
The Antibiofilm Effects of Antimony Tin Oxide Nanoparticles against Polymicrobial Biofilms of Uropathogenic Escherichia coli and Staphylococcus aureus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Microbial Culture
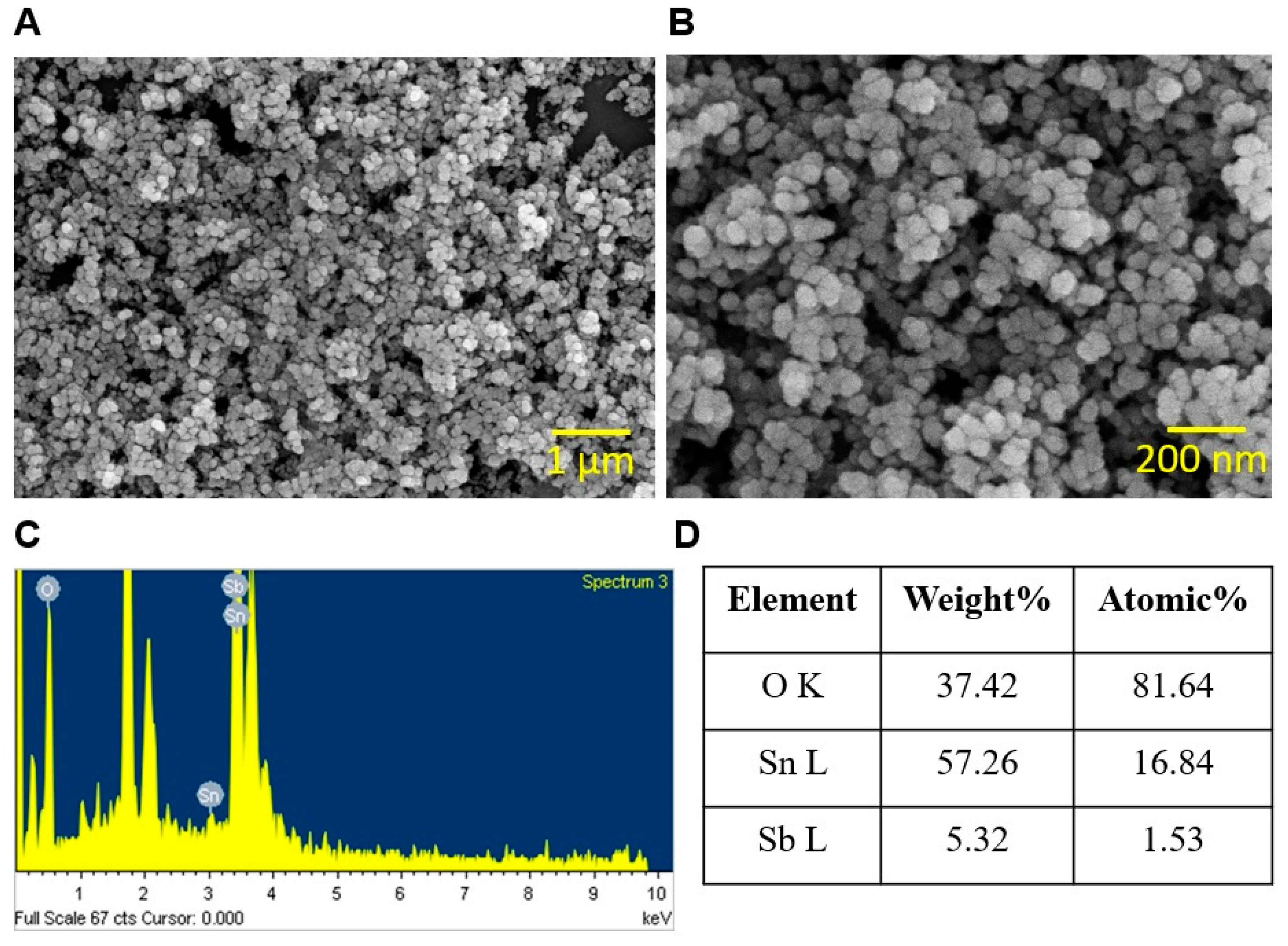
2.2. Characterization of ATO NPs with SEM, X-ray Diffraction, TEM, DLS, and Zeta-Potential Analyzer
2.3. Crystal Violet Assay in 96-Well Polystyrene Plates
2.4. Microscopic Imaging of Biofilms
2.5. Cell Surface Hydrophobicity (CSH)
2.6. Sheep Blood Hemolysis Assay
2.7. Slime Production by S. aureus
2.8. Gene Expression Study with qRT PCR
2.9. C. elegans Toxicity Study
2.10. Effect of ATO NPs on Plant Brassica campestris Growth
2.11. Statistical Analysis
3. Results
3.1. Characterization of ATO Nanoparticles
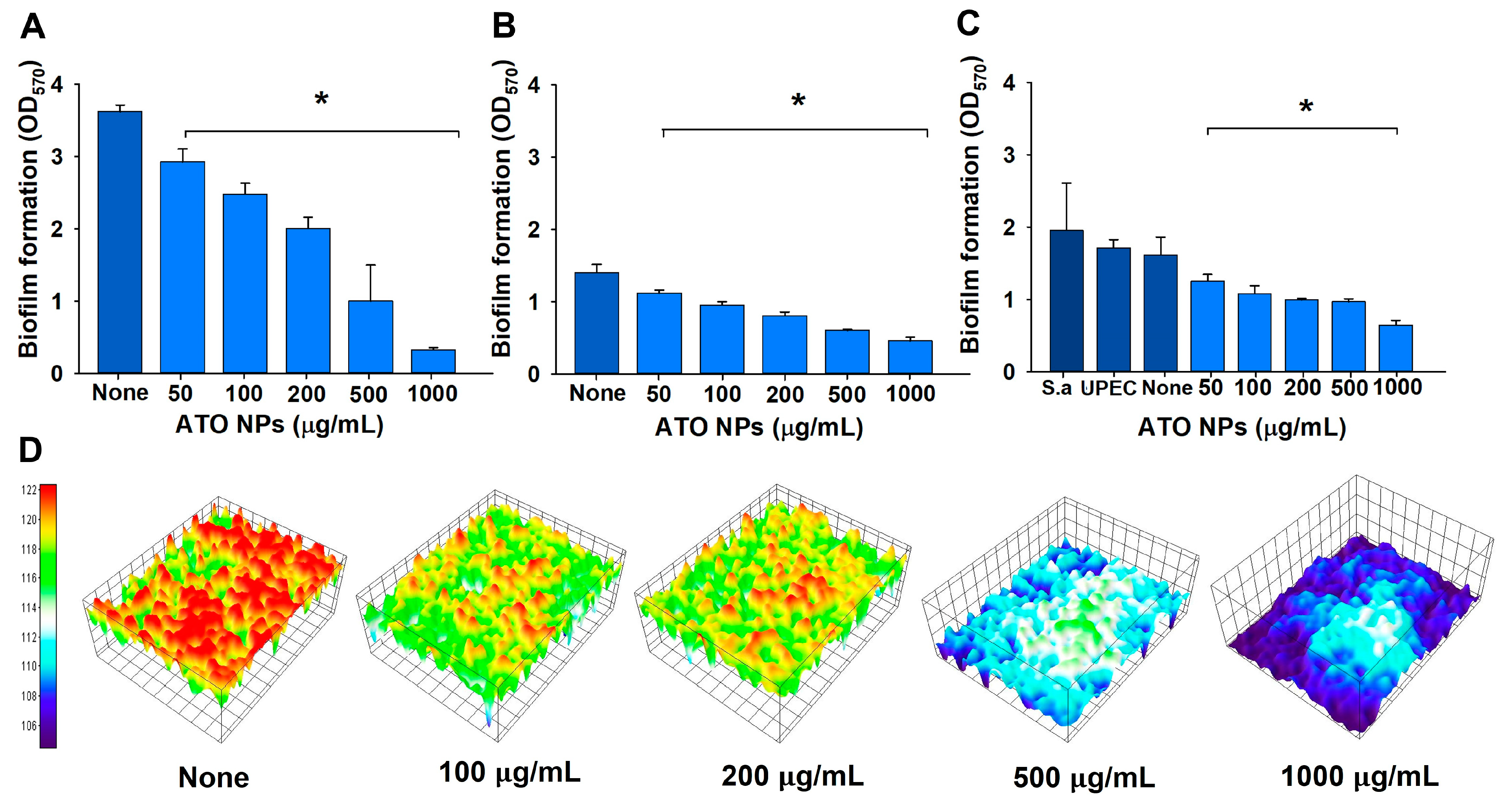
3.2. Inhibition of Single- and Polymicrobial-Biofilm Formation by ATO NPs
3.3. Visualization of Biofilms after ATO NP Treatment
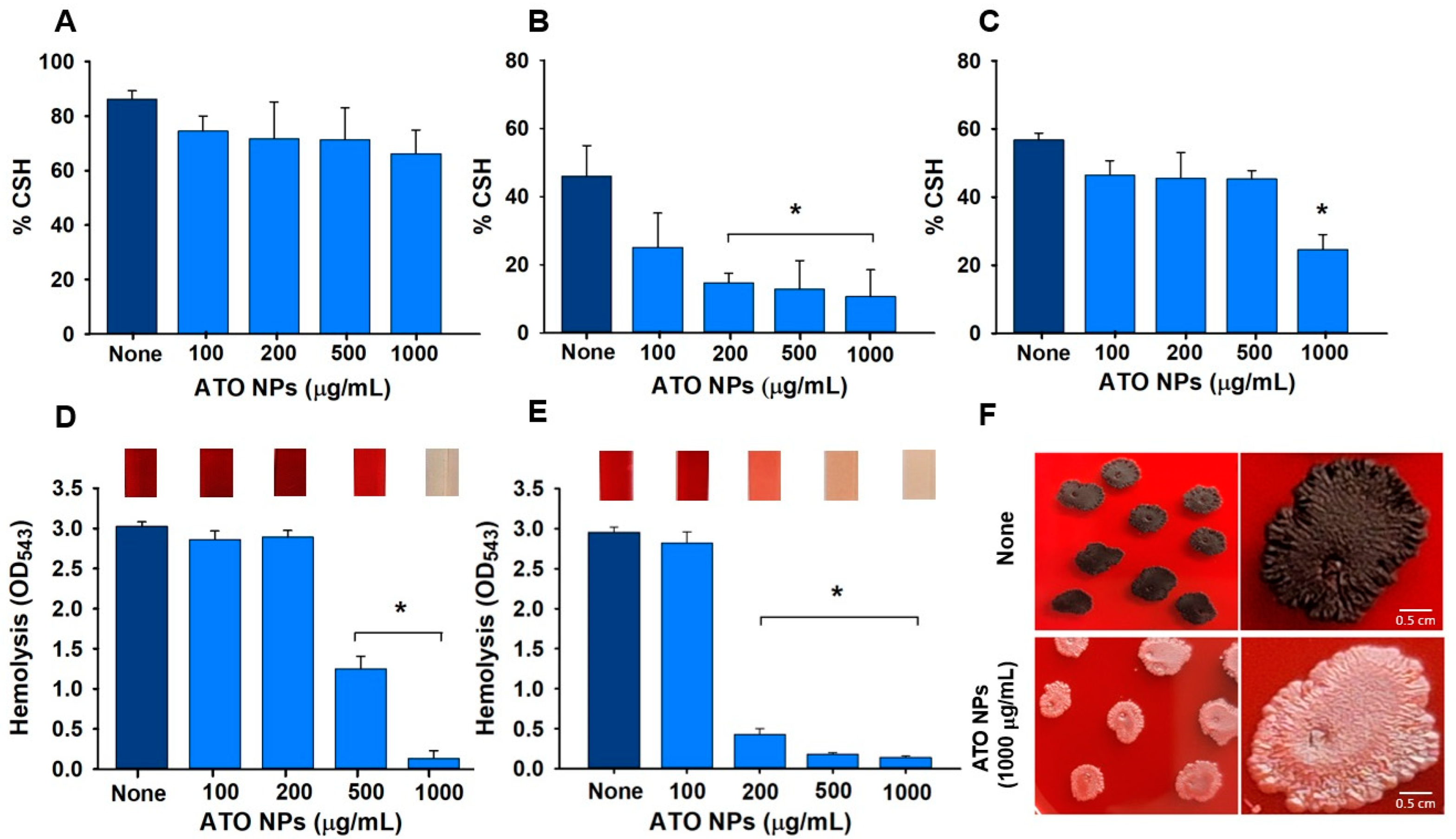
3.4. Effects of ATO NPs on CSH
3.5. Impact of ATO NPs on Hemolysis
3.6. Slime Production by S. aureus
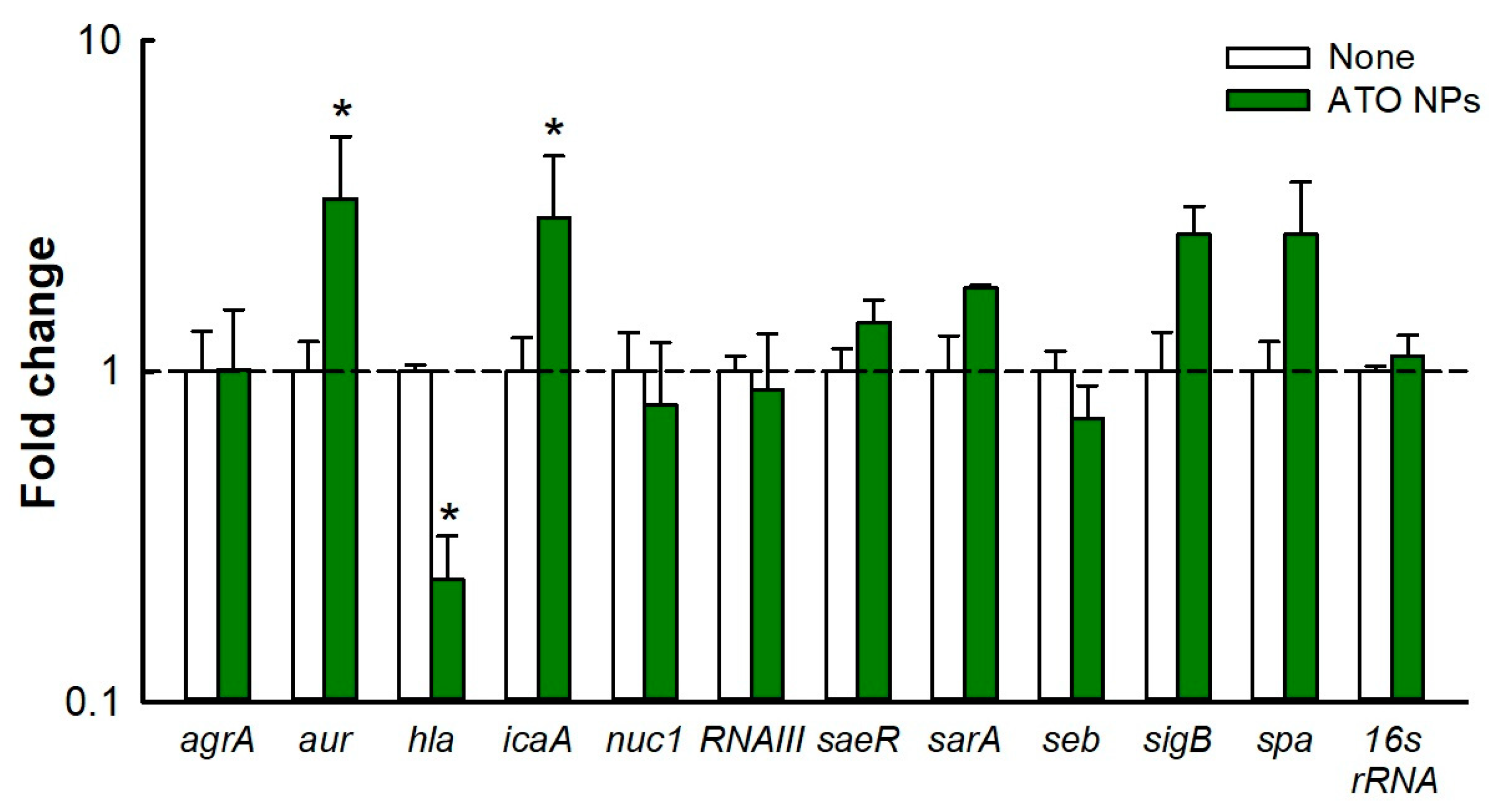
3.7. Gene Expression Analysis
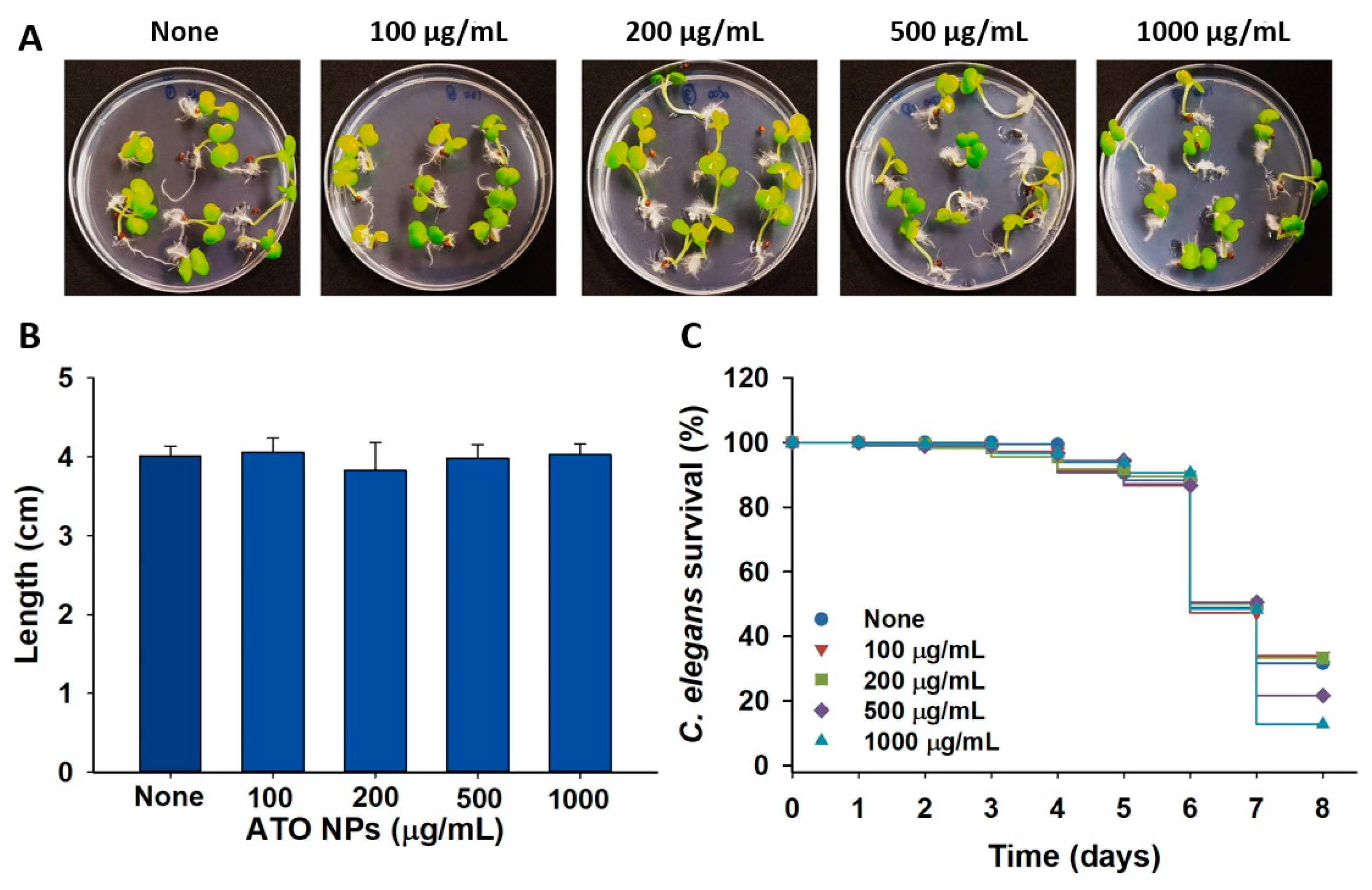
3.8. Toxic Effects of ATO NPs on Plant Growth and Nematode Survival
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harriott, M.M.; Noverr, M.C. Importance of Candida-bacterial polymicrobial biofilms in disease. Trends Microbiol. 2011, 19, 557–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigoryan, L.; Trautner, B.W.; Gupta, K. Diagnosis and management of urinary tract infections in the outpatient setting: A review. JAMA 2014, 312, 1677–1684. [Google Scholar] [CrossRef]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate point-prevalence survey of health care–associated infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) infections: Virulence factors, bladder responses, antibiotic, and non-antibiotic antimicrobial strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, R.J.; Monroe, P.W. Bacteremic urinary tract infection in older people. J. Am. Geriatr. Soc. 1996, 44, 927–933. [Google Scholar] [CrossRef]
- Yousefi, M.; Pourmand, M.R.; Fallah, F.; Hashemi, A.; Mashhadi, R.; Nazari-Alam, A. Characterization of Staphylococcus aureus biofilm formation in urinary tract infection. Iran. J. Public. Health 2016, 45, 485–493. [Google Scholar] [PubMed]
- Walker, J.N.; Flores-Mireles, A.L.; Pinkner, C.L.; Schreiber, H.L., IV; Joens, M.S.; Park, A.M.; Potretzke, A.M.; Bauman, T.M.; Pinkner, J.S.; Fitzpatrick, J.A.J.; et al. Catheterization alters bladder ecology to potentiate Staphylococcus aureus infection of the urinary tract. Proc. Natl. Acad. Sci. USA 2017, 114, E8721–E8730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Mahdy, R.; Mahmoud, R.; Shrief, R. Characterization of E. coli phylogroups causing catheter-associated urinary tract infection. Infect. Drug Resist. 2021, 14, 3183–3193. [Google Scholar] [CrossRef]
- Rather, I.A.; Wani, M.Y.; Kamli, M.R.; Sabir, J.S.; Hakeem, K.R.; Firoz, A.; Park, Y.H.; Hor, Y.Y. Limosilactobacillus fermentum KAU0021 abrogates mono- and polymicrobial biofilms formed by Candida albicans and Staphylococcus aureus. Pharmaceutics 2023, 15, 1079. [Google Scholar] [CrossRef]
- Sycz, Z.; Wojnicz, D.; Tichaczek-Goska, D. Does secondary plant metabolite ursolic acid exhibit antibacterial activity against Uropathogenic Escherichia coli living in single- and multispecies biofilms? Pharmaceutics 2022, 14, 1691. [Google Scholar] [CrossRef]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavi, M.; Varma, R.S. Phytosynthesis and modification of metal and metal oxide nanoparticles/nanocomposites for antibacterial and anticancer activities: Recent advances. Sustain. Chem. Pharm. 2021, 21, 100412. [Google Scholar] [CrossRef]
- Ramasamy, M.; Lee, J. Recent nanotechnology approaches for prevention and treatment of biofilm-associated infections on medical devices. BioMed Res. Int. 2016, 2016, 1851242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stankic, S.; Suman, S.; Haque, F.; Vidic, J. Pure and multi metal oxide nanoparticles: Synthesis, antibacterial and cytotoxic properties. J. Nanobiotechnol. 2016, 14, 73. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, R.M.; Habeeb, M.A.; Hashim, A. Effect of antimony oxide nanoparticles on structural, optical and AC electrical properties of (PEO-PVA) blend for antibacterial applications. Int. J. Emerg. Trends Eng. Res. 2020, 8, 4726–4738. [Google Scholar] [CrossRef]
- Sagadevan, S.; Lett, J.A.; Fatimah, I.; Lokanathan, Y.; Léonard, E.; Oh, W.C.; Hossain, M.M.; Johan, M.R. Current trends in the green syntheses of tin oxide nanoparticles and their biomedical applications. Mater. Res. Express 2021, 8, 082001. [Google Scholar] [CrossRef]
- Beyer, H.; Metzger, M.; Sicklinger, J.; Wu, X.; Schwenke, K.U.; Gasteiger, H.A. Antimony doped tin oxide–synthesis, characterization and application as cathode material in Li-O2 cells: Implications on the prospect of carbon-free cathodes for rechargeable lithium-air batteries. J. Electrochem. Soc. 2017, 164, A1026. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.; Li, J.; Yang, F.; Cao, K.; Bao, Q.; Sun, Y.; Yuan, J. Antimony-doped tin oxide nanocrystals for enhanced photothermal theragnosis therapy of cancers. Front. Bioeng. Biotechnol. 2020, 8, 673. [Google Scholar] [CrossRef]
- Amutha, E.; Rajaduraipandian, S.; Sivakavinesan, M.; Annadurai, G. Hydrothermal synthesis and characterization of the antimony–tin oxide nanomaterial and its application as a high-performance asymmetric supercapacitor, photocatalyst, and antibacterial agent. Nanoscale Adv. 2023, 5, 255–267. [Google Scholar] [CrossRef]
- Lee, J.-H.; Park, J.H.; Cho, H.S.; Joo, S.W.; Cho, M.H.; Lee, J. Anti-biofilm activities of quercetin and tannic acid against Staphylococcus aureus. Biofouling 2013, 29, 491–499. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, S.; Cho, K.H.; Lee, J.-H.; Lee, J. Antibiofilm activities of cinnamaldehyde analogs against uropathogenic Escherichia coli and Staphylococcus aureus. Int. J. Mol. Sci. 2022, 23, 7225. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Jailani, A.; Lee, J.-H.; Lee, J. Effect of halogenated indoles on biofilm formation, virulence, and root surface colonization by Agrobacterium tumefaciens. Chemosphere 2022, 293, 133603. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Kim, Y.-G.; Ryu, S.Y.; Lee, J. Calcium-chelating alizarin and other anthraquinones inhibit biofilm formation and the hemolytic activity of Staphylococcus aureus. Sci. Rep. 2016, 6, 19267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-H.; Kim, Y.-G.; Khadke, S.K.; Lee, J. Antibiofilm and antifungal activities of medium-chain fatty acids against Candida albicans via mimicking of the quorum-sensing molecule farnesol. Microb. Biotechnol. 2021, 14, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Caiazza, N.C.; O’Toole, G.A. Alpha-toxin is required for biofilm formation by Staphylococcus aureus. J. Bacteriol. 2003, 185, 3214–3217. [Google Scholar] [CrossRef] [Green Version]
- Kannappan, A.; Gowrishankar, S.; Srinivasan, R.; Pandian, S.K.; Ravi, A.V. Antibiofilm activity of Vetiveria zizanioides root extract against methicillin-resistant Staphylococcus aureus. Microb. Pathog. 2017, 110, 313–324. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Parham, S.; Wicaksono, D.H.; Bagherbaigi, S.; Lee, S.L.; Nur, H. Antimicrobial treatment of different metal oxide nanoparticles: A critical review. J. Chin. Chem. Soc. 2016, 63, 385–393. [Google Scholar] [CrossRef]
- Reygaert, W.C. An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 2018, 4, 482–501. [Google Scholar] [CrossRef]
- Joseph, R.; Kaizerman, D.; Herzog, I.M.; Hadar, M.; Feldman, M.; Fridman, M.; Cohen, Y. Phosphonium pillar [5] arenes as a new class of efficient biofilm inhibitors: Importance of charge cooperativity and the pillar platform. Chem. Commun. 2016, 52, 10656–10659. [Google Scholar] [CrossRef]
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle-biofilm interactions: The role of the EPS matrix. Trends Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Ceri, H.; Stremick, C.A.; Turner, R.J. Biofilm susceptibility to metal toxicity. Environ. Microbiol. 2004, 6, 1220–1227. [Google Scholar] [CrossRef]
- An, Y.J.; Kim, M. Effect of antimony on the microbial growth and the activities of soil enzymes. Chemosphere 2009, 74, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Mitik-Dineva, N.; Wang, J.; Truong, V.K.; Stoddart, P.; Malherbe, F.; Crawford, R.J.; Ivanova, E.P. Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus attachment patterns on glass surfaces with nanoscale roughness. Curr. Microbiol. 2009, 58, 268–273. [Google Scholar] [CrossRef]
- Habimana, O.; Steenkeste, K.; Fontaine-Aupart, M.P.; Bellon-Fontaine, M.N.; Kulakauskas, S.; Briandet, R. Diffusion of nanoparticles in biofilms is altered by bacterial cell wall hydrophobicity. Appl. Environ. Microbiol. 2011, 77, 367–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Qi, Y.; Guo, S.; Hao, K.; Zhao, M.; Guo, N. Effect of AgWPA nanoparticles on the inhibition of Staphylococcus aureus growth in biofilms. Food Control 2019, 100, 240–246. [Google Scholar] [CrossRef]
- Lee, K.; Lee, J.-H.; Ryu, S.Y.; Cho, M.H.; Lee, J. Stilbenes reduce Staphylococcus aureus hemolysis, biofilm formation, and virulence. Foodborne Pathog. Dis. 2014, 11, 710–717. [Google Scholar] [CrossRef]
- Saghalli, M.; Bidoki, S.K.; Jamali, A.; Bagheri, H.; Ghaemi, E.A. Sub-minimum inhibitory concentrations of zinc oxide nanoparticles reduce the expression of the Staphylococcus aureus alpha-hemolysin. Indian J. Pharm. Sci. 2017, 78, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Pacheco-Blandino, I.; Vanner, R.; Buzea, C. Toxicity of nanoparticles. In Toxicity of Building Materials; Woodhead Publishing: Sawston, UK, 2012; pp. 427–475. [Google Scholar] [CrossRef]
- Sundar, S.; Chakravarty, J. Antimony Toxicity. Int. J. Environ. Res. Public Health 2010, 7, 4267–4277. [Google Scholar] [CrossRef]
- Bessa, M.J.; Brandão, F.; Fokkens, P.; Cassee, F.R.; Salmatonidis, A.; Viana, M.; Vulpoi, A.; Simon, S.; Monfort, E.; Teixeira, J.P.; et al. Toxicity assessment of industrial engineered and airborne process-generated nanoparticles in a 3D human airway epithelial in vitro model. Nanotoxicology 2021, 15, 542–557. [Google Scholar] [CrossRef]
- Rajendran, N.K.; Kumar, S.S.D.; Houreld, N.N.; Abrahamse, H. A review on nanoparticle based treatment for wound healing. J. Drug Deliv. Sci. Technol. 2018, 44, 421–430. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, I.; Jailani, A.; Lee, J.-H.; Ahmed, B.; Lee, J. The Antibiofilm Effects of Antimony Tin Oxide Nanoparticles against Polymicrobial Biofilms of Uropathogenic Escherichia coli and Staphylococcus aureus. Pharmaceutics 2023, 15, 1679. https://doi.org/10.3390/pharmaceutics15061679
Park I, Jailani A, Lee J-H, Ahmed B, Lee J. The Antibiofilm Effects of Antimony Tin Oxide Nanoparticles against Polymicrobial Biofilms of Uropathogenic Escherichia coli and Staphylococcus aureus. Pharmaceutics. 2023; 15(6):1679. https://doi.org/10.3390/pharmaceutics15061679
Chicago/Turabian StylePark, Inji, Afreen Jailani, Jin-Hyung Lee, Bilal Ahmed, and Jintae Lee. 2023. "The Antibiofilm Effects of Antimony Tin Oxide Nanoparticles against Polymicrobial Biofilms of Uropathogenic Escherichia coli and Staphylococcus aureus" Pharmaceutics 15, no. 6: 1679. https://doi.org/10.3390/pharmaceutics15061679