
New Imidazolium Alkaloids with Broad Spectrum of Action from the Marine Bacterium Shewanella aquimarina
 , , , , , , , ,
, , , , , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Media, and Buffers
- LB broth (Lennox) (Condalab, Madrid, Spain);
- Marine Broth (MB) (Condalab, Madrid, Spain);
- Trypticasein Soy Broth (TSB) (Condalab, Madrid, Spain);
- Thioglycollate Medium ISO (TGB) (Condalab, Madrid, Spain);
- ISP2 (glucose 4 g/L, YE 4 g/L, malt extract 10 g/L);
- AUR (starch 10 g/L, glucose 10 g/L, glycerol 10 g/L, peptone 5 g/L, and YE 2 g/L);
- Mineral Salt Medium (MSM) (KH2PO4 0.7 g/L, Na2HPO4 0.9 g/L, NaNO3 2 g/L, MgSO4 0.4 g/L, and CaCl2 0.1 g/L) supplemented with:
- glucose 10 g/L (MSM Glu);
- glucose 10 g/L and arginine 30 mM (MSM Glu Arg).
- Mueller Hinton (MH) broth (Merck KgaA, Darmstadt, Germany);
- MH supplemented with 1% (w/v) glucose (MHG);
- Dulbecco’s phosphate buffered saline (DPBS) (Microgem, Naples, Italy).
2.2. Propagation and Maintenance of Biological Specimens
2.2.1. Cell Line and Viruses
2.2.2. Caenorhabditis Elegans Propagation and Maintenance
2.3. Cultivation and Extracts Production
2.4. Extract Fractionation
2.5. Liquid Chromatography–High-Resolution Tandem Mass Spectrometry (LC-HRMS2)
2.6. Feature-Based Molecular Networking and Spectral Library Search
2.7. Oil-Spreading Assay
2.8. Antibacterial Assay
2.8.1. MBC Assay
2.8.2. Antimicrobial Synergy and Checkerboard Testing
- FICI ≤ 0.5, synergy;
- 0.5 < FICI ≤ 1.0, partial synergy;
- 1.0 < FICI ≤ 4.0, no interaction;
- FICI > 4.0, antagonism.
2.9. Antibiofilm Activity
2.9.1. Biofilm Initial Cell Attachment Assay
2.9.2. Biofilm Inhibition Assay
2.9.3. Biofilm Degradation Assay
2.10. Antiviral Assays
2.10.1. Co-Treatment Assay
2.10.2. Virus Pre-Treatment Assay
2.10.3. Post-Treatment Assay
2.10.4. Cell Pre-Treatment
2.11. Anthelmintic Activity
2.12. Statistical Analysis
3. Results and Discussion
3.1. Production and Extraction of Secondary Metabolites from S. aquimarina
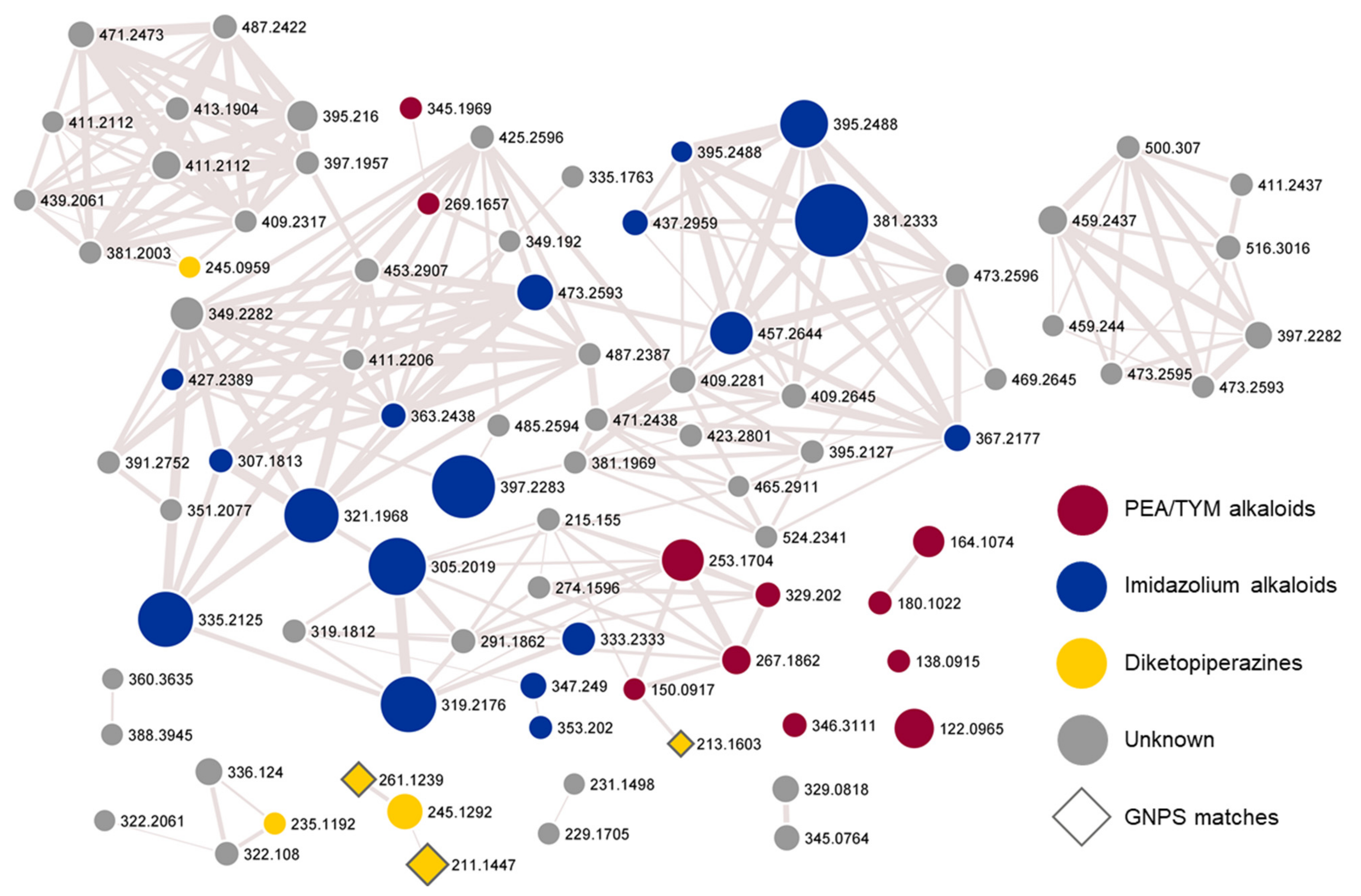
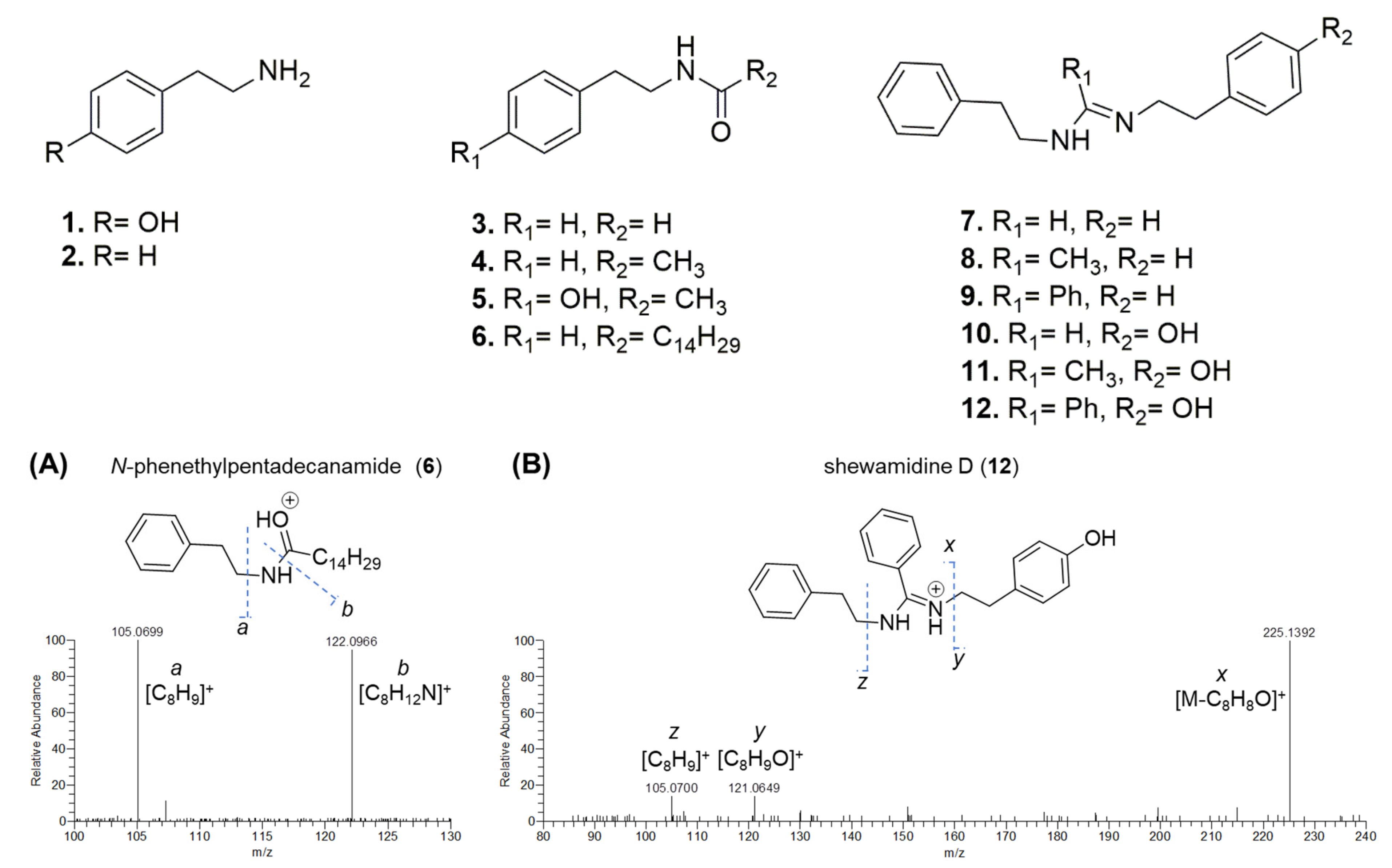
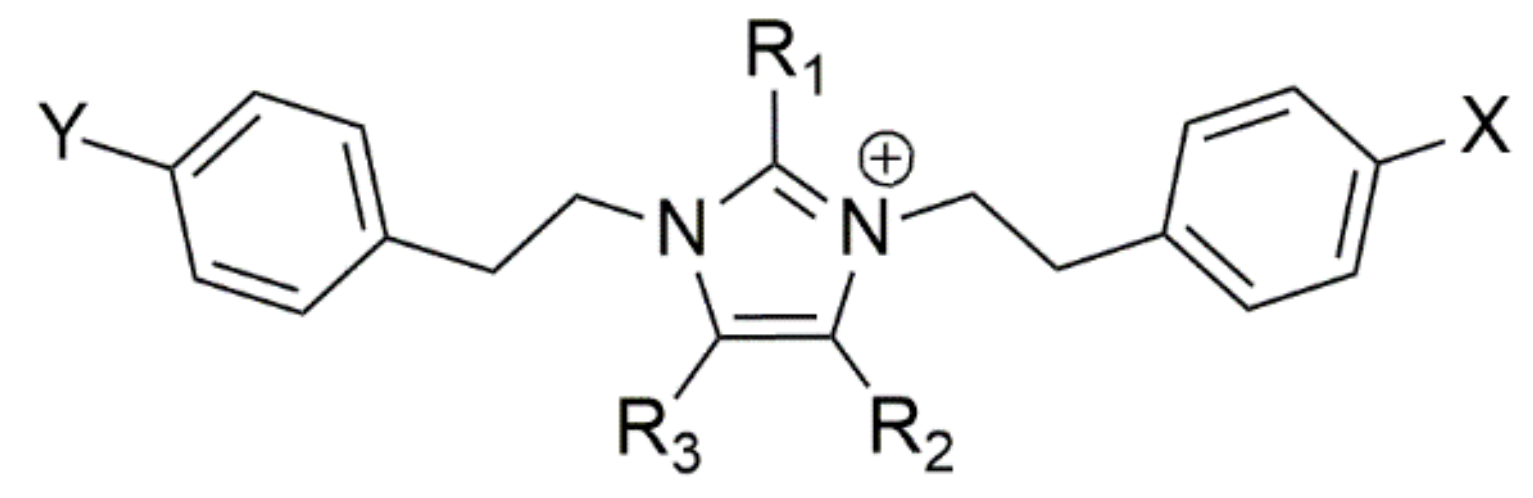
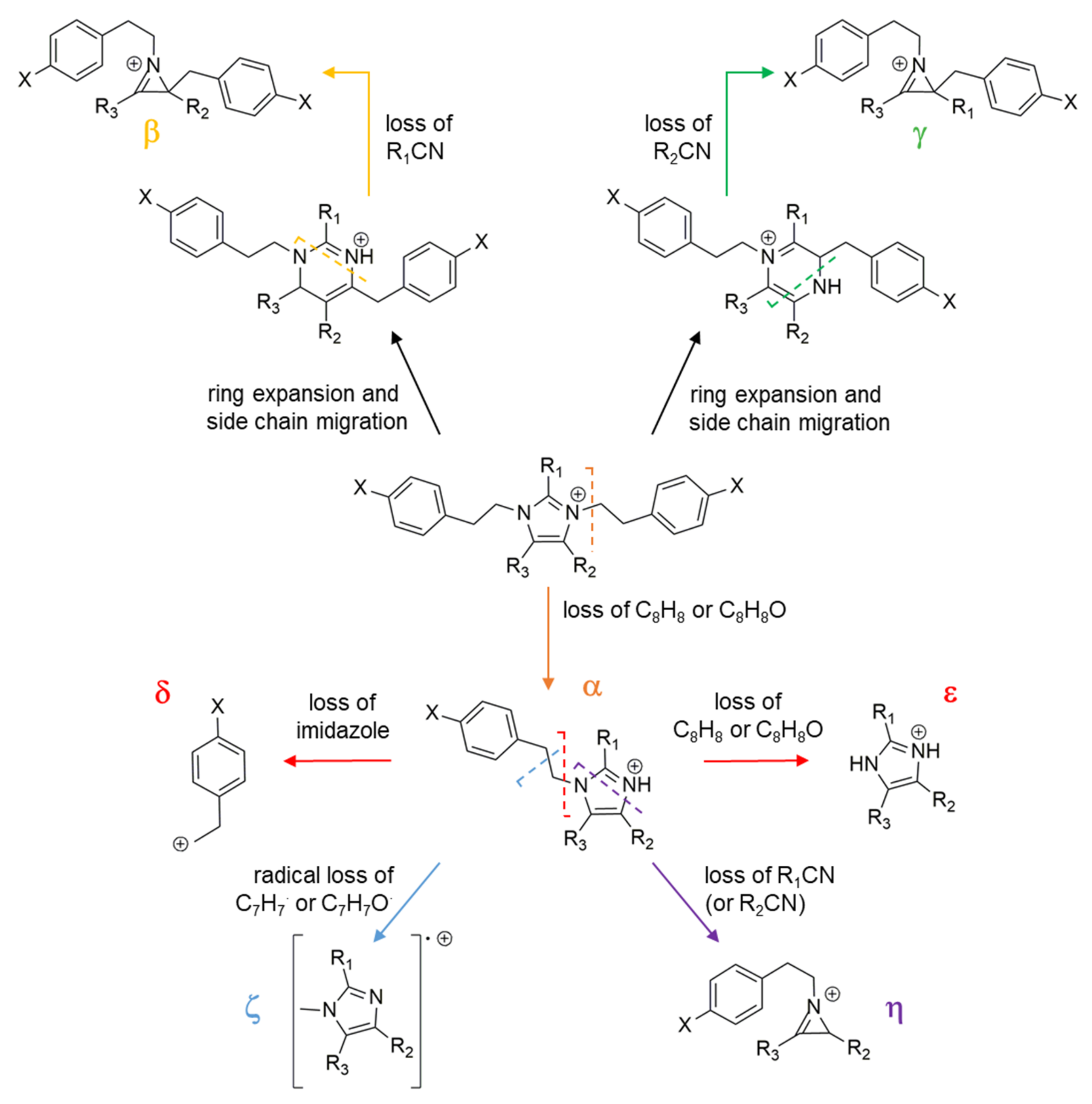
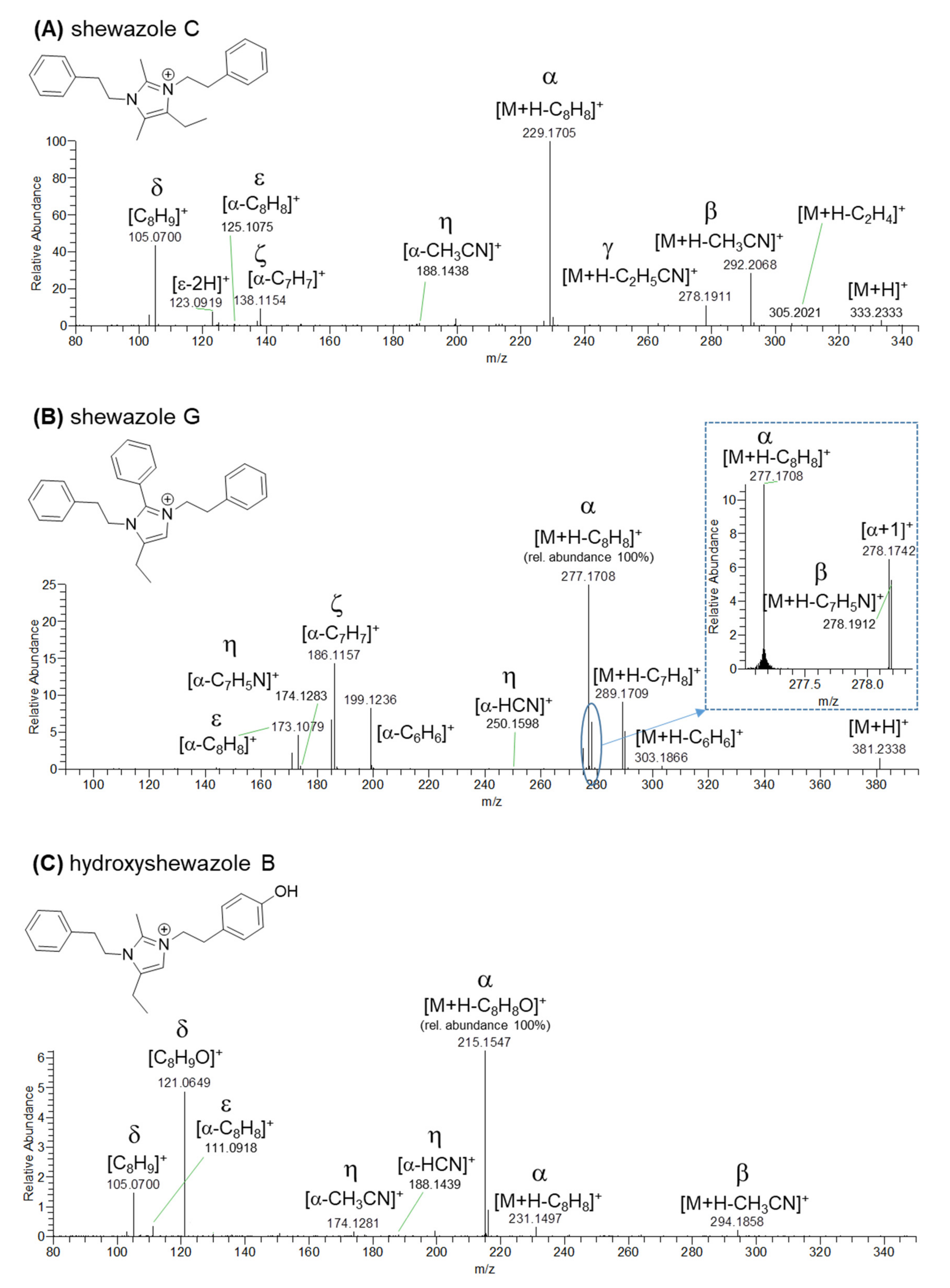
3.2. Extract Fractionation and Structural Prediction of Alkaloids from the F2 Fraction
3.3. F2 Biosurfactant Activity
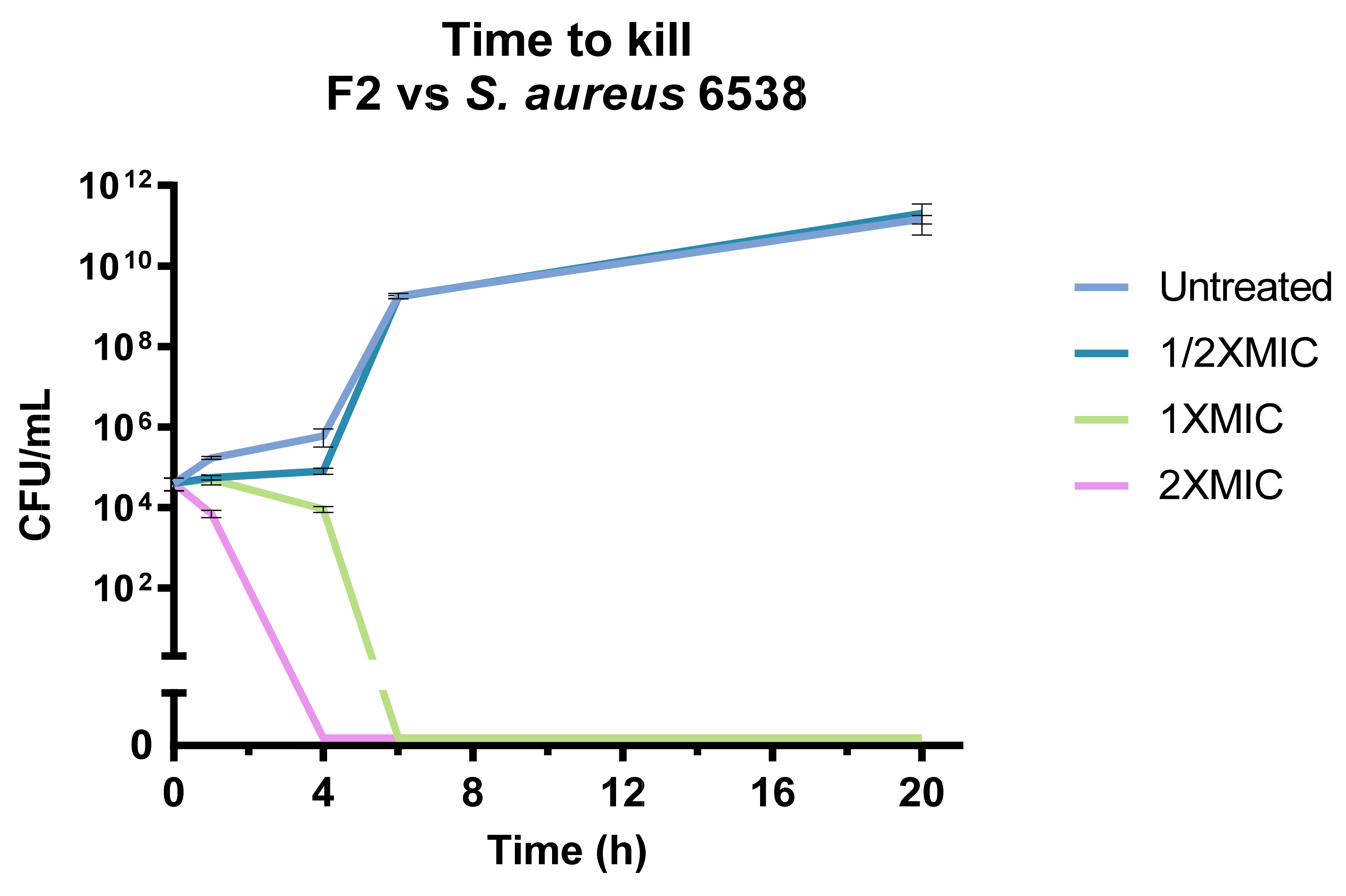
3.4. F2 Antimicrobial Activity against Clinically Isolated S. aureus Strains
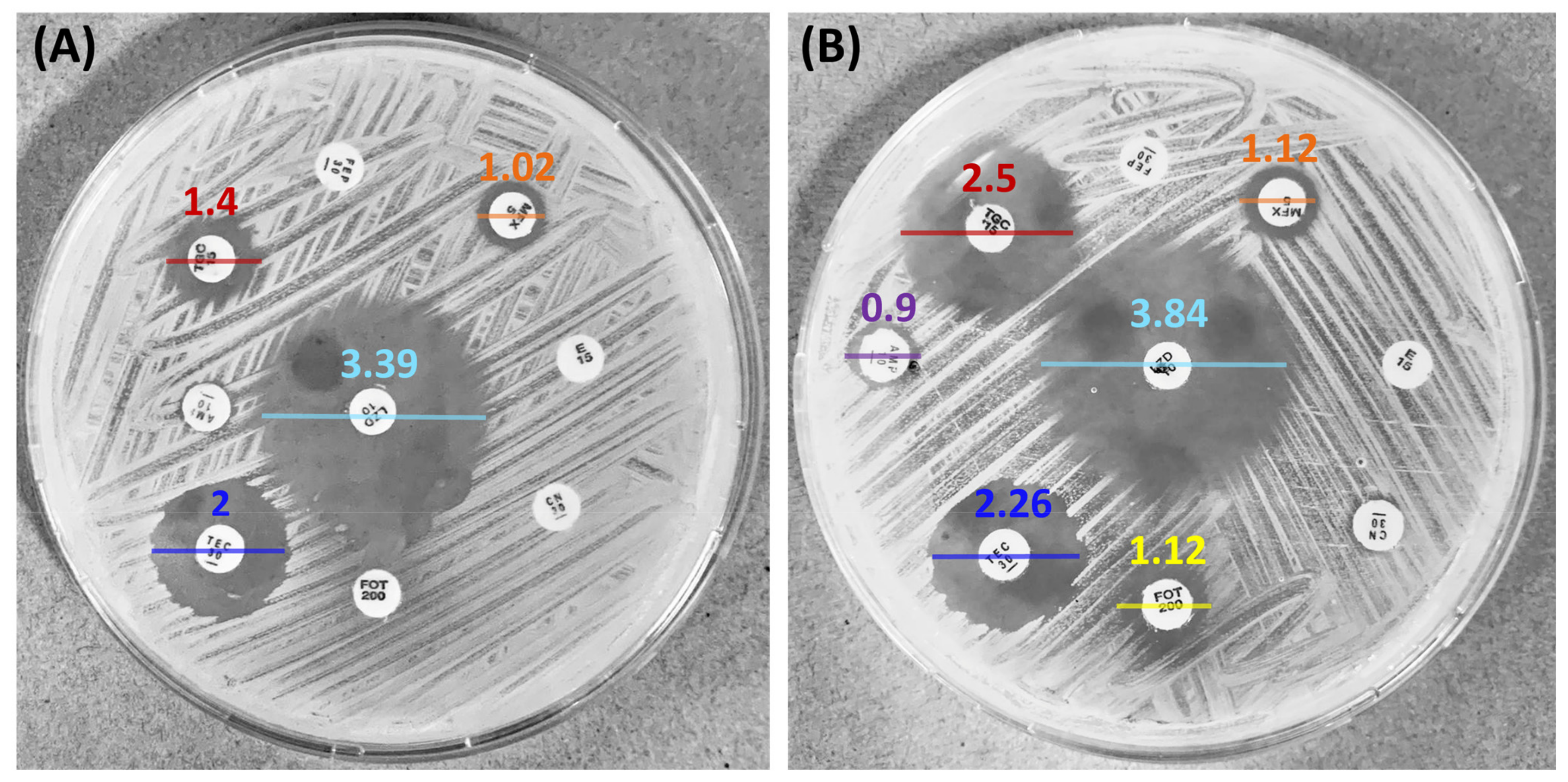
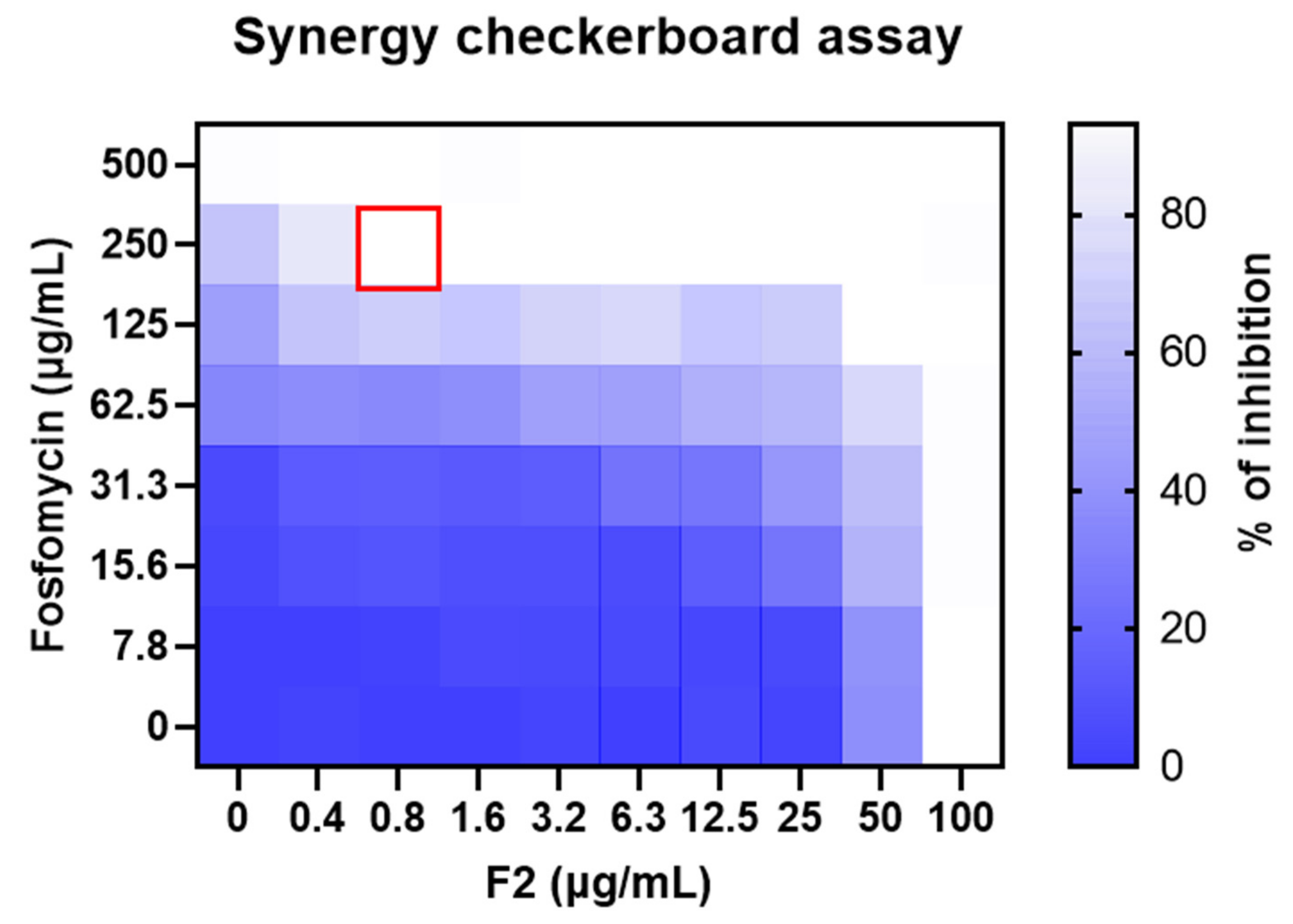
3.5. Synergistic Effect of Antibiotics and F2 on MRSA
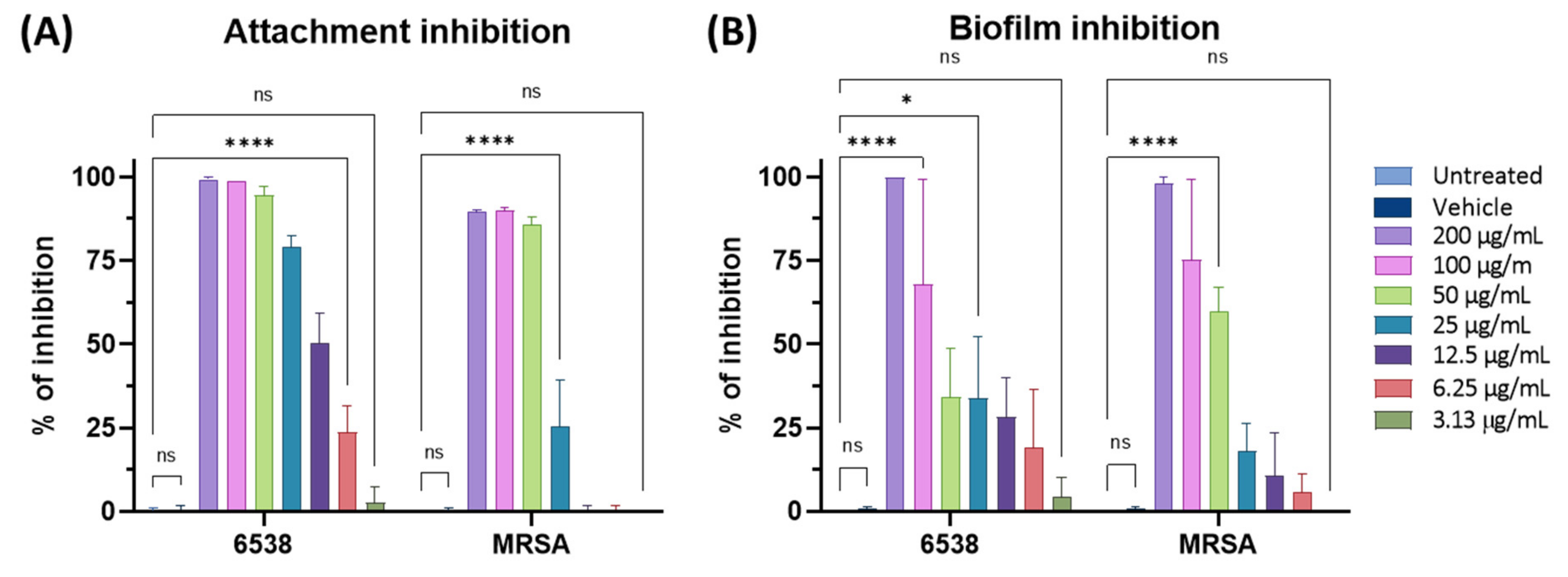
3.6. Antibiofilm Activity of F2 on S. aureus
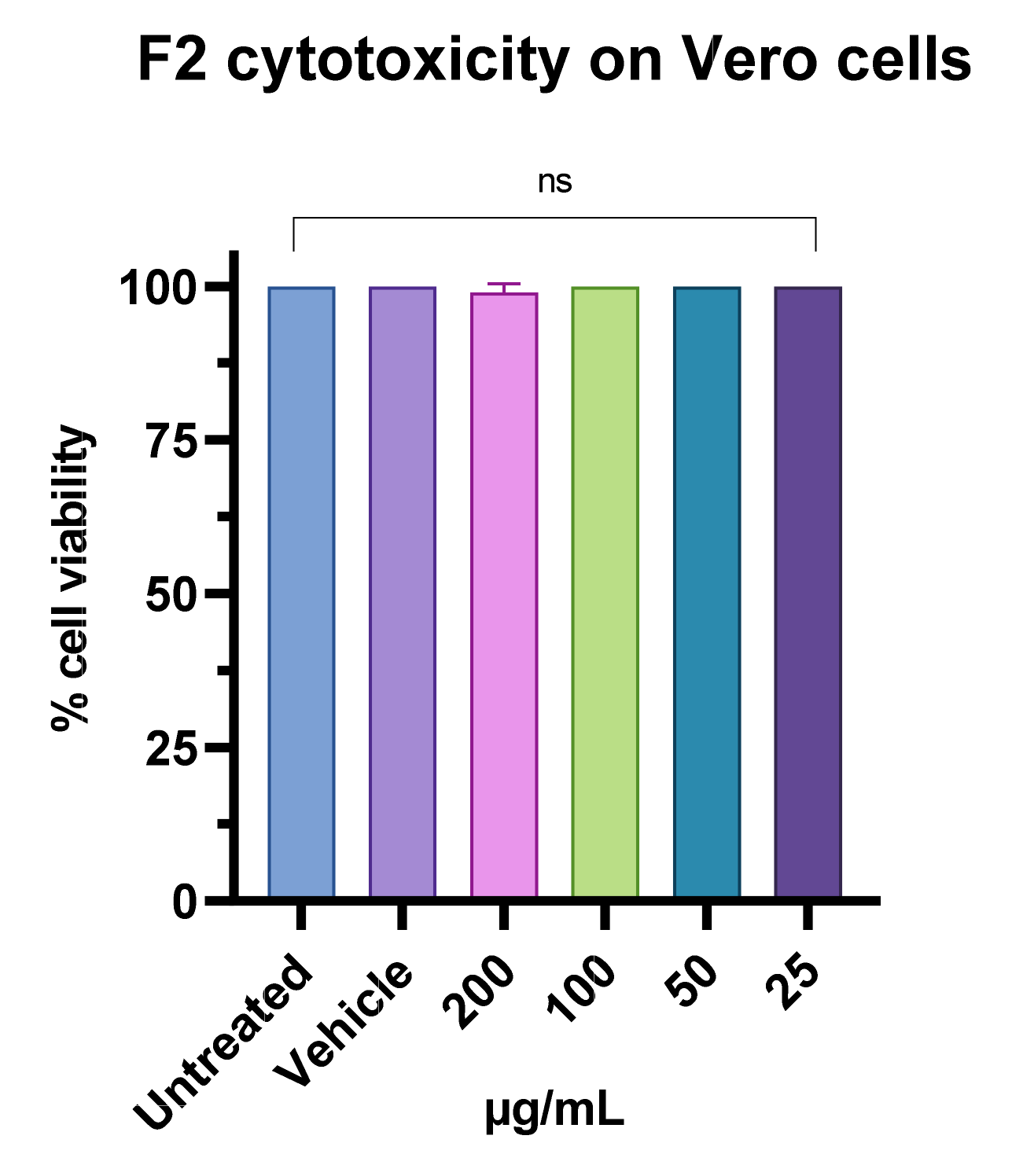
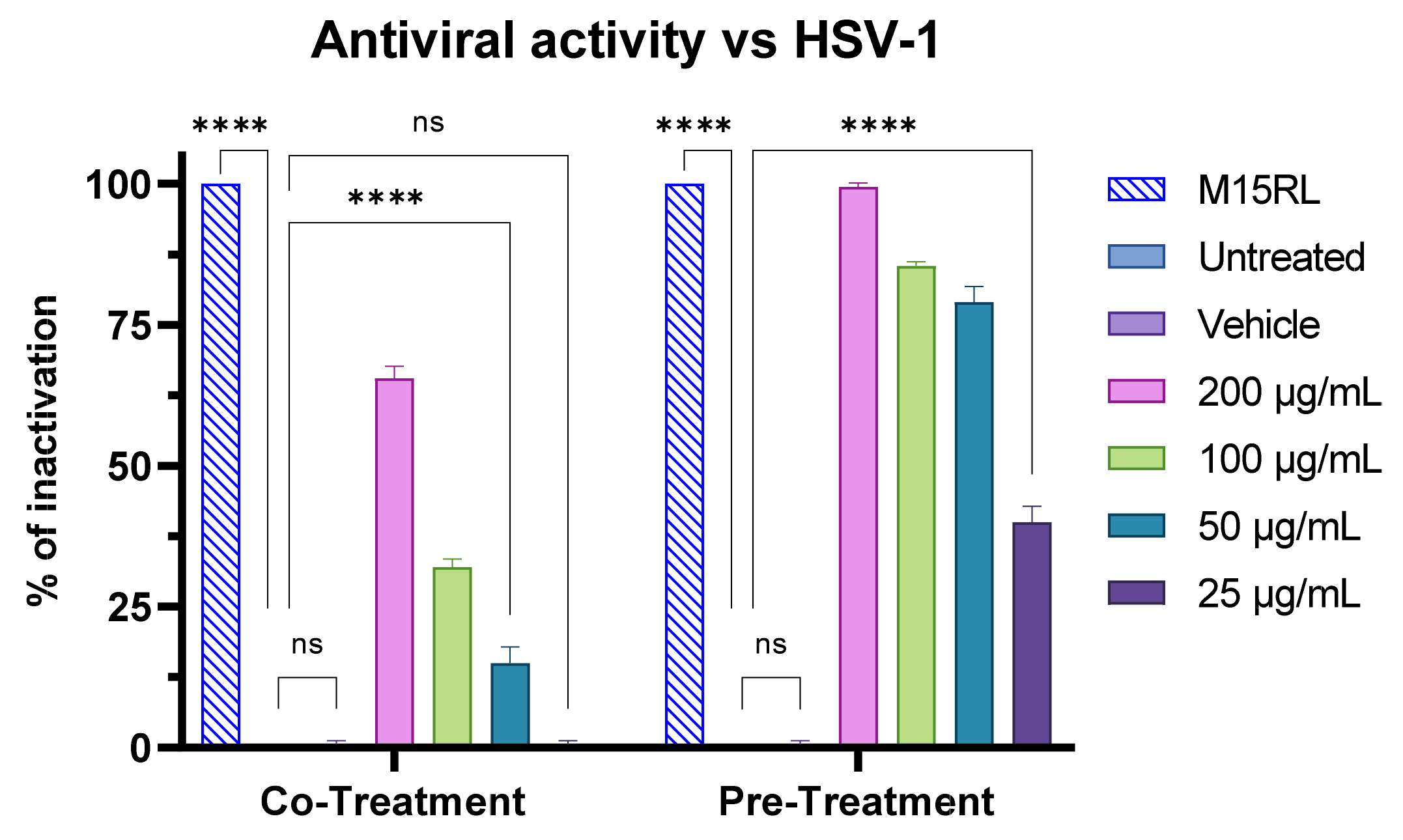
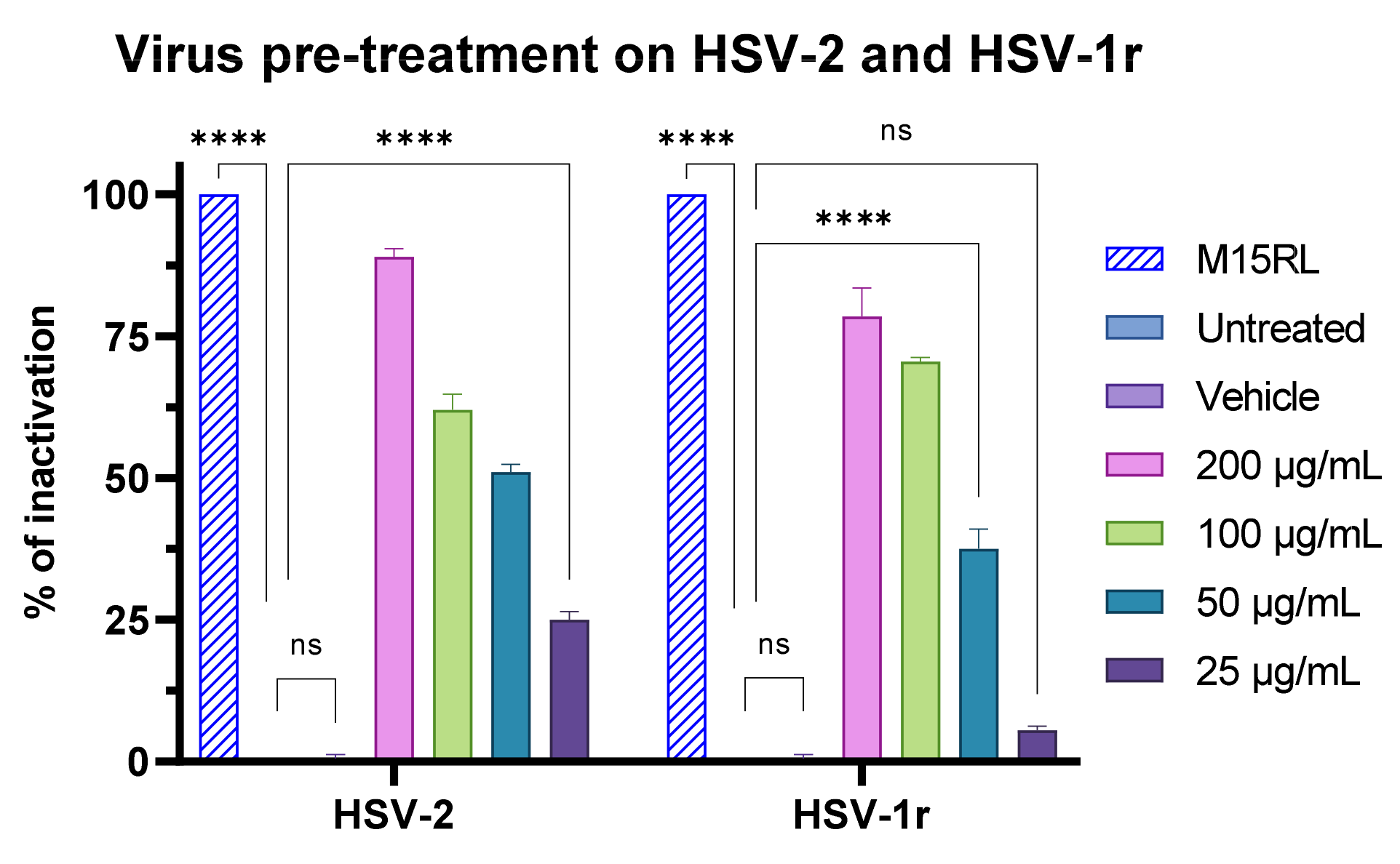
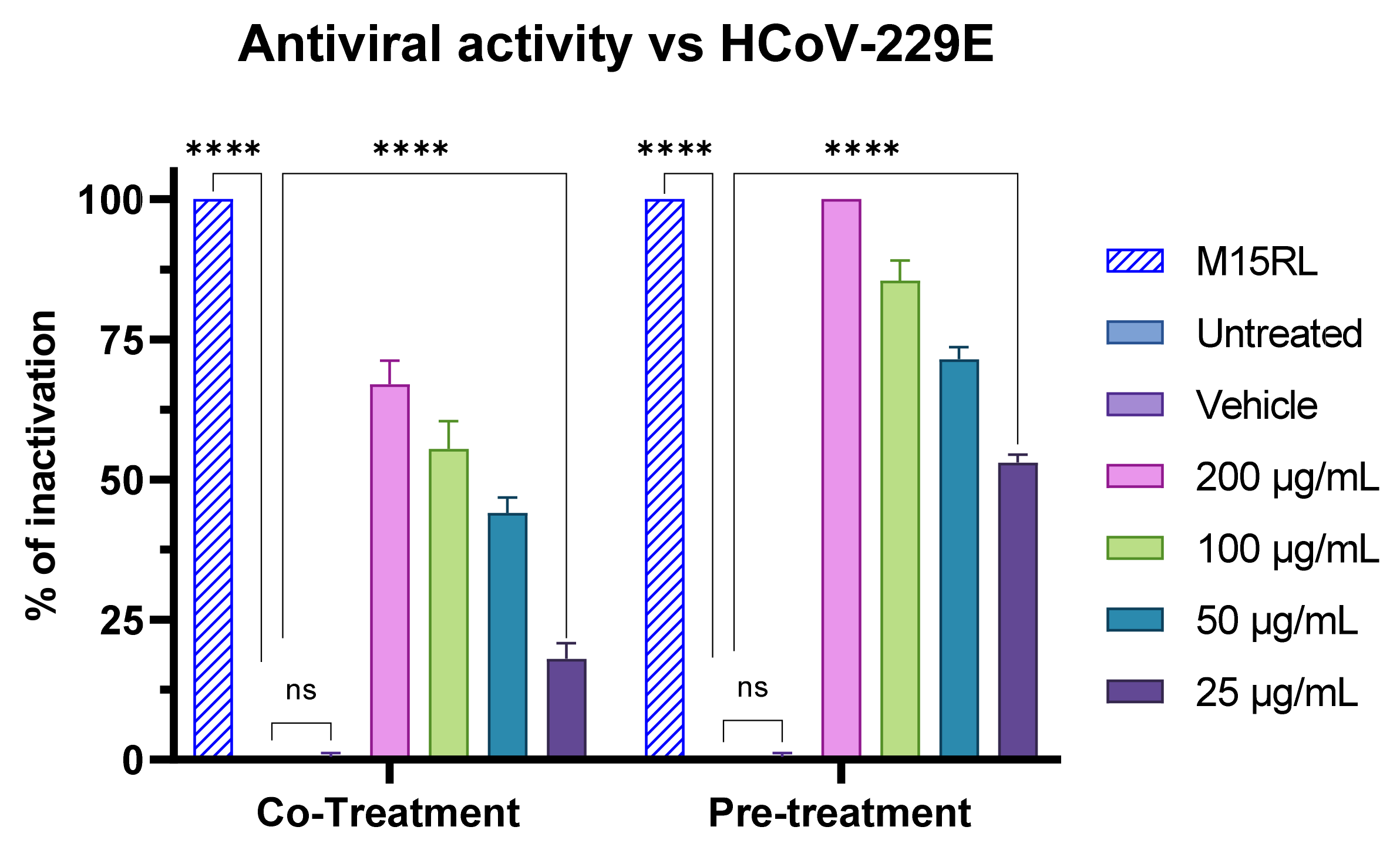
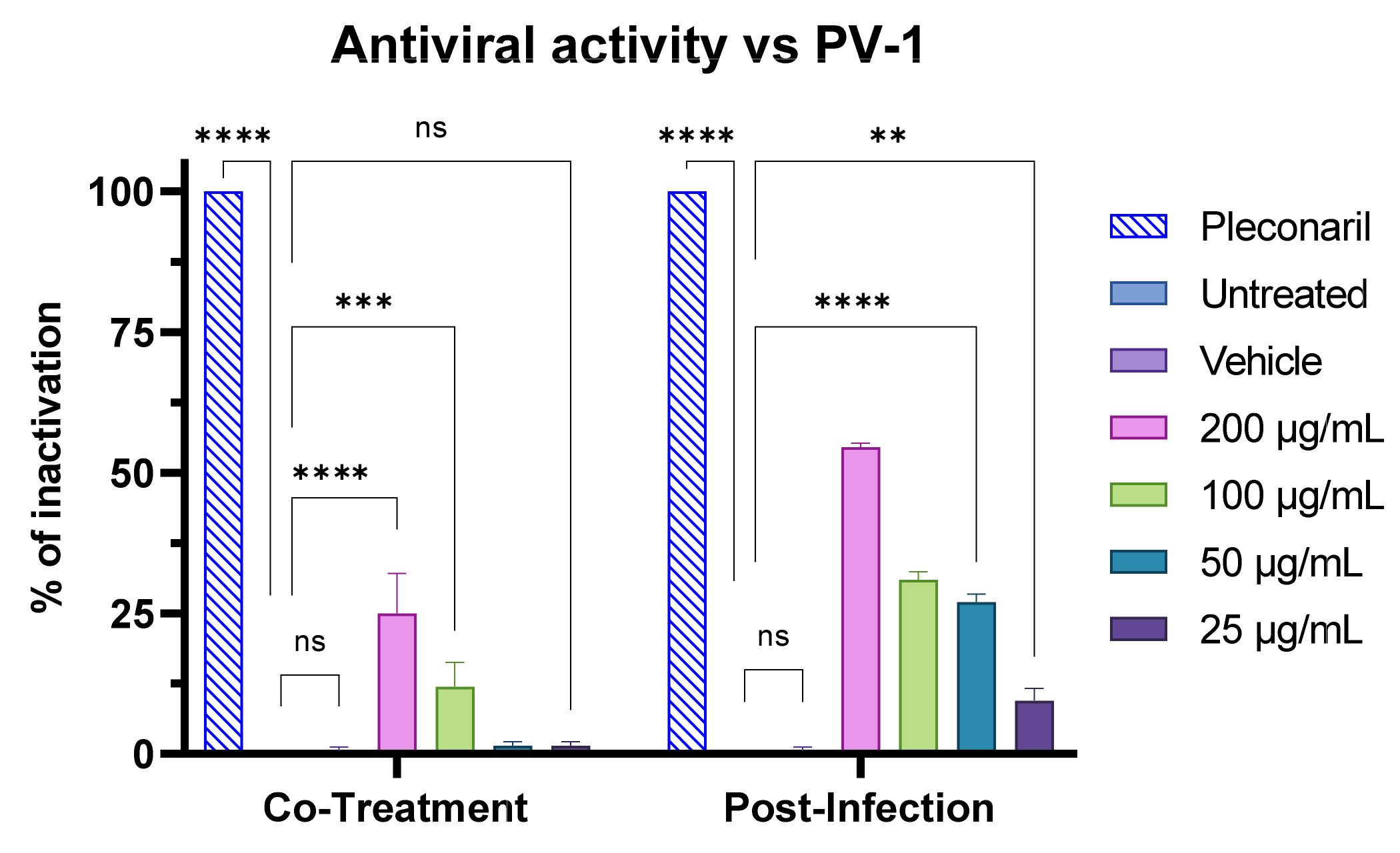
3.7. Antiviral Activity of F2
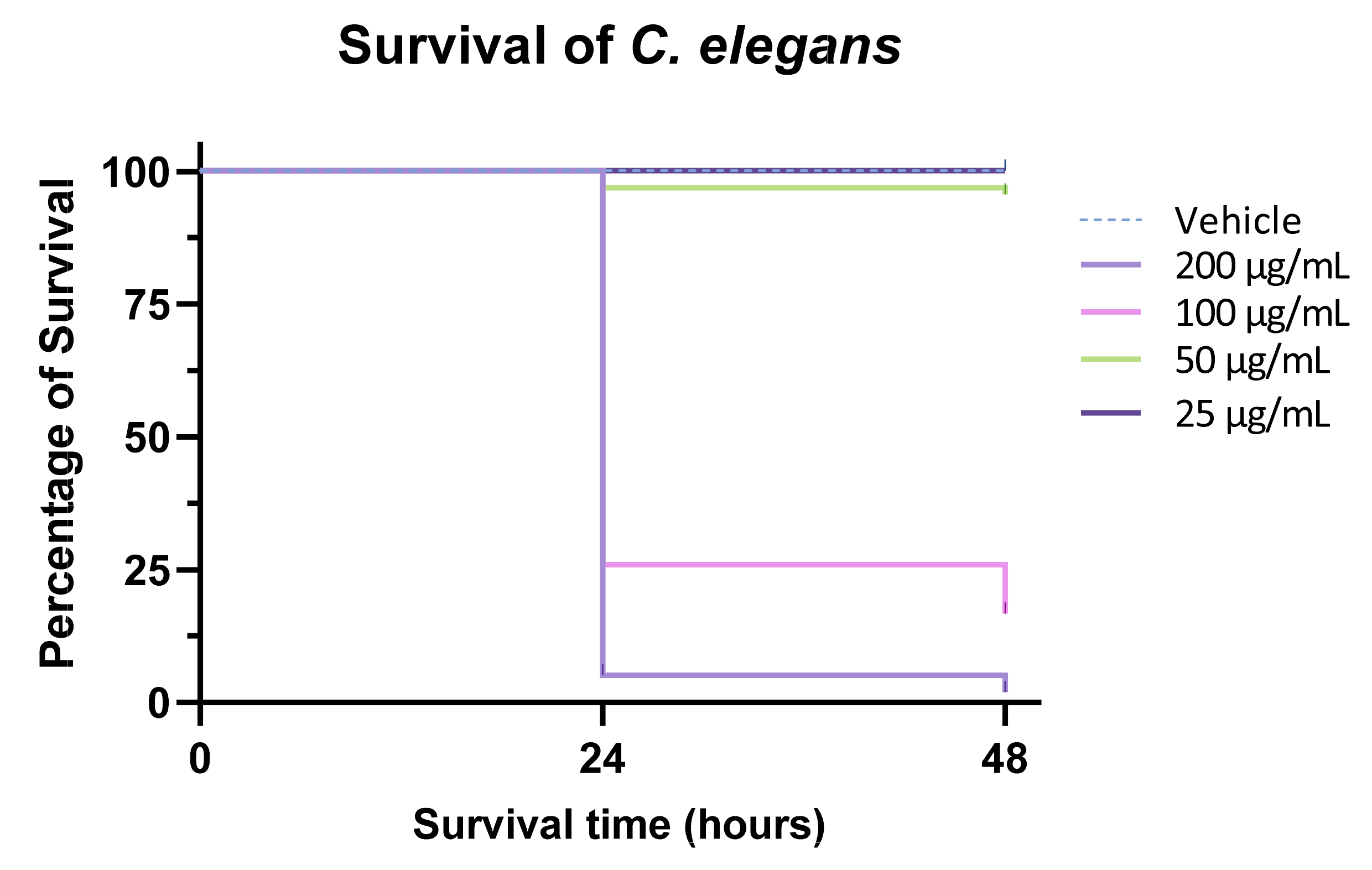
3.8. Anthelmintic Activity of F2 on C. elegans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMP | Ampicillin |
| CFUs | Colonies forming units |
| CMC | Carboxymethylcellulose |
| CN | Gentamycin |
| CV | Crystal violet |
| DKPs | Diketopiperazines |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | Dimethyl sulfoxide |
| DPBS | Dulbecco’s phosphate buffered saline |
| E | Erythromycin |
| EtOAc | Ethyl acetate |
| FBMN | Feature-Based Molecular Networking |
| FBS | Fetal Bovine Serum |
| FEP | Cefepime |
| FICI | Fractional inhibitory concentration index |
| FOT | Fosfomycin |
| HCoVs | Human coronaviruses |
| HRMS/MS | High-resolution tandem mass spectrometry |
| HSV-1 | Herpes simplex virus type 1 |
| HSV-1r | HSV-1 acyclovir-resistant |
| HSV-2 | Herpes simplex virus type 2 |
| IHME | Institute for Health Metrics and Evaluation |
| LC HRMS/MS | Liquid Chromatography–High-Resolution Tandem Mass Spectrometry |
| LZD | Linezolid |
| MB | Marine Broth |
| MBC | Minimal bactericidal concentration |
| MeOH | Methanol |
| MFX | Moxifloxacin |
| MH | Muller Hinton |
| MIC | Minimum inhibitory concentration |
| MLSB | Constitutive resistance to macrolides |
| MOI | Multiplicity of infection |
| MRSA | Methicillin resistance |
| MSM | Mineral Salt Medium |
| MSSA | Multisensitive |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| NGM | Nematode Growth Medium |
| NTDs | Neglected tropical diseases |
| OD | Optical density |
| OSMAC | One strain many compounds |
| PEA | Phenethylamine |
| PFU | Plaque-forming unit |
| PV-1 | Poliovirus type-1 |
| QRSA | Quinolones resistance |
| RT | Room Temperature |
| SARS-CoV-2 | Severe Acute Respiratory Syndrome coronavirus 2 |
| SPE | Solid phase extraction |
| TEC | Teicloplanin |
| TGB | Thioglycollate |
| TGC | Tigecycline |
| TSB | Trypticasein Soy Broth |
| TYM | Tyramine |
References
- Roser, M.; Ritchie, H.; Spooner, F. Burden of Disease. 2021. Available online: https://ourworldindata.org/burden-of-disease (accessed on 10 August 2023).
- Craft, K.M.; Nguyen, J.M.; Berg, L.J.; Townsend, S.D. Methicillin-resistant Staphylococcus aureus (MRSA): Antibiotic-resistance and the biofilm phenotype. MedChemComm 2019, 10, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Abraham, L.; Bamberger, D.M. Staphylococcus aureus Bacteremia: Contemporary Management. Mo. Med. 2020, 117, 341–345. [Google Scholar]
- Kranjec, C.; Morales Angeles, D.; Torrissen Marli, M.; Fernandez, L.; Garcia, P.; Kjos, M.; Diep, D.B. Staphylococcal Biofilms: Challenges and Novel Therapeutic Perspectives. Antibiotics 2021, 10, 131. [Google Scholar] [CrossRef] [PubMed]
- Burrell, C.J.; Howard, C.R.; Murphy, F.A. Epidemiology of Viral Infections. In Fenner and White’s Medical Virology; Academic Press: Cambridge, MA, USA, 2017; pp. 185–203. [Google Scholar] [CrossRef]
- Malary, M.; Abedi, G.; Hamzehgardeshi, Z.; Afshari, M.; Moosazadeh, M. The prevalence of herpes simplex virus type 1 and 2 infection in Iran: A meta-analysis. Int. J. Reprod. Biomed. 2016, 14, 615–624. [Google Scholar]
- Jiang, Y.C.; Feng, H.; Lin, Y.C.; Guo, X.R. New strategies against drug resistance to herpes simplex virus. Int. J. Oral Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef]
- Liu, D.X.; Liang, J.Q.; Fung, T.S. Human Coronavirus-229E, -OC43, -NL63, and -HKU1 (Coronaviridae). In Encyclopedia of Virology; Academic Press: Cambridge, MA, USA, 2021; pp. 428–440. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Joseph, A.; Nair, B.G. Promising bioactive compounds from the marine environment and their potential effects on various diseases. J. Genet. Eng. Biotechnol. 2022, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.J.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- Rotter, A.; Gaudêncio, S.P.; Klun, K.; Macher, J.-N.; Thomas, O.P.; Deniz, I.; Edwards, C.; Grigalionyte-Bembič, E.; Ljubešić, Z.; Robbens, J.; et al. A New Tool for Faster Construction of Marine Biotechnology Collaborative Networks. Front. Mar. Sci. 2021, 8, 685164. [Google Scholar] [CrossRef]
- Zhou, Q.; Hotta, K.; Deng, Y.; Yuan, R.; Quan, S.; Chen, X. Advances in Biosynthesis of Natural Products from Marine Microorganisms. Microorganisms 2021, 9, 2551. [Google Scholar] [CrossRef]
- König, G.M.; Kehraus, S.; Seibert, S.F.; Abdel-Lateff, A.; Müller, D. Natural Products from Marine Organisms and Their Associated Microbes. ChemBioChem 2006, 7, 229–238. [Google Scholar] [CrossRef]
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine Bacterial Secondary Metabolites: A Treasure House for Structurally Unique and Effective Antimicrobial Compounds. Mar. Drugs 2021, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- De Santi, C.; Tedesco, P.; Ambrosino, L.; Altermark, B.; Willassen, N.P.; de Pascale, D. A new alkaliphilic cold-active esterase from the psychrophilic marine bacterium Rhodococcus sp.: Functional and structural studies and biotechnological potential. Appl. Biochem. Biotechnol. 2014, 172, 3054–3068. [Google Scholar] [CrossRef] [PubMed]
- Hurdle, J.G.; O’Neill, A.J.; Chopra, I.; Lee, R.E. Targeting bacterial membrane function: An underexploited mechanism for treating persistent infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjerk, T.R.; Severino, P.; Jain, S.; Marques, C.; Silva, A.M.; Pashirova, T.; Souto, E.B. Biosurfactants: Properties and Applications in Drug Delivery, Biotechnology and Ecotoxicology. Bioengineering 2021, 8, 115. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, P.; Maida, I.; Palma Esposito, F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial Activity of Monoramnholipids Produced by Bacterial Strains Isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallimore, W. Chapter 18—Marine Metabolites: Oceans of Opportunity. In Pharmacognosy; Badal, S., Delgoda, R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 377–400. [Google Scholar] [CrossRef]
- Abbas, S.; Mahmoud, H. Identification of Sponge-Associated Bacteria from the Coast of Kuwait and Their Potential Biotechnological Applications. Front. Microbiol. 2022, 13, 896718. [Google Scholar] [CrossRef] [PubMed]
- Qasim, M.S.; Lampi, M.; Heinonen, M.K.; Garrido-Zabala, B.; Bamford, D.H.; Kakela, R.; Roine, E.; Sarin, L.P. Cold-Active Shewanella glacialimarina TZS-4(T) nov. Features a Temperature-Dependent Fatty Acid Profile and Putative Sialic Acid Metabolism. Front. Microbiol. 2021, 12, 737641. [Google Scholar] [CrossRef]
- Teli, P.; Sahiba, N.; Sethiya, A.; Soni, J.; Agarwal, S. Imidazole derivatives: Impact and prospects in antiviral drug discovery. In Imidazole-Based Drug Discovery; Elsevier: Amsterdam, The Netherlands, 2022; pp. 167–193. [Google Scholar] [CrossRef]
- Cui, B.; Zheng, B.L.; He, K.; Zheng, Q.Y. Imidazole Alkaloids from Lepidium meyenii and Methods of Usage. US6878731B2, 12 April 2005. [Google Scholar]
- Mingzhang, A.; Wenwen, J.; Longjang, Y. Application of Maca Imidazole Alkaloid in Preparation of Cardiovascular Drugs. CN101756965A, 30 June 2010. [Google Scholar]
- Wang, L.; Linares-Otoya, V.; Liu, Y.; Mettal, U.; Marner, M.; Armas-Mantilla, L.; Willbold, S.; Kurtán, T.; Linares-Otoya, L.; Schäberle, T.F. Discovery and Biosynthesis of Antimicrobial Phenethylamine Alkaloids from the Marine Flavobacterium Tenacibaculum discolor sv11. J. Nat. Prod. 2022, 85, 1039–1051. [Google Scholar] [CrossRef]
- Yan, J.-X.; Wu, Q.; Helfrich, E.J.N.; Chevrette, M.G.; Braun, D.R.; Heyman, H.; Ananiev, G.E.; Rajski, S.R.; Currie, C.R.; Clardy, J.; et al. Bacillimidazoles A-F, Imidazolium-Containing Compounds Isolated from a Marine Bacillus. Mar. Drugs 2022, 20, 43. [Google Scholar] [CrossRef]
- Jin, Z. Muscarine, imidazole, oxazole, and thiazole alkaloids. Nat. Prod. Rep. 2011, 28, 1143–1191. [Google Scholar] [CrossRef]
- Tanaka, N.; Kusama, T.; Takahashi-Nakaguchi, A.; Gonoi, T.; Fromont, J.; Kobayashi, J. Nagelamides X–Z, Dimeric Bromopyrrole Alkaloids from a Marine Sponge Agelas sp. Org. Lett. 2013, 15, 3262–3265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, B.; Prasad, P.; Capon, R.J.; Jia, Y. Asymmetric Total Synthesis of (+)-Dragmacidin D Reveals Unexpected Stereocomplexity. Org. Lett. 2015, 17, 1529–1532. [Google Scholar] [CrossRef] [PubMed]
- Gong, A.D.; Li, H.P.; Shen, L.; Zhang, J.B.; Wu, A.B.; He, W.J.; Yuan, Q.S.; He, J.D.; Liao, Y.C. The Shewanella algae strain YM8 produces volatiles with strong inhibition activity against Aspergillus pathogens and aflatoxins. Front. Microbiol. 2015, 6, 1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franci, G.; Folliero, V.; Cammarota, M.; Zannella, C.; Sarno, F.; Schiraldi, C.; de Lera, A.R.; Altucci, L.; Galdiero, M. Epigenetic modulator UVI5008 inhibits MRSA by interfering with bacterial gyrase. Sci. Rep. 2018, 8, 13117. [Google Scholar] [CrossRef] [PubMed]
- Stiernagle, T. Maintenance of C. elegans. WormBook 2006, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Sala, G.; Coppola, D.; Virgili, R.; Vitale, G.A.; Tanduo, V.; Teta, R.; Crocetta, F.; de Pascale, D. Untargeted Metabolomics Yields Insights into the Lipidome of Botrylloides niger Herdman, 1886, An Ascidian Invading the Mediterranean Sea. Front. Mar. Sci. 2022, 9, 865751. [Google Scholar] [CrossRef]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Meth. 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Scarpato, S.; Teta, R.; Della Sala, G.; Pawlik, J.R.; Costantino, V.; Mangoni, A. New Tricks with an Old Sponge: Feature-Based Molecular Networking Led to Fast Identification of New Stylissamide L from Stylissa caribica. Mar. Drugs 2020, 18, 443. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Hirata, Y.; Imanaka, T. A study on the structure–function relationship of lipopeptide biosurfactants. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2000, 1488, 211–218. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018; pp. 19087–19898. [Google Scholar]
- Buonocore, C.; Giugliano, R.; Della Sala, G.; Palma Esposito, F.; Tedesco, P.; Folliero, V.; Galdiero, M.; Franci, G.; de Pascale, D. Evaluation of Antimicrobial Properties and Potential Applications of Pseudomonas gessardii M15 Rhamnolipids towards Multiresistant Staphylococcus aureus. Pharmceutics 2023, 15, 700. [Google Scholar] [CrossRef] [PubMed]
- Folliero, V.; Franci, G.; Dell’Annunziata, F.; Giugliano, R.; Foglia, F.; Sperlongano, R.; De Filippis, A.; Finamore, E.; Galdiero, M. Evaluation of Antibiotic Resistance and Biofilm Production among Clinical Strain Isolated from Medical Devices. Int. J. Microbiol. 2021, 2021, 9033278. [Google Scholar] [CrossRef] [PubMed]
- Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Palma Esposito, F.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics 2021, 13, 2121. [Google Scholar] [CrossRef] [PubMed]
- Gharaei, S.; Ohadi, M.; Hassanshahian, M.; Porsheikhali, S.; Forootanfar, H. Isolation, Optimization, and Structural Characterization of Glycolipid Biosurfactant Produced by Marine Isolate Shewanella algae B12 and Evaluation of Its Antimicrobial and Anti-biofilm Activity. Appl. Biochem. Biotechnol. 2022, 194, 1755–1774. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.L.; Berthomier, L.; Braganza, C.D.; MacKichan, J.K.; Ryan, J.L.; Visnovsky, G.; Keyzers, R.A. Identification, library synthesis and anti-vibriosis activity of 2-benzyl-4-chlorophenol from cultures of the marine bacterium Shewanella halifaxensis. Bioorg. Med. Chem. Lett. 2016, 26, 3086–3088. [Google Scholar] [CrossRef]
- Ezeoke, M.C.; Krishnan, P.; Sim, D.S.; Lim, S.H.; Low, Y.Y.; Chong, K.W.; Lim, K.H. Unusual phenethylamine-containing alkaloids from Elaeocarpus tectorius. Phytochemistry 2018, 146, 75–81. [Google Scholar] [CrossRef]
- Asakawa, D.; Sugiyama, E.; Mizuno, H.; Todoroki, K. Study of Substituted Phenethylamine Fragmentation Induced by Electrospray Ionization Mass Spectrometry and Its Application for Highly Sensitive Analysis of Neurotransmitters in Biological Samples. J. Am. Soc. Mass Spectrom. 2021, 32, 2144–2152. [Google Scholar] [CrossRef]
- Wang, L.; Marner, M.; Mettal, U.; Liu, Y.; Schäberle, T.F. Seven New Alkaloids Isolated from Marine Flavobacterium Tenacibaculum discolor sv11. Mar. Drugs 2022, 20, 620. [Google Scholar] [CrossRef]
- Lesimple, A.; Mamer, O.; Miao, W.; Chan, T.H. Electrospray mass spectral fragmentation study of N,N′-disubstituted imidazolium ionic liquids. J. Am. Soc. Mass Spectrom. 2006, 17, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, G.; Irwin, W.J. Mass spectra of some substituted imidazoles. Org. Mass Spectrom. 1975, 10, 885–895. [Google Scholar] [CrossRef]
- Bowie, J.; Cooks, R.; Lawesson, S.; Schroll, G. Electron impact studies. XII. Mass spectra of substituted imidazoles. Aust. J. Chem. 1967, 20, 1613–1624. [Google Scholar] [CrossRef]
- Wang, D.; Galla, H.J.; Drucker, P. Membrane interactions of ionic liquids and imidazolium salts. Biophys. Rev. 2018, 10, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Chiba, R.; Ito, M.; Nishio, Y. Addition effects of imidazolium salts on mesophase structure and optical properties of concentrated hydroxypropyl cellulose aqueous solutions. Polymer. J. 2010, 42, 232–241. [Google Scholar] [CrossRef]
- Modaressi, A.; Sifaoui, H.; Mielcarz, M.; Domańska, U.; Rogalski, M. Influence of the molecular structure on the aggregation of imidazolium ionic liquids in aqueous solutions. Colloid Surf. Physicochem. Eng Asp. 2007, 302, 181–185. [Google Scholar] [CrossRef]
- Buonocore, C.; Tedesco, P.; Vitale, G.A.; Palma Esposito, F.; Giugliano, R.; Monti, M.C.; D’Auria, M.V.; de Pascale, D. Characterization of a New Mixture of Mono-Rhamnolipids Produced by Pseudomonas gessardii Isolated from Edmonson Point (Antarctica). Mar. Drugs 2020, 18, 269. [Google Scholar] [CrossRef]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.-S.; Williamson, P.R.; Zheng, W. Improving therapy of severe infections through drug repurposing of synergistic combinations. Curr. Opin. Pharmacol. 2019, 48, 92–98. [Google Scholar] [CrossRef]
- Hendlin, D.; Stapley, E.O.; Jackson, M.; Wallick, H.; Miller, A.K.; Wolf, F.J.; Miller, T.W.; Chaiet, L.; Kahan, F.M.; Foltz, E.L.; et al. Phosphonomycin, a New Antibiotic Produced by Strains of Streptomyces. Science 1969, 166, 122–123. [Google Scholar] [CrossRef]
- Antonello, R.M.; Principe, L.; Maraolo, A.E.; Viaggi, V.; Pol, R.; Fabbiani, M.; Montagnani, F.; Lovecchio, A.; Luzzati, R.; Di Bella, S. Fosfomycin as Partner Drug for Systemic Infection Management. A Systematic Review of Its Synergistic Properties from In Vitro and In Vivo Studies. Antibiotics 2020, 9, 500. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Chen, P.-Y.; Wang, J.-T.; Chang, S.-C. A study on combination of daptomycin with selected antimicrobial agents: In vitro synergistic effect of MIC value of 1 mg/L against MRSA strains. BMC Pharmacol. Toxicol. 2019, 20, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingscheid, T.; Poeppl, W.; Bernitzky, D.; Veletzky, L.; Kussmann, M.; Plasenzotti, R.; Burgmann, H. Daptomycin plus fosfomycin, a synergistic combination in experimental implant-associated osteomyelitis due to methicillin-resistant Staphylococcus aureus in rats. Antimicrob. Agents Chemother. 2015, 59, 859–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kügler, J.H.; Le Roes-Hill, M.; Syldatk, C.; Hausmann, R. Surfactants tailored by the class Actinobacteria. Front Microbiol. 2015, 6, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhand, A.; Bayer, A.S.; Pogliano, J.; Yang, S.-J.; Bolaris, M.; Nizet, V.; Wang, G.; Sakoulas, G. Use of antistaphylococcal β-lactams to increase daptomycin activity in eradicating persistent bacteremia due to methicillin-resistant Staphylococcus aureus: Role of enhanced daptomycin binding. J. Clin. Infect. Dis. 2011, 53, 158–163. [Google Scholar] [CrossRef]
- Melander, R.J.; Liu, H.B.; Stephens, M.D.; Bewley, C.A.; Melander, C. Marine sponge alkaloids as a source of anti-bacterial adjuvants. Bioorg. Med. Chem. Lett. 2016, 26, 5863–5866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouzet, M.; Le Senechal, C.; Brözel, V.S.; Costaglioli, P.; Barthe, C.; Bonneu, M.; Garbay, B.; Vilain, S. Exploring early steps in biofilm formation: Set-up of an experimental system for molecular studies. BMC Microbiol. 2014, 14, 253. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Front. Microbiol. 2020, 11, 928. [Google Scholar] [CrossRef]
- Verderosa, A.D.; Totsika, M.; Fairfull-Smith, K.E. Bacterial Biofilm Eradication Agents: A Current Review. Front. Chem. 2019, 7, 824. [Google Scholar] [CrossRef] [Green Version]
- Seidah, N.G.; Pasquato, A.; Andréo, U. How Do Enveloped Viruses Exploit the Secretory Proprotein Convertases to Regulate Infectivity and Spread? Viruses 2021, 13, 1229. [Google Scholar] [CrossRef]
- Rathbun, M.M.; Szpara, M.L. A holistic perspective on herpes simplex virus (HSV) ecology and evolution. Adv. Virus Res. 2021, 110, 27–57. [Google Scholar] [CrossRef] [PubMed]
- Palma Esposito, F.; Giugliano, R.; Della Sala, G.; Vitale, G.A.; Buonocore, C.; Ausuri, J.; Galasso, C.; Coppola, D.; Franci, G.; Galdiero, M.; et al. Combining OSMAC Approach and Untargeted Metabolomics for the Identification of New Glycolipids with Potent Antiviral Activity Produced by a Marine Rhodococcus. Int. J. Mol. Sci. 2021, 22, 9055. [Google Scholar] [CrossRef] [PubMed]
- Connolly, S.A.; Jardetzky, T.S.; Longnecker, R. The structural basis of herpesvirus entry. Nat. Rev. Microbiol. 2021, 19, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Klasse, P.J.; Bron, R.; Marsh, M. Mechanisms of enveloped virus entry into animal cells. Adv. Drug Deliv. Rev. 1998, 34, 65–91. [Google Scholar] [CrossRef] [PubMed]
- Gong, K.K.; Tang, X.L.; Liu, Y.S.; Li, P.L.; Li, G.Q. Imidazole Alkaloids from the South China Sea Sponge Pericharax heteroraphis and Their Cytotoxic and Antiviral Activities. Molecules 2016, 21, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.X.; Zhou, Z.; Jiang, D.X.; Han, J.; Wang, J.F.; Zhao, L.W.; Li, J. In vivo anthelmintic activity of five alkaloids from Macleaya microcarpa (Maxim) Fedde against Dactylogyrus intermedius in Carassius auratus. Vet. Parasitol. 2010, 171, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.R.; Ropiak, H.M.; Fryganas, C.; Desrues, O.; Mueller-Harvey, I.; Thamsborg, S.M. Assessment of the anthelmintic activity of medicinal plant extracts and purified condensed tannins against free-living and parasitic stages of Oesophagostomum dentatum. Parasites Vectors 2014, 7, 518. [Google Scholar] [CrossRef]
- Hahnel, S.R.; Dilks, C.M.; Heisler, I.; Andersen, E.C.; Kulke, D. Caenorhabditis elegans in anthelmintic research—Old model, new perspectives. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 237–248. [Google Scholar] [CrossRef]
- Rocha, J.A.; Andrade, I.M.; Véras, L.M.; Quelemes, P.V.; Lima, D.F.; Soares, M.J.; Pinto, P.L.; Mayo, S.J.; Ivanova, G.; Rangel, M.; et al. Anthelmintic, Antibacterial and Cytotoxicity Activity of Imidazole Alkaloids from Pilocarpus microphyllus Leaves. Phytother. Res. 2017, 31, 624–630. [Google Scholar] [CrossRef] [Green Version]
- Tempone, A.G.; Pieper, P.; Borborema, S.E.T.; Thevenard, F.; Lago, J.H.G.; Croft, S.L.; Anderson, E.A. Marine alkaloids as bioactive agents against protozoal neglected tropical diseases and malaria. Nat. Prod. Rep. 2021, 38, 2214–2235. [Google Scholar] [CrossRef]
- Wolthers, K.C.; Susi, P.; Jochmans, D.; Koskinen, J.; Landt, O.; Sanchez, N.; Palm, K.; Neyts, J.; Butcher, S.J. Progress in human picornavirus research: New findings from the AIROPico consortium. Antivir. Res. 2019, 161, 100–107. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial sp. | Strain Code | Resistance Phenotype | Source |
|---|---|---|---|
| S. aureus | MSSA | Multisensitive | Eye |
| S. aureus | β-lactamase | Beta-lactamase producer | Wound |
| S. aureus | MLSB | Constitutive resistance to macrolides | Blood |
| S. aureus | QRSA | Quinolones resistance | Sputum |
| S. aureus | MRSA | Methicillin resistance | Blood |
| Compound | Rt | [M+H]+ | m/z |
|---|---|---|---|
| tyramine (1) | 1.1 | C8H12NO | 138.0915 |
| phenylethylamine (2) | 1.7 | C8H12N | 122.0965 |
| N-(4-hydroxyphenethyl)acetamide (5) | 5.7 | C10H14NO2 | 180.1022 |
| N-formyl-2-phenylethylamine (3) | 11.1 | C9H12NO | 150.0917 |
| N-acetyl-2-phenylethylamine (4) | 11.6 | C10H14NO | 164.1071 |
| shewamidine A (10) | 12.2 | C17H21N2O | 269.1656 |
| shewamidine B (11) | 12.9 | C18H23N2O | 283.1813 |
| N,N’-diphenethylformimidamide (7) | 14.6 | C17H21N2 | 253.1704 |
| shewamidine C (8) | 15.3 | C18H23N2 | 267.1861 |
| shewamidine D (12) | 15.4 | C23H25N2O | 345.1969 |
| shewamidine E (9) | 17.6 | C23H25N2 | 329.2019 |
| N-phenethylpentadecanamide (6) | 32.3 | C23H39NO | 346.3111 |
| Imidazolium PEA Alkaloids | ||||||||
|---|---|---|---|---|---|---|---|---|
| Compound | Rt (min.) | [M+H]+ | m/z | R1 | R2 | R3 | X | Y |
| shewazole A (13) a | 15.8 | C20H23N2 | 291.1862 | H | CH3 | H | H | H |
| bacillimidazole A (14) [26] | 16.5 | C21H25N2 | 305.2017 | H | CH3 | CH3 | H | H |
| shewazole B (15) | 17.1 | C22H27N2 | 319.2174 | CH3 | H | C2H5 | H | H |
| isoshewazole B (16) a | 17.5 | C22H27N2 | 319.2174 | H | C2H5 | CH3 | H | H |
| shewazole C (17) | 17.9 | C23H29N2 | 333.2333 | CH3 | C2H5 | CH3 | H | H |
| isoshewazole C (18) a | 18.5 | C23H29N2 | 333.2333 | C3H7 | CH3 | H | H | H |
| shewazole D (19) | 18.5 | C25H25N2 | 353.2018 | H | Ph | H | H | H |
| shewazole E (20) | 18.8 | C26H27N2 | 367.2178 | Ph | CH3 | H | H | H |
| shewazole F (21) | 18.9 | C24H31N2 | 347.2490 | CH3 | C3H7 | CH3 | H | H |
| isoshewazole F (22) a | 19.3 | C24H31N2 | 347.2490 | C4H9 | CH3 | H | H | H |
| shewazole G (23) | 19.4 | C27H29N2 | 381.2331 | Ph | H | C2H5 | H | H |
| shewazole H (24) a | 19.5 | C25H33N2 | 361.2645 | C4H9 | CH3 | CH3 | H | H |
| shewazole I (25) | 19.8 | C28H31N2 | 395.2488 | Ph | C2H5 | CH3 | H | H |
| isoshewazole I (26) | 20.0 | C28H31N2 | 395.2488 | Ph | C3H7 | H | H | H |
| shewazole J (27) | 21.6 | C33H33N2 | 457.2643 | Ph | Ph | C2H5 | H | H |
| shewazole K (28) | 21.8 | C31H37N2 | 437.2958 | C4H9 | Ph | C2H5 | H | H |
| Imidazolium TYM Alkaloids | ||||||||
| Compound | Rt (min.) | [M+H]+ | m/z | R1 | R2 | R3 | X | Y |
| hydroxyshewazole A (29) b | 13.5 | C20H23N2O | 307.1809 | H | CH3 | H | OH c | H |
| hydroxybacillimidazole A (30) b | 14.3 | C21H25N2O | 321.1967 | H | CH3 | CH3 | OH | H |
| hydroxyshewazole B (31) | 14.8 | C22H27N2O | 335.2122 | CH3 | H | C2H5 | OH | H |
| dihydroxyshewazole I (32) | 16.0 | C28H31N2O2 | 427.2388 | Ph | C2H5 | CH3 | OH | OH |
| hydroxyshewazole F (33) | 16.8 | C24H31N2O | 363.2438 | CH3 | C3H7 | CH3 | OH c | H |
| hydroxyshewazole H (34) a | 17.3 | C25H33N2O | 377.2596 | C4H9 | CH3 | CH3 | OH | H |
| hydroxyshewazole G (35) b | 17.4 | C27H29N2O | 397.2280 | Ph | H | C2H5 | OH c | H |
| hydroxyshewazole I (36) a | 17.9 | C28H31N2O | 411.2440 | Ph | C2H5 | CH3 | OH c | H |
| hydroxyshewazole J (37) b | 19.7 | C33H33N2O | 473.2593 | Ph | Ph | C2H5 | OH c | H |
| F2 Minimal Inhibitory and Bactericidal Concentrations (µg/mL) | ||
|---|---|---|
| S. aureus Strain | MIC | MBC |
| ATCC 6538 | 100 | 100 |
| MLSB | 100 | 200 |
| Quinolone res. | 100 | 200 |
| Β-lactamase | 100 | 200 |
| MSSA | 100 | 200 |
| MRSA | 100 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giugliano, R.; Della Sala, G.; Buonocore, C.; Zannella, C.; Tedesco, P.; Palma Esposito, F.; Ragozzino, C.; Chianese, A.; Morone, M.V.; Mazzella, V.; et al. New Imidazolium Alkaloids with Broad Spectrum of Action from the Marine Bacterium Shewanella aquimarina. Pharmaceutics 2023, 15, 2139. https://doi.org/10.3390/pharmaceutics15082139
Giugliano R, Della Sala G, Buonocore C, Zannella C, Tedesco P, Palma Esposito F, Ragozzino C, Chianese A, Morone MV, Mazzella V, et al. New Imidazolium Alkaloids with Broad Spectrum of Action from the Marine Bacterium Shewanella aquimarina. Pharmaceutics. 2023; 15(8):2139. https://doi.org/10.3390/pharmaceutics15082139
Chicago/Turabian StyleGiugliano, Rosa, Gerardo Della Sala, Carmine Buonocore, Carla Zannella, Pietro Tedesco, Fortunato Palma Esposito, Costanza Ragozzino, Annalisa Chianese, Maria Vittoria Morone, Valerio Mazzella, and et al. 2023. "New Imidazolium Alkaloids with Broad Spectrum of Action from the Marine Bacterium Shewanella aquimarina" Pharmaceutics 15, no. 8: 2139. https://doi.org/10.3390/pharmaceutics15082139