L.Y.; Mitchell, J.J. Insulin-induced insulin receptor substrate-1 degradation is mediated by the proteasome degradation pathway. Diabetes 1999, 48, 1359–1364. [Google Scholar] [CrossRef] [PubMed]
- Carnagarin, R.; Dharmarajan, A.; Dass, C.R. Pigment epithelium-derived factor stimulates skeletal muscle glycolytic activity through NADPH oxidase-dependent reactive oxygen species production. Int. J. Biochem. Cell Biol. 2016, 78, 229–236. [Google Scholar] [CrossRef]
- Klaus, S. Functional differentiation of white and brown adipocytes. Bioessays 1997, 19, 215–223. [Google Scholar] [CrossRef]
- Qian, S.-W.; Tang, Y.; Li, X.; Liu, Y.; Zhang, Y.Y.; Huang, H.Y.; Xue, R.D.; Yu, H.Y.; Guo, L.; Gao, H.D.; et al. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, E798–E807. [Google Scholar] [CrossRef]
- Montanari, T.; Pošćić, N.; Colitti, M. Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: A review. Obes. Rev. 2017, 18, 495–513. [Google Scholar] [CrossRef]
- Rodeheffer, M.; Birsoy, K.; Friedman, J.M. Identification of white adipocyte progenitor cells in vivo. Cell 2008, 135, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Unser, A.; Tian, Y.; Xie, Y. Opportunities and challenges in three-dimensional brown adipogenesis of stem cells. Biotechnol. Adv. 2015, 133, 962–979. [Google Scholar] [CrossRef] [Green Version]
- Gattu, A.K.; Swenson, E.S.; Iwakiri, Y.; Samuel, V.T.; Troiano, N.; Berry, R.; Church, C.D.; Rodeheffer, M.S.; Carpenter, T.O.; Chung, C. Determination of mesenchymal stem cell fate by pigment epithelium-derived factor (PEDF) results in increased adiposity and reduced bone mineral content. FASEB J. 2013, 27, 4384–4394. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Song, N.; Tombran-Tink, J.; Niyibizi, C. Pigment epithelium derived factor suppresses expression of Sost/Sclerostin by osteocytes: Implication for its role in bone matrix mineralization. J. Cell Physiol. 2015, 230, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Burgers, T.A.; Williams, B.O. Regulation of Wnt/β-catenin signaling within and from osteocytes. Bone 2013, 54, 244–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doll, J.; Stellmach, V.M.; Bouck, N.P.; Bergh, A.R.; Lee, C.; Abramson, L.P.; Cornwell, M.L.; Pins, M.R. Pigment epithelium-derived factor regulates the vasculature and mass of the prostate and pancreas. Nat. Med. 2003, 9, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Angiogenesis modulates adipogenesis and obesity. J. Clin. Investig. 2007, 117, 2362–2368. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, H.; Golozoubova, V.; Cannon, B.; Nedergaard, J. UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living in thermal neutrality. Cell Metab. 2009, 9, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y. Adipose tissue angiogenesis as a therapeutic target for obesity and metabolic diseases. Nat. Rev. Drug Discov. 2010, 9, 107–115. [Google Scholar] [CrossRef]
- Tang, W.; Zeve, D.; Suh, J.M.; Bosnakovski, D.; Kyba, M.; Hammer, R.E.; Tallquist, M.D.; Graff, J.M. White fat progenitor cells reside in the adipose vasculature. Science 2008, 322, 583–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupnick, M.; Panigrahy, D.; Zhang, C.-Y.; Dallabrida, S.M.; Lowell, B.B.; Langer, R.; Folkman, M.J. Adipose tissue mass can be regulated through the vasculature. Proc. Natl. Acad. Sci. USA 2002, 99, 10730–10735. [Google Scholar] [CrossRef] [PubMed]
- Brakenhielm, E.; Cao, R.; Gao, B.; Angelin, B.; Cannon, B.; Parini, P.; Cao, Y. Angiogenesis inhibitor, TNP-470, prevents diet-induced and genetic obesity in mice. Circ. Res. 2004, 94, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.; Duda, D.G.; Perentes, J.Y.; Quadri, R.S.; Fukumura, D.; Jain, R.K. Blockade of VEGFR2 and not VEGFR1 can limit diet-induced fat tissue expansion: Role of local versus bone marrow-derived endothelial cells. PLoS ONE 2009, 4, e4974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Elahy, M.; Doschak, M.; Hughes, J.D.; Dass, C.R. Triple-threat activity of PEDF in bone tumors: Tumor inhibition, tissue preservation and cardioprotection against doxorubicin. Bone 2019, 124, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Yagihashi, S. Diabetes and pancreas size, does it matter? J. Diabetes Investig. 2017, 8, 413–415. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Carlessi, R.; Walz, N.; Cruzat, V.F.; Keane, K.; John, A.N.; Jiang, F.X.; Carnagarin, R.; Dass, C.R.; Newsholme, P. Pigment epithelium-derived factor (PEDF) regulates metabolism and insulin secretion from a clonal rat pancreatic beta cell line BRIN-BD11 and mouse islets. Mol. Cell Endocrinol. 2016, 426, 50–60. [Google Scholar] [CrossRef]
- Riccillo, F.; Bracamonte, M.I.; Montenegro, S.; Martínez, S.M.; Ronderos, J.R. Progressive histopathological changes and ß-cell loss in the pancreas of a new spontaneous rat model of type 2 diabetes. Tissue Cell 2012, 44, 101–110. [Google Scholar] [CrossRef]
- Wang, Q.; Jin, T. The role of insulin signaling in the development of β-cell dysfunction and diabetes. Islets 2009, 1, 95–101. [Google Scholar] [CrossRef]
- Dai, C.; Brissova, M.; Reinert, R.B.; Nyman, L.; Liu, E.H.; Thompson, C.; Shostak, A.; Shiota, M.; Takahashi, T.; Powers, A.C. Pancreatic islet vasculature adapts to insulin resistance through dilation and not angiogenesis. Diabetes 2013, 62, 4144–4153. [Google Scholar] [CrossRef] [Green Version]
- Tsuruhisa, S.; Matsui, T.; Koga, Y.; Sotokawauchi, A.; Yagi, M.; Yamagishi, S.-I. Pigment epithelium-derived factor inhibits advanced glycation end product-induced proliferation, VEGF and MMP-9 expression in breast cancer cells via interaction with laminin receptor. Oncol. Lett. 2021, 22, 629. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.-I.; Nakamura, K.; Ueda, S.; Kato, S.; Imaizumi, T. Pigment epithelium-derived factor (PEDF) blocks angiotensin II signaling in endothelial cells via suppression of NADPH oxidase: A novel anti-oxidative mechanism of PEDF. Cell Tissue Res. 2005, 320, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Achkar, I.W.; Kader, S.; Dib, S.S.; Suhre, K.; Halama, A. Metabolic signatures of tumor responses to doxorubicin elucidated by metabolic profiling in ovo. Metabolites 2020, 10, 268. [Google Scholar] [CrossRef] [PubMed]
- Lemma, S.; Di Pompo, G.; Porporato, P.E.; Sonveaux, P.; Avnet, S. MDA-MB-231 breast cancer cells fuel osteoclast metabolism and activity: A new rationale for the pathogenesis of osteolytic bone metastases. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 3254–3264. [Google Scholar] [CrossRef]
- Abooshahab, R.; Hoorshmand, K.; Luna, G.; Al-Salami, H.; Dass, C.R. Metabolomics profiling reveals the role of PEDF in triple-negative breast cancer cell MDA-MB-231 under glycaemic loading. Pharmaceutics 2023, 15, 543. [Google Scholar] [CrossRef]
- Walenta, S.; Wetterling, M.; Lehrke, M.; Rofstad, E.K.; Mueller-Klieser, W. High lactate levels predict likelihood of metastases, tumor recurrence, and restricted patient survival in human cervical cancers. Cancer Res. 2000, 60, 916–921. [Google Scholar]
- Huang, B.; Miao, H.; Yuan, Y.; Dong, H.; Zhang, Z. PEDF decreases cardiomyocyte edema during oxygen-glucose deprivation and recovery via inhibiting lactate accumulation and expression of AQP1. Int. J. Mol. Med. 2019, 43, 1979–1990. [Google Scholar] [CrossRef]
- Zhu, H.; Sarkar, S.; Scott, L.; Danelisen, I.; Trush, M.A.; Jia, Z.; Li, Y.R. A.; Jia, Z.; Li, Y.R. Doxorubicin redox biology: Redox cycling, topoisomerase inhibition, and oxidative stress. React. Oxyg. Species (Apex) 2016, 1, 189–198. [Google Scholar] [CrossRef] [Green Version]
- Farhane, Z.; Bonnier, F.; Maher, M.A.; Casey, A.; Byrne, H.J. Differentiating responses of lung cancer cell lines to doxorubicin exposure: In vitro Raman micro spectroscopy, oxidative stress and bcl-2 protein expression. J. Biophotonics 2017, 10, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Branco, A.F.; Sampaio, S.F.; Moreira, A.C.; Oliveira, P.J.; Sardão, V.A. Differentiation-dependent doxorubicin toxicity on H9c2 cardiomyoblasts. Cardiovasc. Toxicol. 2012, 12, 326–340. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharmacol. 2004, 142, 231–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, I.C.; Carnagarin, R.; Armstrong, J.; Lin, D.P.L.; Baxter-Holland, M.; Elahy, M.; Dass, C.R. Pigment Epithelium-Derived Factor: Inhibition of Phosphorylation of Insulin Receptor (IR)/IR Substrate (IRS), Osteogeneration from Adipocytes, and Increased Levels Due to Doxorubicin Exposure. Pharmaceutics 2023, 15, 1960. https://doi.org/10.3390/pharmaceutics15071960
Jones IC, Carnagarin R, Armstrong J, Lin DPL, Baxter-Holland M, Elahy M, Dass CR. Pigment Epithelium-Derived Factor: Inhibition of Phosphorylation of Insulin Receptor (IR)/IR Substrate (IRS), Osteogeneration from Adipocytes, and Increased Levels Due to Doxorubicin Exposure. Pharmaceutics. 2023; 15(7):1960. https://doi.org/10.3390/pharmaceutics15071960
Chicago/Turabian StyleJones, Isobel C., Revathy Carnagarin, Jo Armstrong, Daphne P. L. Lin, Mia Baxter-Holland, Mina Elahy, and Crispin R. Dass. 2023. "Pigment Epithelium-Derived Factor: Inhibition of Phosphorylation of Insulin Receptor (IR)/IR Substrate (IRS), Osteogeneration from Adipocytes, and Increased Levels Due to Doxorubicin Exposure" Pharmaceutics 15, no. 7: 1960. https://doi.org/10.3390/pharmaceutics15071960