
Limosilactobacillus fermentum KAU0021 Abrogates Mono- and Polymicrobial Biofilms Formed by Candida albicans and Staphylococcus aureus

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Characterization of the Lactobacillus Strains
2.2. Pathogenic Strains and Growth Conditions
2.3. Preparation of Lactobacillus Metabolite Extract
2.4. Antimicrobial Activity of LMEs
2.5. Antimicrobial Activity of LMEs against Mixed Microbial Population
2.6. Assessment of Anti-Biofilm Activity of LMEs on Mono- and Polymicrobial Biofilms
2.7. Assessment of Microbial Cell Viability
2.8. Confocal Laser Scanning Microscopy (CLSM) of Mono- and Polymicrobial Biofilms
2.9. Scanning Electron Microscopy (SEM) of Mono- and Polymicrobial Biofilms
2.10. Assessment of Microbial Cell Membrane Integrity
2.11. Cytotoxicity Studies
2.12. GC-MS Analysis
2.13. Statistical Analysis
3. Results and Discussion
3.1. Lactobacillus Metabolite Extracts and Their Antimicrobial Activity against Planktonic Cells
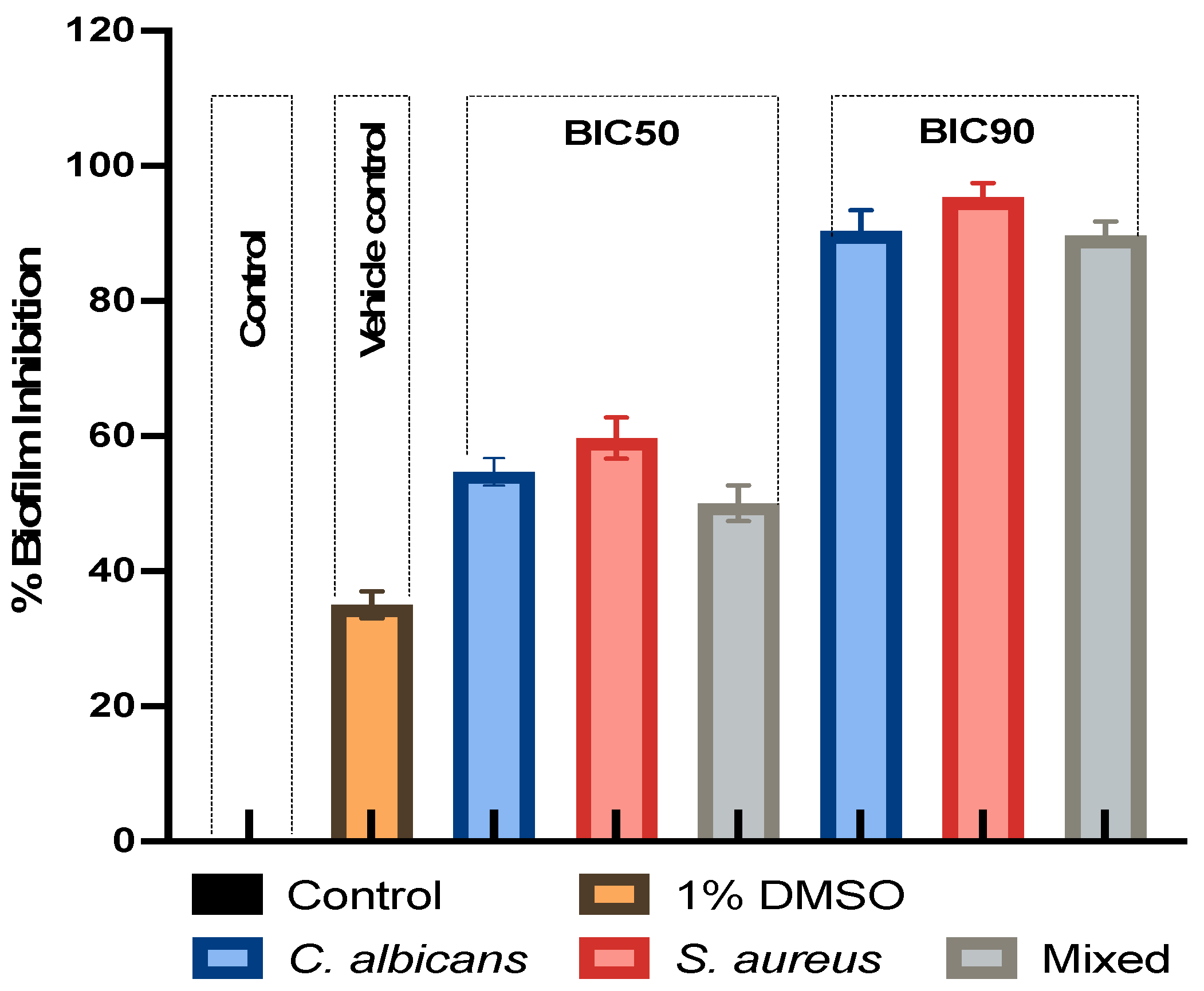
3.2. Effect of LMEKAU0021 on Biofilms
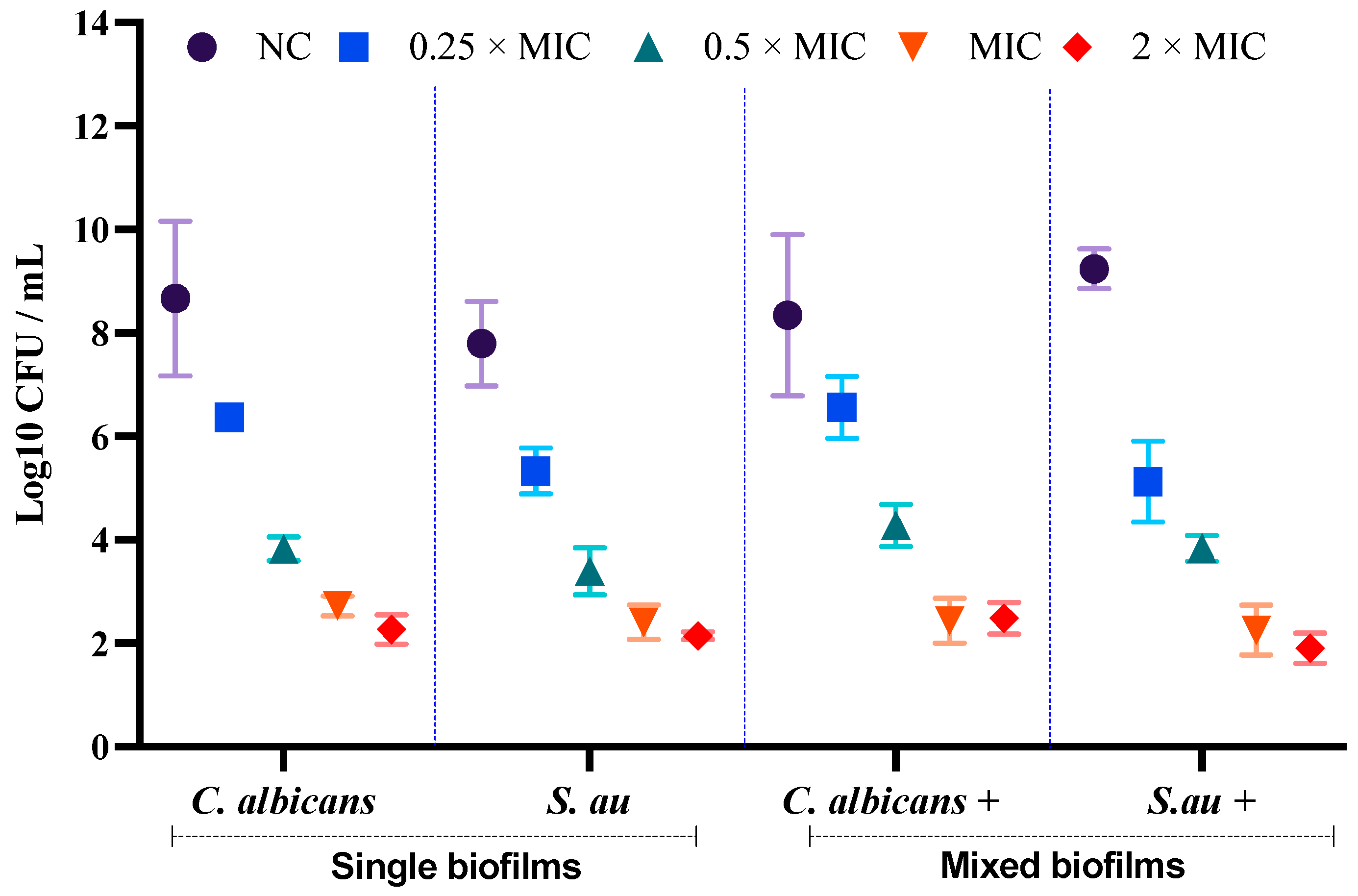
3.3. Assessment of Cell Viability of C. albicans and S. aureus in Single and Mixed Biofilms
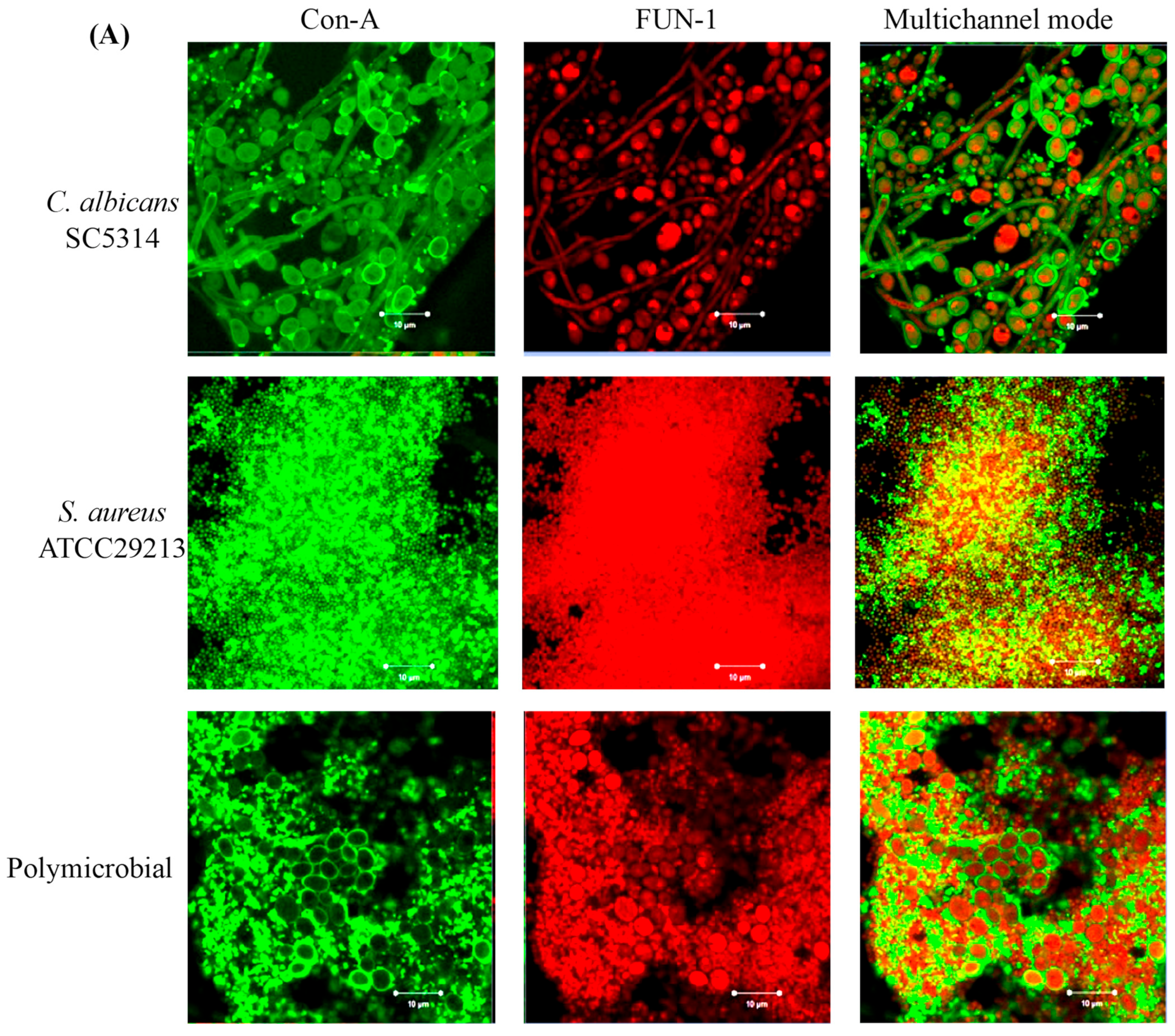
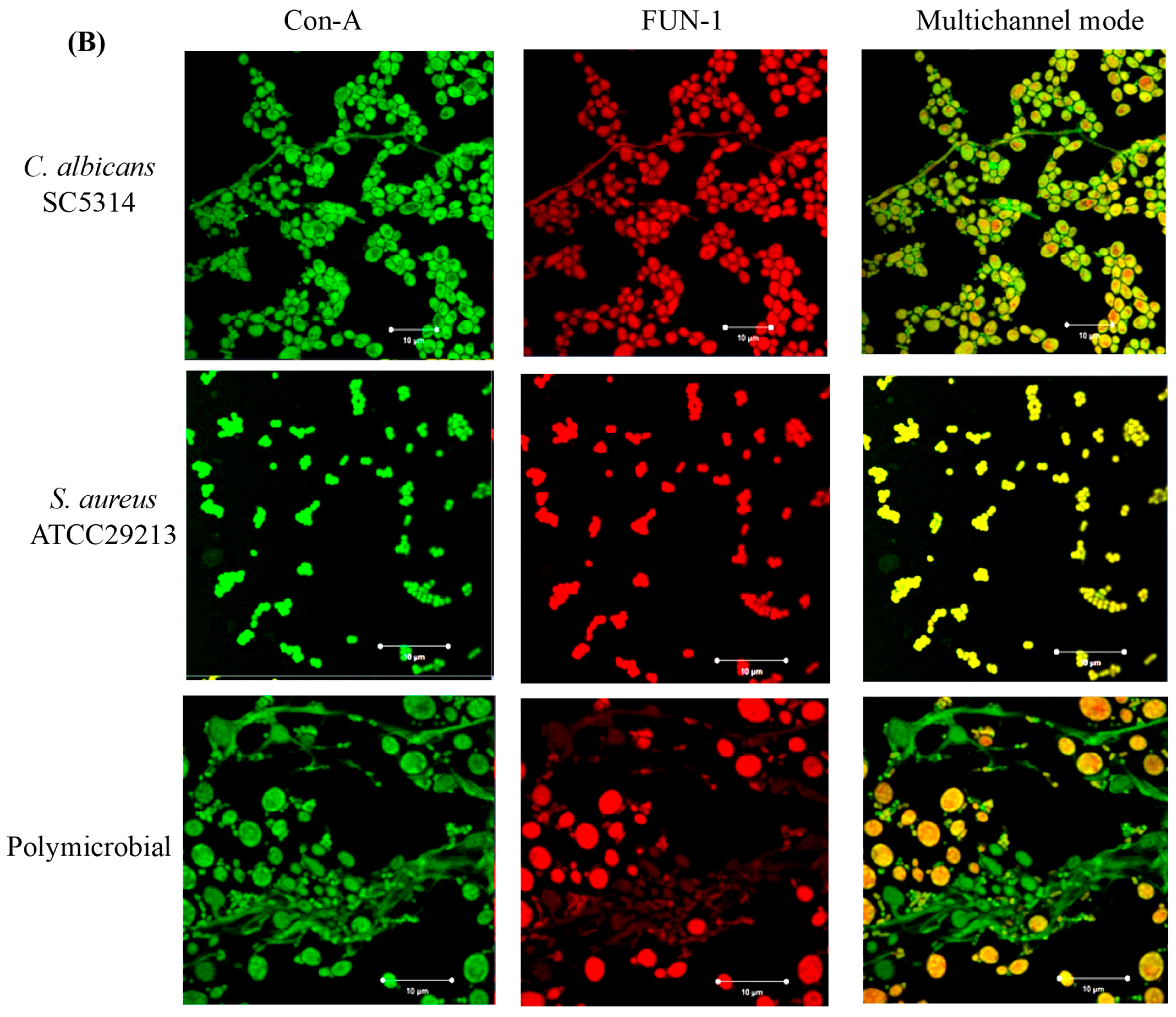
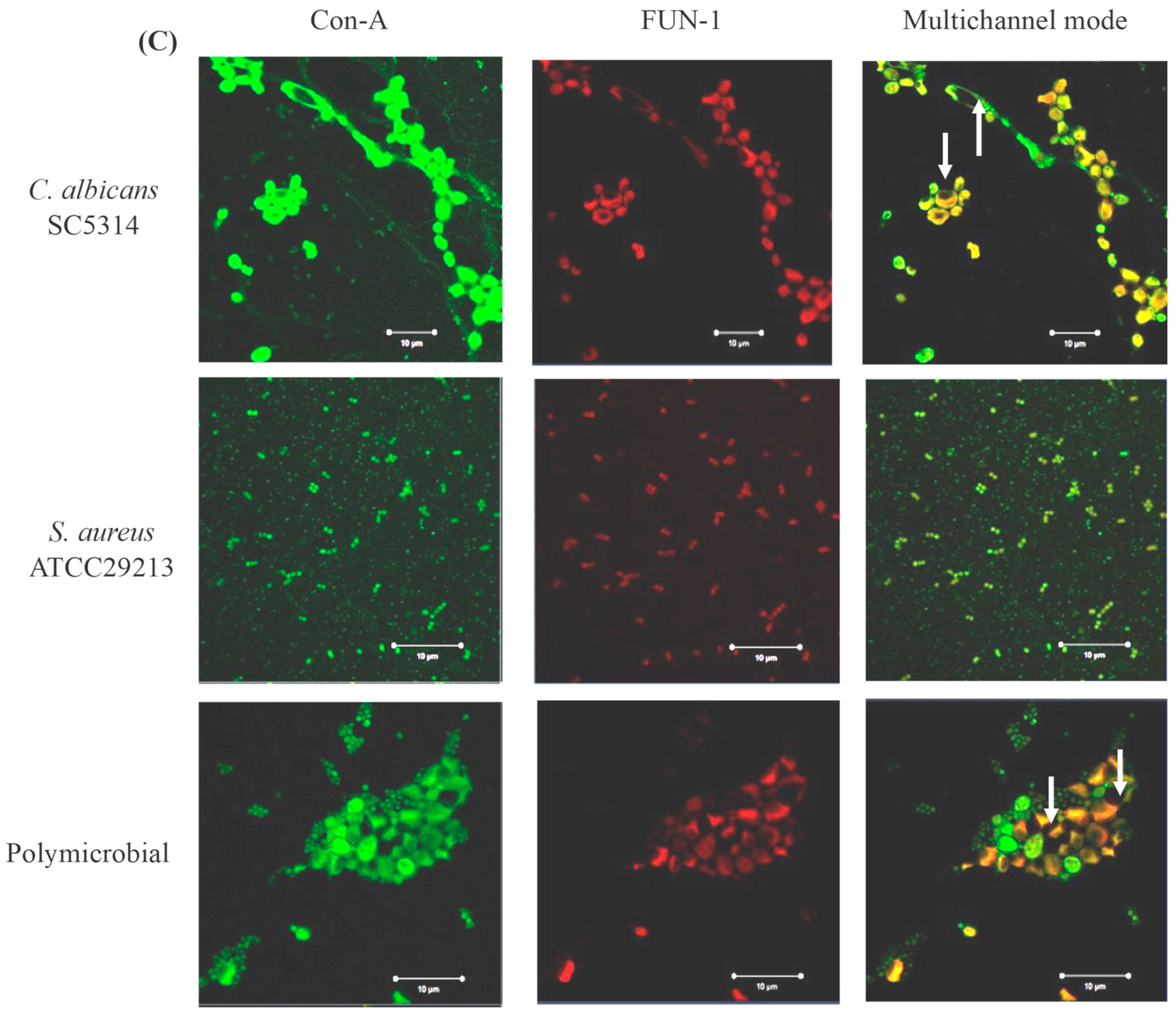
3.4. Confocal Laser Scanning Microscopy (CLSM) of Mono- and Polymicrobial Biofilms
3.5. SEM Analysis of Mono- and Polymicrobial Biofilms
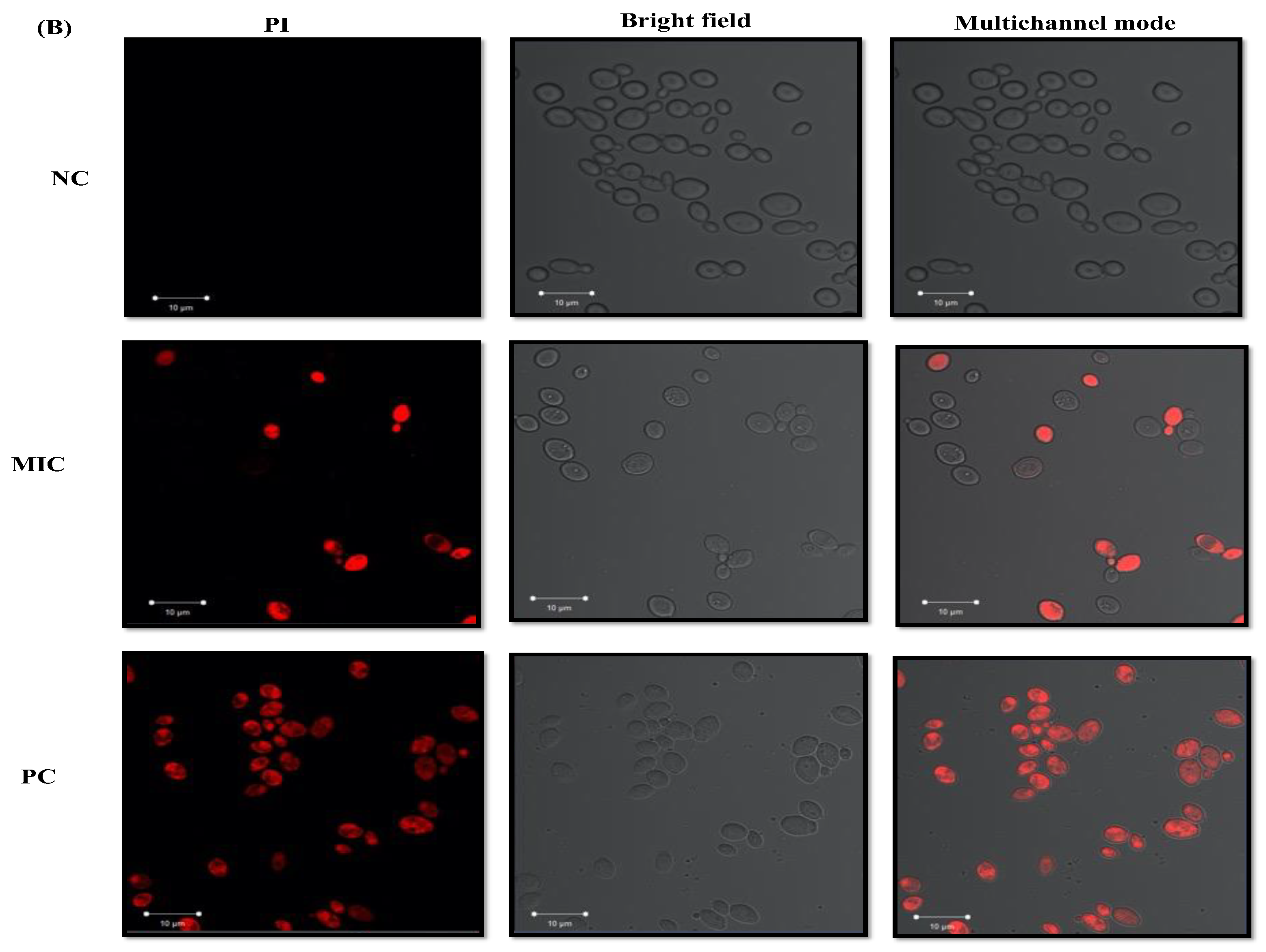
3.6. Assessment of Microbial Cell Membrane Integrity
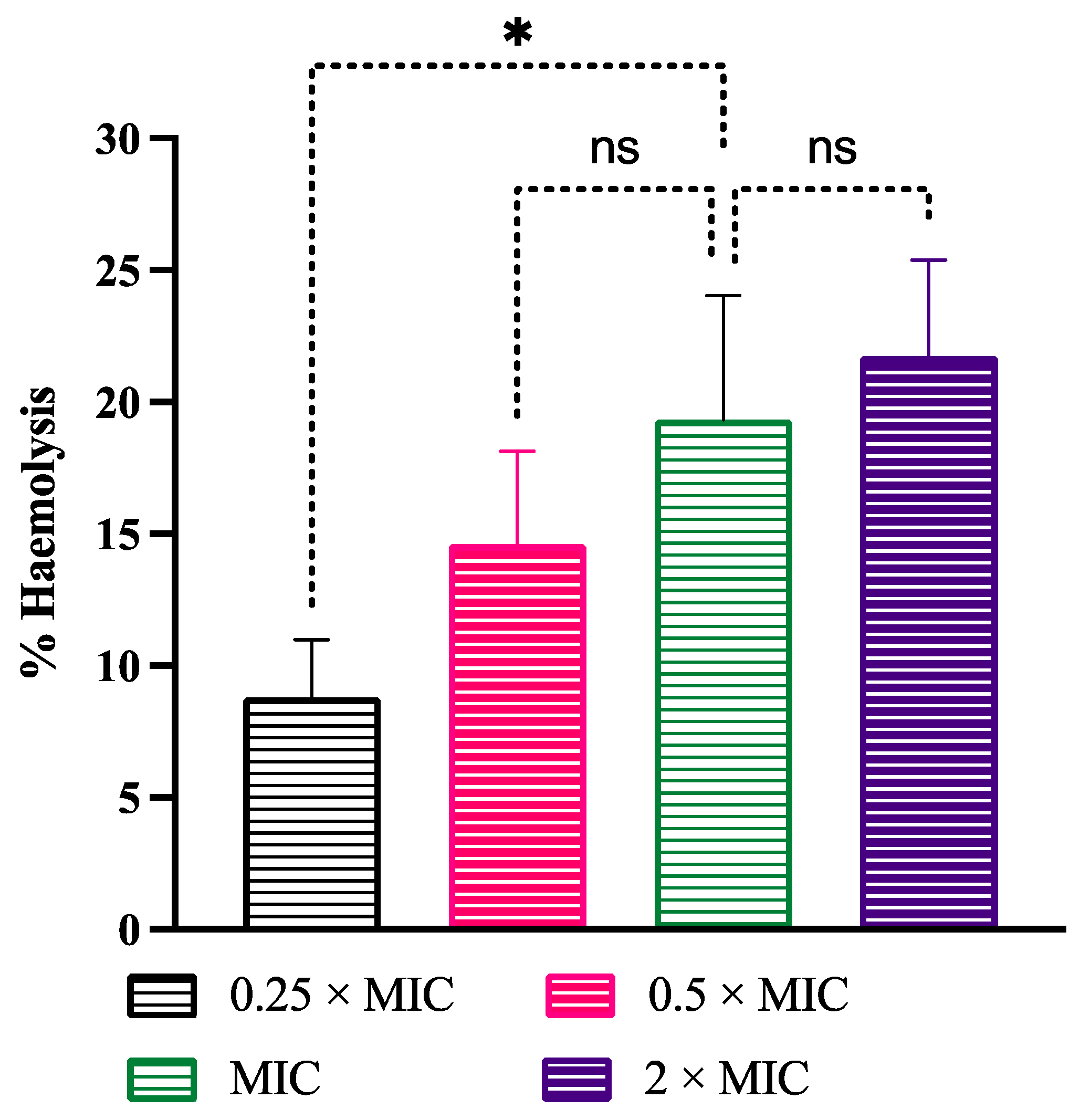
3.7. Cytotoxicity
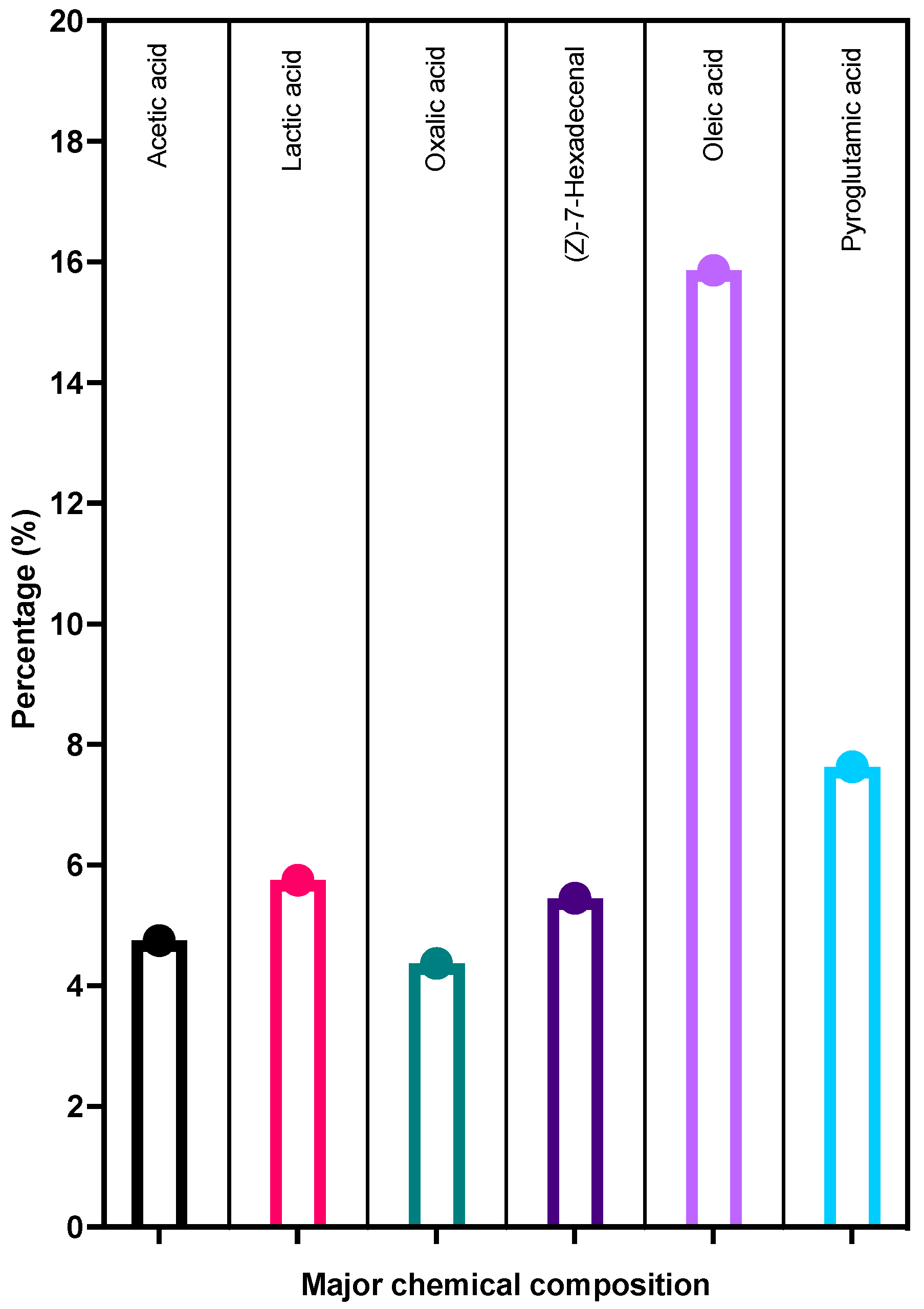
3.8. GC-MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carolus, H.; van Dyck, K.; van Dijck, P. Candida albicans and Staphylococcus Species: A Threatening Twosome. Front. Microbiol. 2019, 10, 2162. [Google Scholar] [CrossRef]
- Klotz, S.A.; Chasin, B.S.; Powell, B.; Gaur, N.K.; Lipke, P.N. Polymicrobial Bloodstream Infections Involving Candida Species: Analysis of Patients and Review of the Literature. Diagn. Microbiol. Infect. Dis. 2007, 59, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.; Jabra-Rizk, M.A.; Scheper, M.A.; Leid, J.G.; Costerton, J.W.; Shirtliff, M.E. Microbial Interactions and Differential Protein Expression in Staphylococcus Aureus -Candida Albicans Dual-Species Biofilms. FEMS Immunol. Med. Microbiol. 2010, 59, 493–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marra, A.R.; Camargo, L.F.A.; Pignatari, A.C.C.; Sukiennik, T.; Behar, P.R.P.; Medeiros, E.A.S.; Ribeiro, J.; Girão, E.; Correa, L.; Guerra, C.; et al. Nosocomial Bloodstream Infections in Brazilian Hospitals: Analysis of 2,563 Cases from a Prospective Nationwide Surveillance Study. J. Clin. Microbiol. 2011, 49, 1866–1871. [Google Scholar] [CrossRef] [Green Version]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans Biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Landini, P.; Antoniani, D.; Burgess, J.G.; Nijland, R. Molecular Mechanisms of Compounds Affecting Bacterial Biofilm Formation and Dispersal. Appl. Microbiol. Biotechnol. 2010, 86, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Moser, C.; Bassi, G.L.; Coenye, T.; Donelli, G.; Hall-Stoodley, L.; Holá, V.; Imbert, C.; Kirketerp-Møller, K.; et al. ESCMID* Guideline for the Diagnosis and Treatment of Biofilm Infections 2014. Clin. Microbiol. Infect. 2015, 21, S1–S25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef] [Green Version]
- Suresh, M.K.; Biswas, R.; Biswas, L. An Update on Recent Developments in the Prevention and Treatment of Staphylococcus aureus Biofilms. Int. J. Med. Microbiol. 2018, 309, 1–12. [Google Scholar] [CrossRef]
- Qu, Y.; Locock, K.; Verma-Gaur, J.; Hay, I.D.; Meagher, L.; Traven, A. Searching for New Strategies against Polymicrobial Biofilm Infections: Guanylated Polymethacrylates Kill Mixed Fungal/Bacterial Biofilms. J. Antimicrob. Chemother. 2016, 71, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Carr, F.J.; Chill, D.; Maida, N. The Lactic Acid Bacteria: A Literature Survey. Crit. Rev. Microbiol. 2002, 28, 281–370. [Google Scholar] [CrossRef]
- Ruíz, F.O.; Gerbaldo, G.; García, M.J.; Giordano, W.; Pascual, L.; Barberis, I.L. Synergistic Effect between Two Bacteriocin-like Inhibitory Substances Produced by Lactobacilli Strains with Inhibitory Activity for Streptococcus agalactiae. Curr. Microbiol. 2012, 64, 349–356. [Google Scholar] [CrossRef]
- Madhu, A.N.; Prapulla, S.G. Evaluation and Functional Characterization of a Biosurfactant Produced by Lactobacillus plantarum CFR 2194. Appl. Biochem. Biotechnol. 2014, 172, 1777–1789. [Google Scholar] [CrossRef]
- Sharma, D.; Saharan, B.S. Functional Characterization of Biomedical Potential of Biosurfactant Produced by Lactobacillus helveticus. Biotechnol. Rep. 2016, 11, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Fuochi, V.; Cardile, V.; Petronio Petronio, G.; Furneri, P.M. Biological Properties and Production of Bacteriocins-like-Inhibitory Substances by Lactobacillus Sp. Strains from Human Vagina. J. Appl. Microbiol. 2019, 126, 1541–1550. [Google Scholar] [CrossRef]
- Chen, C.-C.; Lai, C.-C.; Huang, H.-L.; Huang, W.-Y.; Toh, H.-S.; Weng, T.-C.; Chuang, Y.-C.; Lu, Y.-C.; Tang, H.-J. Antimicrobial Activity of Lactobacillus Species Against Carbapenem-Resistant Enterobacteriaceae. Front. Microbiol. 2019, 10, 789. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, R.; Yocheva, L.; Tserovska, L.; Zhelezova, G.; Stefanova, N.; Atanasova, A.; Danguleva, A.; Ivanova, G.; Karapetkov, N.; Rumyan, N.; et al. Antimicrobial activity and antibiotic susceptibility of Lactobacillus and Bifidobacterium spp. intended for use as starter and probiotic cultures. Biotechnol. Biotechnol. Equip. 2015, 29, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Jamalifar, H.; Rahimi, H.; Samadi, N.; Shahverdi, A.; Sharifian, Z.; Hosseini, F.; Eslahi, H. Antimicrobial activity of different Lactobacillus species against multi- drug resistant clinical isolates of Pseudomonas aeruginosa. Iran. J. Microbiol. 2011, 3, 21–25. [Google Scholar]
- Gautam, N.; Sharma, N. Purification and Characterization of Bacteriocin Produced by Strain of Lactobacillus brevis MTCC 7539. Indian J. Biochem. Biophys. 2009, 46, 337–341. [Google Scholar]
- Hata, T.; Tanaka, R.; Ohmomo, S. Isolation and Characterization of Plantaricin ASM1: A New Bacteriocin Produced by Lactobacillus Plantarum A-1. Int. J. Food Microbiol. 2010, 137, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, L.; Hao, Y.; Zhong, S.; Liu, H.; Han, T.; Xie, Y. Isolation and Partial Characterization of a Bacteriocin Produced by Lactobacillus plantarum BM-1 Isolated from a Traditionally Fermented Chinese Meat Product. Microbiol. Immunol. 2013, 57, 746–755. [Google Scholar] [CrossRef]
- Ge, J.; Sun, Y.; Xin, X.; Wang, Y.; Ping, W. Purification and Partial Characterization of a Novel Bacteriocin Synthesized by Lactobacillus paracasei HD1-7 Isolated from Chinese Sauerkraut Juice. Sci. Rep. 2016, 6, srep19366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rather, I.A.; Kamli, M.R.; Sabir, J.S.M.; Paray, B.A. Potential Antiviral Activity of Lactiplantibacillus plantarum KAU007 against Influenza Virus H1N1. Vaccines 2022, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Al Malki, A.; Yoon, S.-H.; Firoz, A.; Ali, H.M.; Park, Y.-H.; Hor, Y.-Y.; Rather, I.A. Characterization of New Probiotic Isolates from Fermented Ajwa Dates of Madinah and Their Anti-Inflammatory Potential. Appl. Sci. 2022, 12, 5082. [Google Scholar] [CrossRef]
- Alzahrani, O.R.; Hawsawi, Y.M.; Alanazi, A.D.; Alatwi, H.E.; Rather, I.A. In Vitro Evaluation of Leuconostoc mesenteroides Cell-Free-Supernatant GBUT-21 against SARS-CoV-2. Vaccines 2022, 10, 1581. [Google Scholar] [CrossRef]
- Badwaik, L.S.; Borah, P.K.; Deka, S.C. Production and Purification of Anti-Bacterial Biometabolite from Wild-Type Lactobacillus, Isolated from Fermented Bamboo Shoot: Future Suggestions and a Proposed System for Secondary Metabolite Onsite Recovery During Continuous Fermentation. Appl. Biochem. Biotechnol. 2015, 175, 1915–1925. [Google Scholar] [CrossRef]
- Fernandes, L.; Fortes, B.N.; Lincopan, N.; Ishida, K. Caspofungin and Polymyxin B Reduce the Cell Viability and Total Biomass of Mixed Biofilms of Carbapenem-Resistant Pseudomonas aeruginosa and Candida spp. Front. Microbiol. 2020, 11, 573263. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, K.; Morinaga, Y.; Nakamura, S.; Seki, M.; Izumikawa, K.; Kakeya, H.; Yamamoto, Y.; Yamada, Y.; Kamihira, S.; Kohno, S.; et al. Subinhibitory concentrations of telithromycin, clarithromycin and azithromycin reduce methicillin-resistant Staphylococcus aureus coagulase in vitro and in vivo. J. Antimicrob. Chemother. 2008, 61, 647–650. [Google Scholar] [CrossRef] [Green Version]
- Clancy, C.J.; Yu, V.L.; Morris, A.J.; Snydman, D.; Nguyen, M.H. Fluconazole MIC and the fluconazole dose/MIC ratio correlate with therapeutic response among patients with candidemia. Antimicrob. Agents Chemother. 2005, 49, 3171–3177. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, C.; Liu, P.; Liu, W.; Gao, Y.; Sun, S. In Vitro Interactions between Fluconazole and Minocycline against Mixed Cultures of Candida albicans and Staphylococcus aureus. J. Microbiol. Immunol. Infect. 2015, 48, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Scheunemann, G.; Fortes, B.N.; Lincopan, N.; Ishida, K. Caspofungin Inhibits Mixed Biofilms of Candida albicans and Methicillin-Resistant Staphylococcus aureus and Displays Effectiveness in Coinfected Galleria Mellonella Larvae. Microbiol. Spectr. 2021, 9, e00744-21. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.E.; Gomes, F.; Rodrigues, C.F. Candida spp./Bacteria Mixed Biofilms. J. Fungi 2020, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.; Dubey, A.K. Anti-Biofilm Activity of the Metabolites of Streptomyces chrestomyceticus Strain ADP4 against Candida Albicans. J. Biosci. Bioeng. 2016, 122, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Ahmad, A. Abrogation of Pathogenic Attributes in Drug Resistant Candida auris Strains by Farnesol. PLoS ONE 2020, 15, e0233102. [Google Scholar] [CrossRef]
- Suchodolski, J.; Muraszko, J.; Bernat, P.; Krasowska, A. Lactate like Fluconazole Reduces Ergosterol Content in the Plasma Membrane and Synergistically Kills Candida albicans. Int. J. Mol. Sci. 2021, 22, 5219. [Google Scholar] [CrossRef] [PubMed]
- Lone, S.A.; Ahmad, A. Inhibitory Effect of Novel Eugenol Tosylate Congeners on Pathogenicity of Candida albicans. BMC Complement. Med. Ther. 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Rather, I.A.; Wani, M.Y.; Kamli, M.R.; Sabir, J.S.; Hakeem, K.R.; Firoz, A.; Park, Y.H.; Hor, Y.Y. Lactiplantibacillus plantarum KAU007 Extract Modulates Critical Virulence Attributes and Biofilm Formation in Sinusitis Causing Streptococcus Pyogenes. Pharmaceutics 2022, 14, 2702. [Google Scholar] [CrossRef]
- Park, C.W.; Youn, M.; Jung, Y.M.; Kim, H.; Jeong, Y.; Lee, H.K.; Kim, H.O.; Lee, I.; Lee, S.W.; Kang, K.H.; et al. New Functional Probiotic Lactobacillus sakei Probio 65 Alleviates Atopic Symptoms in the Mouse. J. Med. Food 2008, 11, 405–412. [Google Scholar] [CrossRef]
- Kim, H.; Rather, I.A.; Kim, H.; Kim, S.; Kim, T.; Jang, J.; Seo, J.; Lim, J.; Park, Y.-H. A Double-Blind, Placebo Controlled-Trial of a Probiotic Strain Lactobacillus sakei Probio-65 for the Prevention of Canine Atopic Dermatitis. J. Microbiol. Biotechnol. 2015, 25, 1966–1969. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Rather, I.A.; Park, Y.-H. Partially Purified Exo-Polysaccharide from Lactobacillus sakei Probio 65 with Antioxidant, α-Glucosidase and Tyrosinase Inhibitory Potential. J. Food Biochem. 2016, 40, 264–274. [Google Scholar] [CrossRef]
- Rather, I.A.; Bajpai, V.K.; Huh, Y.S.; Han, Y.-K.; Bhat, E.A.; Lim, J.; Paek, W.K.; Park, Y.-H. Probiotic Lactobacillus sakei ProBio-65 Extract Ameliorates the Severity of Imiquimod Induced Psoriasis-like Skin Inflammation in a Mouse Model. Front. Microbiol. 2018, 9, 1021. [Google Scholar] [CrossRef] [Green Version]
- Rather, I.A.; Bajpai, V.K.; Ching, L.L.; Majumder, R.; Nam, G.-J.; Indugu, N.; Singh, P.; Kumar, S.; Hajrah, N.H.; Sabir, J.S.M.; et al. Effect of a Bioactive Product SEL001 from Lactobacillus sakei Probio65 on Gut Microbiota and Its Anti-Colitis Effects in a TNBS-Induced Colitis Mouse Model. Saudi J. Biol. Sci. 2020, 27, 261–270. [Google Scholar] [CrossRef]
- Burianek, L.L.; Yousef, A.E. Solvent Extraction of Bacteriocins from Liquid Cultures. Lett. Appl. Microbiol. 2000, 31, 193–197. [Google Scholar] [CrossRef]
- Jafri, H.; Banerjee, G.; Khan, M.S.A.; Ahmad, I.; Abulreesh, H.H.; Althubiani, A.S. Synergistic Interaction of Eugenol and Antimicrobial Drugs in Eradication of Single and Mixed Biofilms of Candida albicans and Streptococcus mutans. AMB Express 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Rather, A.I.; Seo, B.J.; Rejish, K.V.J.; Choi, U.H.; Choi, K.H.; Lim, J.H.; Park, Y.H. Isolation and Characterization of a Proteinaceous Antifungal Aompound from Lactobacillus plantarum YML007 and its Application as a Food Preservative. Lett. Appl. Microbiol. 2013, 57, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Rushdy, A.A.; Gomaa, E.Z. Antimicrobial Compounds Produced by Probiotic Lactobacillus brevis Isolated from Dairy Products. Ann. Microbiol. 2013, 63, 81–90. [Google Scholar] [CrossRef]
- Fijan, S. Probiotics and their Antimicrobial Effect. Microorganisms 2023, 11, 528. [Google Scholar] [CrossRef]
- Karami, S.; Roayaei, M.; Zahedi, E.; Bahmani, M.; Mahmoodnia, L.; Hamzavi, H.; Rafieian-Kopaei, M. Antifungal Effecats of Lactobacillus Species Isolated from Local Dairy Products. Int. J. Pharm. Investig. 2017, 7, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; McAuley, D.F.; Fulton, C.R.; Pessoa, J.S.; McMullan, R.; Lundy, F.T. Targeting Candida albicans in Dual-Species Biofilms with Antifungal Treatment Reduces Staphylococcus aureus and MRSA in Vitro. PLoS ONE 2021, 16, e0249547. [Google Scholar] [CrossRef] [PubMed]
- Lara, H.H.; Lopez-Ribot, J.L. Inhibition of Mixed Biofilms of Candida albicans and Methicillin-Resistant Staphylococcus aureus by Positively Charged Silver Nanoparticles and Functionalized Silicone Elastomers. Pathogens 2020, 9, 784. [Google Scholar] [CrossRef]
- Staib, F.; Grosse, G.; Mishra, S.K. Staphylococcus aureus and Candida albicans Infection (Animal Experiments). Zbl.Bakt. Reihe A 1976, 234, 450–461. [Google Scholar]
- Vila, T.; Kong, E.F.; Ibrahim, A.; Piepenbrink, K.; Shetty, A.C.; McCracken, C.; Bruno, V.; Jabra-Rizk, M.A. Candida albicans Quorum-Sensing Molecule Farnesol Modulates Staphyloxanthin Production and Activates the Thiol-Based Oxidative-Stress Response in Staphylococcus aureus. Virulence 2019, 10, 625–642. [Google Scholar] [CrossRef] [Green Version]
- Todd, O.A.; Fidel, P.L.; Harro, J.M.; Hilliard, J.J.; Tkaczyk, C.; Sellman, B.R.; Noverr, M.C.; Peters, B.M. Candida albicans Augments Staphylococcus aureus Virulence by Engaging the Staphylococcal Agr Quorum Sensing System. Mbio 2019, 10, e00910-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.L.; Lee, R.T.H.; Fang, H.M.; Wang, Y.M.; Li, R.; Zou, H.; Zhu, Y.; Wang, Y. Bacterial Peptidoglycan Triggers Candida albicans Hyphal Growth by Directly Activating the Adenylyl Cyclase Cyr1p. Cell Host Microbe 2008, 4, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.J.; Alsad, L.; Vogel, F.; Koppar, S.; Nevarez, L.; Auguste, F.; Seymour, J.; Syed, A.; Christoph, K.; Loomis, J.S. Interactions between Candida albicans and Staphylococcus aureus within Mixed Species Biofilms. Bios 2013, 84, 30–39. [Google Scholar] [CrossRef]
- Spoering, A.L.; Lewis, K. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J. Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafri, H.; Khan, M.S.A.; Ahmad, I. In Vitro Efficacy of Eugenol in Inhibiting Single and Mixed-Biofilms of Drug-Resistant Strains of Candida albicans and Streptococcus mutans. Phytomedicine 2019, 54, 206–213. [Google Scholar] [CrossRef]
- Millard, P.J.; Roth, B.L.; Thi, H.P.T.; Yue, S.T.; Haugland, R.P. Development of the FUN-1 Family of Fluorescent Probes for Vacuole Labeling and Viability Testing of Yeasts. Appl. Environ. Microbiol. 1997, 63, 2897–2905. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, V.; Singla, R.K.; Dubey, A.K. Inhibition of Biofilm and Virulence Factors of Candida albicans by Partially Purified Secondary Metabolites of Streptomyces chrestomyceticus Strain ADP4. Curr. Top. Med. Chem. 2018, 18, 925–945. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Z.; Du, Q.; Li, P.; Wang, Z.; Wang, A. Stimulated Phase-Shift Acoustic Nanodroplets Enhance Vancomycin Efficacy against Methicillin-Resistant Staphylococcus aureus Biofilms. Int. J. Nanomed. 2017, 12, 4679–4690. [Google Scholar] [CrossRef] [Green Version]
- Ponde, N.O.; Lortal, L.; Ramage, G.; Naglik, J.R.; Richardson, J.P. Candida albicans Biofilms and Polymicrobial Interactions. Crit. Rev. Microbiol. 2021, 47, 91–111. [Google Scholar] [CrossRef]
- Setiawati, S.; Nuryastuti, T.; Ngatidjan, N.; Mustofa, M.; Jumina, J.; Fitriastuti, D. In Vitro Antifungal Activity of (1)-N-2-Methoxybenzyl-1,10-Phenanthrolinium Bromide against Candida albicans and Its Effects on Membrane Integrity. Mycobiology 2017, 45, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, A.; Kueffer, M.; Simmen, A.; Grand, M. Relatedness of Lactobacillus Rhamnosus Strains Isolated from Clinical Specimens and Such from Food-Stuffs, Humans and Technology. LWT 1998, 31, 489–494. [Google Scholar] [CrossRef]
- Santini, C.; Baffoni, L.; Gaggia, F.; Granata, M.; Gasbarri, R.; di Gioia, D.; Biavati, B. Characterization of Probiotic Strains: An Application as Feed Additives in Poultry against Campylobacter jejuni. Int. J. Food Microbiol. 2010, 141, S98–S108. [Google Scholar] [CrossRef] [PubMed]
- Maragkoudakis, P.A.; Zoumpopoulou, G.; Miaris, C.; Kalantzopoulos, G.; Pot, B.; Tsakalidou, E. Probiotic Potential of Lactobacillus Strains Isolated from Dairy Products. Int. Dairy J. 2006, 16, 189–199. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogens | LMEKAU0021 (μg/mL) | LMEKAU0010 (μg/mL) | LMEKAU007 (μg/mL) | LMEPro−65 (μg/mL) | ||||
|---|---|---|---|---|---|---|---|---|
| MIC | MFC/MBC | MIC | MFC/MBC | MIC | MFC/MBC | MIC | MFC/MBC | |
| C. albicans | 406 | 812 | 406 | 812 | 1624 | 3248 | 1624 | >3248 |
| S. aureus | 203 | 406 | 406 | 812 | 1624 | 3248 | 1624 | >3248 |
| Polymicrobial | 406 | 812 | 406 | 1624 | 3248 | >3248 | 1624 | >3248 |
| LMEKAU0021 | Biofilm Formation | 24 h Preformed Biofilm | ||||
|---|---|---|---|---|---|---|
| C. albicans Alone | S. aureus Alone | Mixed Biofilms | C. albicans Alone | S. aureus Alone | Mixed Biofilms | |
| BIC50 | 101.5 μg/mL | 50.75 μg/mL | 101.5 μg/mL | 203 μg/mL | 101.5 μg/mL | 203 μg/mL |
| BIC90 | 203 μg/mL | 101.5 μg/mL | 203 μg/mL | 406 μg/mL | 203 μg/mL | 406 μg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rather, I.A.; Wani, M.Y.; Kamli, M.R.; Sabir, J.S.M.; Hakeem, K.R.; Firoz, A.; Park, Y.H.; Hor, Y.Y. Limosilactobacillus fermentum KAU0021 Abrogates Mono- and Polymicrobial Biofilms Formed by Candida albicans and Staphylococcus aureus. Pharmaceutics 2023, 15, 1079. https://doi.org/10.3390/pharmaceutics15041079
Rather IA, Wani MY, Kamli MR, Sabir JSM, Hakeem KR, Firoz A, Park YH, Hor YY. Limosilactobacillus fermentum KAU0021 Abrogates Mono- and Polymicrobial Biofilms Formed by Candida albicans and Staphylococcus aureus. Pharmaceutics. 2023; 15(4):1079. https://doi.org/10.3390/pharmaceutics15041079
Chicago/Turabian StyleRather, Irfan A., Mohmmad Younus Wani, Majid Rasool Kamli, Jamal S. M. Sabir, Khalid Rehman Hakeem, Ahmad Firoz, Yong Ha Park, and Yan Yan Hor. 2023. "Limosilactobacillus fermentum KAU0021 Abrogates Mono- and Polymicrobial Biofilms Formed by Candida albicans and Staphylococcus aureus" Pharmaceutics 15, no. 4: 1079. https://doi.org/10.3390/pharmaceutics15041079