

The Exosome-Mediated PI3K/Akt/mTOR Signaling Pathway in Neurological Diseases


 ,
,  , and
, and
Abstract
:1. Introduction
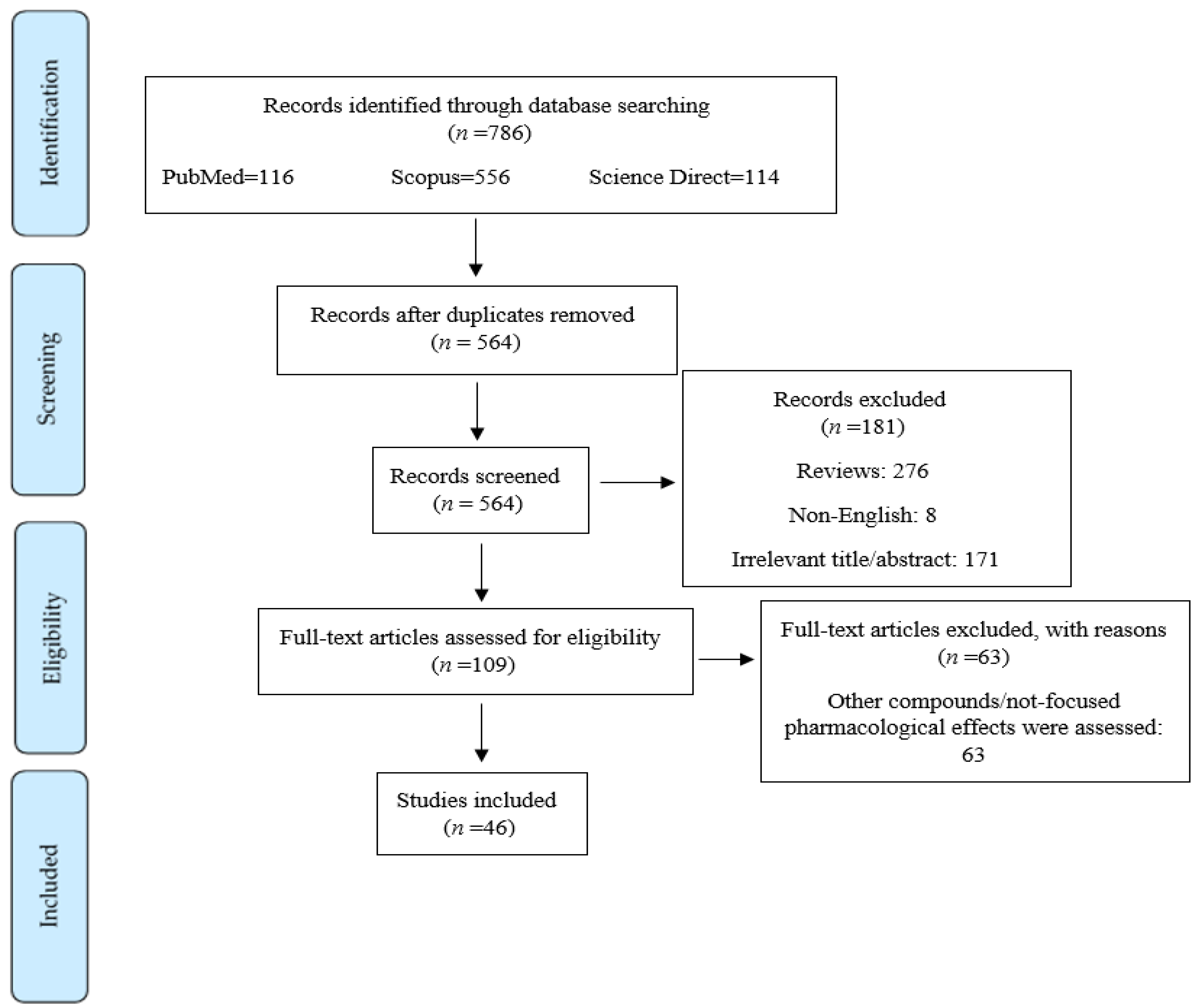
2. Study Design and Methods
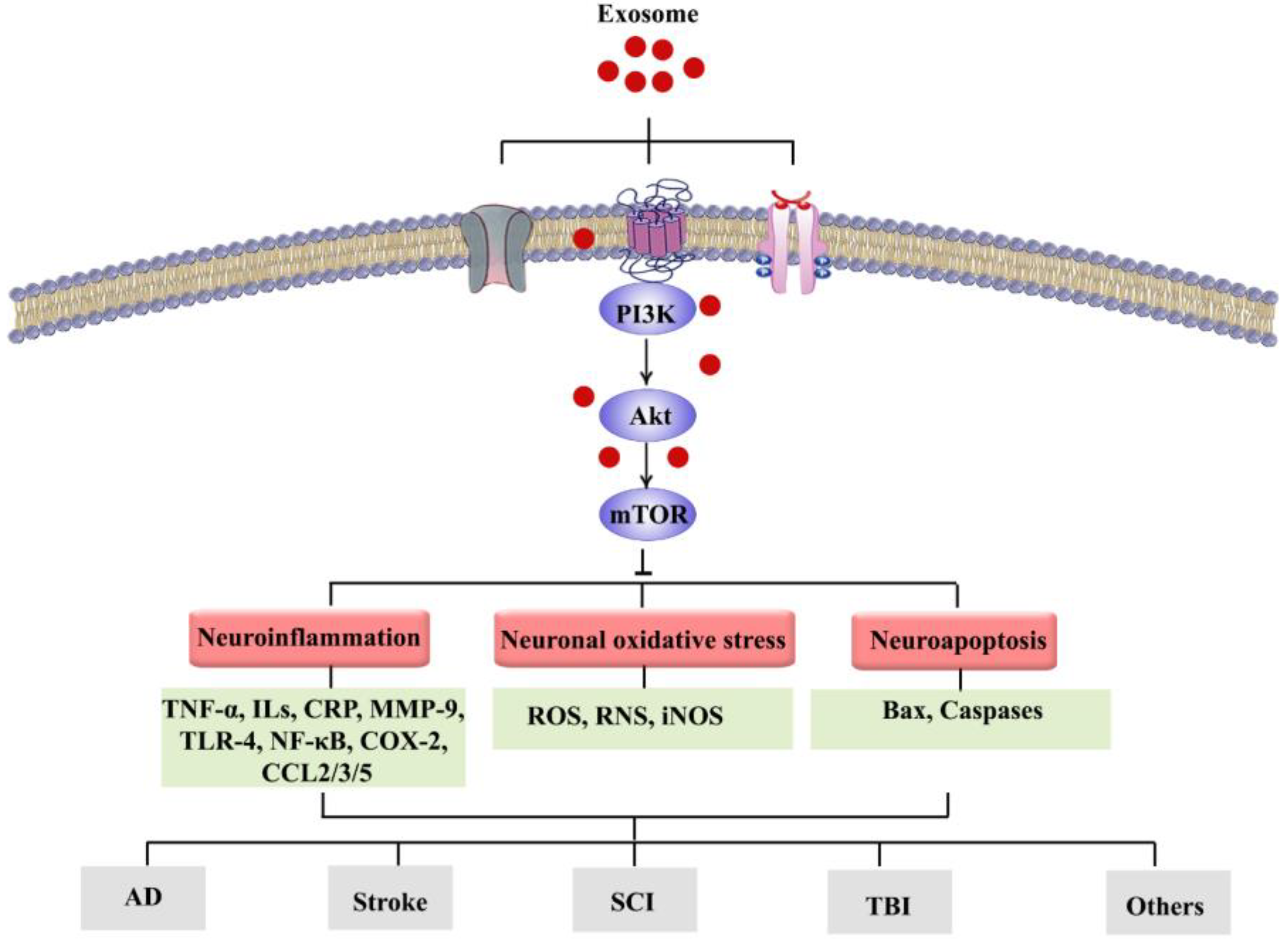
3. Neurodegenerative Diseases, Neurological Disorders, and Exosomes: Focusing on Pivotal Functions of PI3k/Akt/mTOR Signaling Pathway
3.1. Exosomes and Alzheimer’s Disease, Cognition, Learning and Memory
3.2. Exosomes and Stroke
3.3. Exosomes and Spinal Cord Injury
3.4. Exosomes and Traumatic Brain Injury
3.5. Exosomes and Other Neurological Disorders
4. Conclusions and Future Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Ou, G.-Y.; Lin, W.-W.; Zhao, W.-J. Neuregulins in neurodegenerative diseases. Front. Aging Neurosci. 2021, 13, 170. [Google Scholar] [CrossRef]
- Dugger, B.N.; Dickson, D.W. Pathology of neurodegenerative diseases. Cold Spring Harb. Perspect. Biol. 2017, 9, a028035. [Google Scholar] [CrossRef] [Green Version]
- Ayeni, E.A.; Aldossary, A.M.; Ayejoto, D.A.; Gbadegesin, L.A.; Alshehri, A.A.; Alfassam, H.A.; Afewerky, H.K.; Almughem, F.A.; Bello, S.M.; Tawfik, E.A. Neurodegenerative Diseases: Implications of Environmental and Climatic Influences on Neurotransmitters and Neuronal Hormones Activities. Int. J. Environ. Res. Public Health 2022, 19, 12495. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, G.; Sehgal, A.; Bhardwaj, S.; Singh, S.; Buhas, C.; Judea-Pusta, C.; Uivarosan, D.; Munteanu, M.A.; Bungau, S. Multifaceted role of matrix metalloproteinases in neurodegenerative diseases: Pathophysiological and therapeutic perspectives. Int. J. Mol. Sci. 2021, 22, 1413. [Google Scholar] [CrossRef]
- Zheng, J.C.; Chen, S. Translational Neurodegeneration in the era of fast growing international brain research. Transl. Neurodegener. 2022, 11, 1. [Google Scholar] [CrossRef]
- Fakhri, S.; Abdian, S.; Zarneshan, S.N.; Moradi, S.Z.; Farzaei, M.H.; Abdollahi, M. Nanoparticles in Combating Neuronal Dysregulated Signaling Pathways: Recent Approaches to the Nanoformulations of Phytochemicals and Synthetic Drugs Against Neurodegenerative Diseases. Int. J. Nanomed. 2022, 17, 299–331. [Google Scholar] [CrossRef]
- Manoharan, S.; Guillemin, G.J.; Abiramasundari, R.S.; Essa, M.M.; Akbar, M.; Akbar, M.D. The Role of Reactive Oxygen Species in the Pathogenesis of Alzheimer’s Disease, Parkinson’s Disease, and Huntington’s Disease: A Mini Review. Oxidative Med. Cell. Longev. 2016, 2016, 8590578. [Google Scholar] [CrossRef]
- Fakhri, S.; Pesce, M.; Patruno, A.; Moradi, S.Z.; Iranpanah, A.; Farzaei, M.H.; Sobarzo-Sánchez, E. Attenuation of Nrf2/Keap1/ARE in Alzheimer’s disease by plant secondary metabolites: A mechanistic review. Molecules 2020, 25, 4926. [Google Scholar] [CrossRef]
- Fakhri, S.; Iranpanah, A.; Gravandi, M.M.; Moradi, S.Z.; Ranjbari, M.; Majnooni, M.B.; Echeverría, J.; Qi, Y.; Wang, M.; Liao, P. Natural products attenuate PI3K/Akt/mTOR signaling pathway: A promising strategy in regulating neurodegeneration. Phytomedicine 2021, 91, 153664. [Google Scholar] [CrossRef]
- Milatovic, D.; Zaja-Milatovic, S.; Brockett, M.M.; Breyer, R.M.; Aschner, M.; Montine, T.J. Neuroinflammation and oxidative injury in developmental neurotoxicity. In Reproductive and Developmental Toxicology; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1129–1140. [Google Scholar]
- Disabato, D.J.; Quan, N.; Godbout, J.P. Neuroinflammation: The devil is in the details. J. Neurochem. 2016, 139, 136–153. [Google Scholar] [CrossRef] [Green Version]
- Long, H.-Z.; Cheng, Y.; Zhou, Z.-W.; Luo, H.-Y.; Wen, D.-D.; Gao, L.-C. PI3K/AKT signal pathway: A target of natural products in the prevention and treatment of Alzheimer’s disease and Parkinson’s disease. Front. Pharmacol. 2021, 12, 648636. [Google Scholar] [CrossRef]
- Moghaddam, R.H.; Samimi, Z.; Moradi, S.Z.; Little, P.J.; Xu, S.; Farzaei, M.H. Naringenin and naringin in cardiovascular disease prevention: A preclinical review. Eur. J. Pharmacol. 2020, 887, 173535. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khanbabapour Sasi, A.; Hussen, B.M.; Shoorei, H.; Siddiq, A.; Taheri, M.; Ayatollahi, S.A. Interplay between PI3K/AKT pathway and heart disorders. Mol. Biol. Rep. 2022, 49, 9767–9781. [Google Scholar] [CrossRef]
- Tokuhira, N.; Kitagishi, Y.; Suzuki, M.; Minami, A.; Nakanishi, A.; Ono, Y.; Kobayashi, K.; Matsuda, S.; Ogura, Y. PI3K/AKT/PTEN pathway as a target for Crohn’s disease therapy. Int. J. Mol. Med. 2015, 35, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Follo, M.Y.; Manzoli, L.; Poli, A.; McCubrey, J.A.; Cocco, L. PLC and PI3K/Akt/mTOR signalling in disease and cancer. Adv. Biol. Regul. 2015, 57, 10–16. [Google Scholar] [CrossRef]
- Chong, Z.Z.; Shang, Y.C.; Wang, S.; Maiese, K. A critical kinase cascade in neurological disorders: PI3K, Akt and mTOR. Future Neurol. 2012, 7, 733–748. [Google Scholar] [CrossRef] [Green Version]
- Kowshik, J.; Nivetha, R.; Ranjani, S.; Venkatesan, P.; Selvamuthukumar, S.; Veeravarmal, V.; Nagini, S. Astaxanthin inhibits hallmarks of cancer by targeting the PI3K/NF-κΒ/STAT3 signalling axis in oral squamous cell carcinoma models. IUBMB Life 2019, 71, 1595–1610. [Google Scholar] [CrossRef]
- Pena-Leon, V.; Perez-Lois, R.; Seoane, L.M. mTOR pathway is involved in energy homeostasis regulation as a part of the gut–brain axis. Int. J. Mol. Sci. 2020, 21, 5715. [Google Scholar] [CrossRef]
- Heras-Sandoval, D.; Pérez-Rojas, J.M.; Pedraza-Chaverri, J. Novel compounds for the modulation of mTOR and autophagy to treat neurodegenerative diseases. Cell. Signal. 2020, 65, 109442. [Google Scholar] [CrossRef]
- Danielsen, S.A.; Eide, P.W.; Nesbakken, A.; Guren, T.; Leithe, E.; Lothe, R.A. Portrait of the PI3K/AKT pathway in colorectal cancer. Biochim. Et Biophys. Acta (BBA) Rev. Cancer 2015, 1855, 104–121. [Google Scholar] [CrossRef]
- Wang, D.; Zhou, W.; Chen, J.; Wei, W. Upstream regulators of phosphoinositide 3-kinase and their role in diseases. J. Cell. Physiol. 2019, 234, 14460–14472. [Google Scholar] [CrossRef] [PubMed]
- Zarneshan, S.N.; Fakhri, S.; Farzaei, M.H.; Khan, H.; Saso, L. Astaxanthin targets PI3K/Akt signaling pathway toward potential therapeutic applications. Food Chem. Toxicol. 2020, 145, 111714. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Luo, J.; Guo, J.; Yao, X.; Jing, X.; Guo, F. The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A narrative review. Osteoarthr. Cartil. 2020, 28, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Weichhart, T.; Säemann, M. The PI3K/Akt/mTOR pathway in innate immune cells: Emerging therapeutic applications. Ann. Rheum. Dis. 2008, 67, iii70–iii74. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, J.; Sun, Y.; Tang, M.; Kong, L. Effect and mechanism of PI3K/AKT/mTOR signaling pathway in the apoptosis of GC-1 cells induced by nickel nanoparticles. Chemosphere 2020, 255, 126913. [Google Scholar] [CrossRef]
- Yang, C.; Hassan, Y.I.; Liu, R.; Zhang, H.; Chen, Y.; Zhang, L.; Tsao, R. Anti-inflammatory effects of different astaxanthin isomers and the roles of lipid transporters in the cellular transport of astaxanthin isomers in Caco-2 cell monolayers. J. Agric. Food Chem. 2019, 67, 6222–6231. [Google Scholar] [CrossRef]
- Moradi, S.Z.; Jalili, F.; Hoseinkhani, Z.; Mansouri, K. Regenerative Medicine and Angiogenesis; Focused on Cardiovascular Disease. Adv. Pharm. Bull. 2022, 12, 686–699. [Google Scholar] [CrossRef]
- O’Connor, D.M.; Boulis, N.M. Gene therapy for neurodegenerative diseases. Trends Mol. Med. 2015, 21, 504–512. [Google Scholar] [CrossRef]
- Durães, F.; Pinto, M.; Sousa, E. Old drugs as new treatments for neurodegenerative diseases. Pharmaceuticals 2018, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Lunn, J.S.; Sakowski, S.A.; Hur, J.; Feldman, E.L. Stem cell technology for neurodegenerative diseases. Ann. Neurol. 2011, 70, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Tăuţan, A.-M.; Ionescu, B.; Santarnecchi, E. Artificial intelligence in neurodegenerative diseases: A review of available tools with a focus on machine learning techniques. Artif. Intell. Med. 2021, 117, 102081. [Google Scholar] [CrossRef] [PubMed]
- Bellingham, S.; Guo, B.; Coleman, B.; Hill, A. Exosomes: Vehicles for the Transfer of Toxic Proteins Associated with Neurodegenerative Diseases? Front. Physiol. 2012, 3, 124. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Jordan, V.; Blenkiron, C.; Chamley, L.W. Biodistribution of extracellular vesicles following administration into animals: A systematic review. J. Extracell. Vesicles 2021, 10, e12085. [Google Scholar] [CrossRef] [PubMed]
- da Costa, V.R.; Araldi, R.P.; Vigerelli, H.; D’Ámelio, F.; Mendes, T.B.; Gonzaga, V.; Policíquio, B.; Colozza-Gama, G.A.; Valverde, C.W.; Kerkis, I. Exosomes in the tumor microenvironment: From biology to clinical applications. Cells 2021, 10, 2617. [Google Scholar] [CrossRef] [PubMed]
- Jarmalavičiūtė, A.; Pivoriūnas, A. Exosomes as a potential novel therapeutic tools against neurodegenerative diseases. Pharmacol. Res. 2016, 113, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Howitt, J.; Hill, A.F. Exosomes in the pathology of neurodegenerative diseases. J. Biol. Chem. 2016, 291, 26589–26597. [Google Scholar] [CrossRef] [Green Version]
- Pegtel, D.M.; Gould, S.J. Exosomes. Annu. Rev. Biochem. 2019, 88, 487–514. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Zetterberg, H.; Blennow, K.; de Leon, M. Alzheimer’s disease. Lancet 2006, 368, 387. [Google Scholar]
- Zhang, X.X.; Tian, Y.; Wang, Z.T.; Ma, Y.H.; Tan, L.; Yu, J.T. The Epidemiology of Alzheimer’s Disease Modifiable Risk Factors and Prevention. J. Prev. Alzheimer’s Dis. 2021, 8, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, Y.; Zhu, Y.; Chen, X.; Lin, T.; Zhou, D. Adipose Mesenchymal Stem Cell-Derived Exosomes Enhance PC12 Cell Function through the Activation of the PI3K/AKT Pathway. Stem Cells Int. 2021, 2021, 2229477. [Google Scholar] [CrossRef]
- Xiong, W.-P.; Yao, W.-Q.; Wang, B.; Liu, K. BMSCs-exosomes containing GDF-15 alleviated SH-SY5Y cell injury model of Alzheimer’s disease via AKT/GSK-3β/β-catenin. Brain Res. Bull. 2021, 177, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sui, H.; Zheng, Y.; Jiang, Y.; Shi, Y.; Liang, J.; Zhao, L. Curcumin-primed exosomes potently ameliorate cognitive function in AD mice by inhibiting hyperphosphorylation of the Tau protein through the AKT/GSK-3β pathway. Nanoscale 2019, 11, 7481–7496. [Google Scholar] [CrossRef]
- Wei, H.; Xu, Y.; Chen, Q.; Chen, H.; Zhu, X.; Li, Y. Mesenchymal stem cell-derived exosomal miR-223 regulates neuronal cell apoptosis. Cell Death Dis. 2020, 11, 290. [Google Scholar] [CrossRef]
- Spinelli, M.; Natale, F.; Rinaudo, M.; Leone, L.; Mezzogori, D.; Fusco, S.; Grassi, C. Neural Stem Cell-Derived Exosomes Revert HFD-Dependent Memory Impairment via CREB-BDNF Signalling. Int. J. Mol. Sci. 2020, 21, 8994. [Google Scholar] [CrossRef]
- Lucarini, M.; Durazzo, E.B.; Tzvetkov, N.T.; Jóźwik, M.S.; Strzałkowska, N.; Horbańczuk, A.G. Quercetin: Total-scale literature landscape analysis of a valuable nutraceutical with numerous potential applications in the promotion of human and animal health–a review. Anim. Sci. Pap. Rep. 2021, 39, 199–212. [Google Scholar]
- Soliman, H.M.; Ghonaim, G.A.; Gharib, S.M.; Chopra, H.; Farag, A.K.; Hassanin, M.H.; Nagah, A.; Emad-Eldin, M.; Hashem, N.E.; Yahya, G. Exosomes in Alzheimer’s disease: From being pathological players to potential diagnostics and therapeutics. Int. J. Mol. Sci. 2021, 22, 10794. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Mitsutake, S.; Igarashi, Y. Sphingolipid-modulated exosome secretion promotes clearance of amyloid-β by microglia. J. Biol. Chem. 2012, 287, 10977–10989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, H.; Liu, Z.; Buller, B.; Li, Y.; Golembieski, W.; Gan, X.; Wang, F.; Lu, M.; Ali, M.M.; Zhang, Z.G. MiR-17-92 enriched exosomes derived from multipotent mesenchymal stromal cells enhance axon-myelin remodeling and motor electrophysiological recovery after stroke. J. Cereb. Blood Flow Metab. 2021, 41, 1131–1144. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Katakowski, M.; Wang, F.; Qian, J.-Y.; Liu, X.S.; Ali, M.M.; Buller, B.; Zhang, Z.G.; Chopp, M. MicroRNA-17–92 cluster in exosomes enhance neuroplasticity and functional recovery after stroke in rats. Stroke 2017, 48, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.-Y.; Song, J.-X.; Cai, H.; Wang, P.-P.; Yin, Q.-L.; Zhang, Y.-D.; Chen, J.; Li, M.; Song, J.-J.; Wang, Y.-L. Healthy Serum-Derived Exosomes Improve Neurological Outcomes and Protect Blood–Brain Barrier by Inhibiting Endothelial Cell Apoptosis and Reversing Autophagy-Mediated Tight Junction Protein Reduction in Rat Stroke Model. Front. Cell. Neurosci. 2022, 16, 841544. [Google Scholar] [CrossRef]
- Pan, Q.; Kuang, X.; Cai, S.; Wang, X.; Du, D.; Wang, J.; Wang, Y.; Chen, Y.; Bihl, J.; Chen, Y. miR-132-3p priming enhances the effects of mesenchymal stromal cell-derived exosomes on ameliorating brain ischemic injury. Stem Cell Res. Ther. 2020, 11, 260. [Google Scholar] [CrossRef]
- Duan, S.; Wang, F.; Cao, J.; Wang, C. Exosomes derived from MicroRNA-146a-5p-enriched bone marrow mesenchymal stem cells alleviate intracerebral hemorrhage by inhibiting neuronal apoptosis and microglial M1 polarization. Drug Des. Dev. Ther. 2020, 14, 3143. [Google Scholar] [CrossRef]
- Shen, H.; Yao, X.; Li, H.; Li, X.; Zhang, T.; Sun, Q.; Ji, C.; Chen, G. Role of exosomes derived from miR-133b modified MSCs in an experimental rat model of intracerebral hemorrhage. J. Mol. Neurosci. 2018, 64, 421–430. [Google Scholar] [CrossRef]
- Wang, P.; Xue, Y.; Zuo, Y.; Xue, Y.; Zhang, J.H.; Duan, J.; Liu, F.; Liu, A. Exosome-Encapsulated microRNA-140-5p Alleviates Neuronal Injury Following Subarachnoid Hemorrhage by Regulating IGFBP5-Mediated PI3K/AKT Signaling Pathway. Mol. Neurobiol. 2022, 59, 7212–7228. [Google Scholar] [CrossRef]
- Zhao, H.; Li, Y.; Chen, L.; Shen, C.; Xiao, Z.; Xu, R.; Wang, J.; Luo, Y. HucMSCs-derived miR-206-knockdown exosomes contribute to neuroprotection in subarachnoid hemorrhage induced early brain injury by targeting BDNF. Neuroscience 2019, 417, 11–23. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, J.; Liu, J.; Liu, H.; Li, J. Effects of stem cell-derived exosomes on neuronal apoptosis and inflammatory cytokines in rats with cerebral ischemia-reperfusion injury via PI3K/AKT pathway-mediated mitochondrial apoptosis. Immunopharmacol. Immunotoxicol. 2021, 43, 731–740. [Google Scholar] [CrossRef]
- Wu, W.; Liu, J.; Yang, C.; Xu, Z.; Huang, J.; Lin, J. Astrocyte-derived exosome-transported microRNA-34c is neuroprotective against cerebral ischemia/reperfusion injury via TLR7 and the NF-κB/MAPK pathways. Brain Res. Bull. 2020, 163, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Zhou, Y.; Liang, D.; He, H.; Liu, X.; Zhu, R.; Zhang, M.; Luo, X.; Wang, Y.; Huang, G. Exosomes secreted from bone marrow mesenchymal stem cells attenuate oxygen-glucose deprivation/reoxygenation-induced pyroptosis in PC12 cells by promoting AMPK-dependent autophagic flux. Front. Cell. Neurosci. 2020, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Bu, X.; Li, D.; Wang, F.; Sun, Q.; Zhang, Z. Protective role of astrocyte-derived exosomal microRNA-361 in cerebral ischemic-reperfusion injury by regulating the AMPK/mTOR signaling pathway and targeting CTSB. Neuropsychiatr. Dis. Treat. 2020, 16, 1863. [Google Scholar] [CrossRef]
- Hou, K.; Li, G.; Zhao, J.; Xu, B.; Zhang, Y.; Yu, J.; Xu, K. Bone mesenchymal stem cell-derived exosomal microRNA-29b-3p prevents hypoxic-ischemic injury in rat brain by activating the PTEN-mediated Akt signaling pathway. J. Neuroinflamm. 2020, 17, 46. [Google Scholar] [CrossRef] [Green Version]
- Deng, M.; Xiao, H.; Peng, H.; Yuan, H.; Xu, Y.; Zhang, G.; Tang, J.; Hu, Z. Preservation of neuronal functions by exosomes derived from different human neural cell types under ischemic conditions. Eur. J. Neurosci. 2018, 47, 150–157. [Google Scholar] [CrossRef]
- Halurkar, M.S.; Wang, J.; Chen, S.; Bihl, J.C. EPC-EXs improve astrocyte survival and oxidative stress through different uptaking pathways in diabetic hypoxia condition. Stem Cell Res. Ther. 2022, 13, 91. [Google Scholar] [CrossRef] [PubMed]
- Hankey, G.J. Stroke. Lancet 2017, 389, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Blanco, S.; Martínez-Lara, E.; Siles, E.; Peinado, M.Á. New Strategies for Stroke Therapy: Nanoencapsulated Neuroglobin. Pharmaceutics 2022, 14, 1737. [Google Scholar] [CrossRef]
- Liu, Y.; Mu, Y.; Li, Z.; Yong, V.W.; Xue, M. Extracellular matrix metalloproteinase inducer in brain ischemia and intracerebral hemorrhage. Front. Immunol. 2022, 13, 986469. [Google Scholar] [CrossRef]
- Mansfield, A.; Inness, E.L.; McIlroy, W.E. Stroke. Handb. Clin. Neurol. 2018, 159, 205–228. [Google Scholar] [CrossRef]
- Correa-Paz, C.; da Silva-Candal, A.; Polo, E.; Parcq, J.; Vivien, D.; Maysinger, D.; Pelaz, B.; Campos, F. New Approaches in Nanomedicine for Ischemic Stroke. Pharmaceutics 2021, 13, 757. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.D.; Eskey, C.J. Hemorrhagic stroke. Radiol. Clin. North Am. 2011, 49, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Huang, L.-Y.; Hong, R.; Song, J.-X.; Guo, X.-J.; Zhou, W.; Hu, Z.-L.; Wang, W.; Wang, Y.-L.; Shen, J.-G. Momordica charantia Exosome-Like Nanoparticles Exert Neuroprotective Effects Against Ischemic Brain Injury via Inhibiting Matrix Metalloproteinase 9 and Activating the AKT/GSK3β Signaling Pathway. Front. Pharmacol. 2022, 13, 908830. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Li, Y.; Deng, B.; Lin, A.; Zhang, G.; Ma, M.; Wang, Y.; Yang, Y.; Kang, X. The PI3K/AKT signalling pathway in inflammation, cell death and glial scar formation after traumatic spinal cord injury: Mechanisms and therapeutic opportunities. Cell Prolif. 2022, 55, e13275. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wang, C.Z.; Liu, J.; Anastasio, N.C.; Johnson, K.M. Brain-derived neurotrophic factor prevents phencyclidine-induced apoptosis in developing brain by parallel activation of both the ERK and PI-3K/Akt pathways. Neuropharmacology 2010, 58, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, Q.G.; Li, C.; Zhang, G.Y. Subtoxic N-methyl-d-aspartate delayed neuronal death in ischemic brain injury through TrkB receptor-and calmodulin-mediated PI-3K/Akt pathway activation. Hippocampus 2007, 17, 525–537. [Google Scholar] [CrossRef]
- Li, Y.; Tang, Y.; Yang, G.-Y. Therapeutic application of exosomes in ischaemic stroke. Stroke Vasc. Neurol. 2021, 6, 483–495. [Google Scholar] [CrossRef]
- Seyedaghamiri, F.; Salimi, L.; Ghaznavi, D.; Sokullu, E.; Rahbarghazi, R. Exosomes-based therapy of stroke, an emerging approach toward recovery. Cell Commun. Signal. 2022, 20, 110. [Google Scholar] [CrossRef]
- Liau, L.L.; Looi, Q.H.; Chia, W.C.; Subramaniam, T.; Ng, M.H.; Law, J.X. Treatment of spinal cord injury with mesenchymal stem cells. Cell Biosci. 2020, 10, 112. [Google Scholar] [CrossRef]
- Flack, J.A.; Sharma, K.D.; Xie, J.Y. Delving into the recent advancements of spinal cord injury treatment: A review of recent progress. Neural Regen. Res. 2022, 17, 283. [Google Scholar]
- Alcántar-Garibay, O.V.; Incontri-Abraham, D.; Ibarra, A. Spinal cord injury-induced cognitive impairment: A narrative review. Neural Regen. Res. 2022, 17, 2649. [Google Scholar] [PubMed]
- Ding, Y.; Chen, Q. mTOR pathway: A potential therapeutic target for spinal cord injury. Biomed. Pharmacother. 2022, 145, 112430. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tian, Z.; He, L.; Liu, C.; Wang, N.; Rong, L.; Liu, B. Exosomes derived from miR-26a-modified MSCs promote axonal regeneration via the PTEN/AKT/mTOR pathway following spinal cord injury. Stem Cell Res. Ther. 2021, 12, 224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Bai, Z.; Yi, W.; Hu, Z.; Hao, J. Overexpression of miR-338-5p in exosomes derived from mesenchymal stromal cells provides neuroprotective effects by the Cnr1/Rap1/Akt pathway after spinal cord injury in rats. Neurosci. Lett. 2021, 761, 136124. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Rong, Y.; Wang, J.; Zhou, Z.; Ge, X.; Ji, C.; Jiang, D.; Gong, F.; Li, L.; Chen, J. Exosome-shuttled miR-216a-5p from hypoxic preconditioned mesenchymal stem cells repair traumatic spinal cord injury by shifting microglial M1/M2 polarization. J. Neuroinflamm. 2020, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Xu, T.; Liu, W.; Rong, Y.; Wang, J.; Fan, J.; Yin, G.; Cai, W. Exosomes derived from GIT1-overexpressing bone marrow mesenchymal stem cells promote traumatic spinal cord injury recovery in a rat model. Int. J. Neurosci. 2021, 131, 170–182. [Google Scholar] [CrossRef]
- Wang, L.; Pei, S.; Han, L.; Guo, B.; Li, Y.; Duan, R.; Yao, Y.; Xue, B.; Chen, X.; Jia, Y. Mesenchymal stem cell-derived exosomes reduce A1 astrocytes via downregulation of phosphorylated NFκB P65 subunit in spinal cord injury. Cell. Physiol. Biochem. 2018, 50, 1535–1559. [Google Scholar] [CrossRef]
- Huang, J.-H.; Xu, Y.; Yin, X.-M.; Lin, F.-Y. Exosomes derived from miR-126-modified MSCs promote angiogenesis and neurogenesis and attenuate apoptosis after spinal cord injury in rats. Neuroscience 2020, 424, 133–145. [Google Scholar] [CrossRef]
- Jiang, D.; Gong, F.; Ge, X.; Lv, C.; Huang, C.; Feng, S.; Zhou, Z.; Rong, Y.; Wang, J.; Ji, C. Neuron-derived exosomes-transmitted miR-124-3p protect traumatically injured spinal cord by suppressing the activation of neurotoxic microglia and astrocytes. J. Nanobiotechnol. 2020, 18, 105. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, C.; Li, S.; Li, Z.; Lai, X.; Xia, Q. Exosomes derived from nerve stem cells loaded with FTY720 promote the recovery after spinal cord injury in rats by PTEN/AKT signal pathway. J. Immunol. Res. 2021, 2021, 8100298. [Google Scholar] [CrossRef]
- Pan, D.; Li, Y.; Yang, F.; Lv, Z.; Zhu, S.; Shao, Y.; Huang, Y.; Ning, G.; Feng, S. Increasing toll-like receptor 2 on astrocytes induced by Schwann cell-derived exosomes promotes recovery by inhibiting CSPGs deposition after spinal cord injury. J. Neuroinflammation 2021, 18, 172. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Zhu, S.; Zhang, W.; Wei, Z.; Yang, F.; Guo, Z.; Ning, G.; Feng, S. Autophagy induced by Schwann cell-derived exosomes promotes recovery after spinal cord injury in rats. Biotechnol. Lett. 2022, 44, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Xu, Y.; Chen, C.; Xie, H.; Lu, H.; Hu, J. Local delivery of USC-derived exosomes harboring ANGPTL3 enhances spinal cord functional recovery after injury by promoting angiogenesis. Stem Cell Res. Ther. 2021, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lai, X.; Wu, D.; Liu, B.; Wang, N.; Rong, L. Umbilical mesenchymal stem cell-derived exosomes facilitate spinal cord functional recovery through the miR-199a-3p/145-5p-mediated NGF/TrkA signaling pathway in rats. Stem Cell Res. Ther. 2021, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Lin, F.; Dong, J.; Liu, J.; Ding, Z.; Xu, J. Peripheral macrophage-derived exosomes promote repair after spinal cord injury by inducing local anti-inflammatory type microglial polarization via increasing autophagy. Int. J. Biol. Sci. 2021, 17, 1339. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Li, Y.; Huang, S.; Xu, H.; Li, H.; Liu, B. Resveratrol-primed exosomes strongly promote the recovery of motor function in SCI rats by activating autophagy and inhibiting apoptosis via the PI3K signaling pathway. Neurosci. Lett. 2020, 736, 135262. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wu, Q.; Wang, P.; Jing, Y.; Yao, H.; Tang, Y.; Li, Z.; Zhang, H.; Xiu, R. Exosomes derived from pericytes improve microcirculation and protect blood–spinal cord barrier after spinal cord injury in mice. Front. Neurosci. 2019, 13, 319. [Google Scholar] [CrossRef]
- Quillinan, N.; Herson, P.S.; Traystman, R.J. Neuropathophysiology of Brain Injury. Anesthesiol. Clin. 2016, 34, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Goetzl, E.J.; Elahi, F.M.; Mustapic, M.; Kapogiannis, D.; Pryhoda, M.; Gilmore, A.; Gorgens, K.A.; Davidson, B.; Granholm, A.-C.; Ledreux, A. Altered levels of plasma neuron-derived exosomes and their cargo proteins characterize acute and chronic mild traumatic brain injury. FASEB J. 2019, 33, 5082. [Google Scholar] [CrossRef]
- Gao, W.; Li, F.; Liu, L.; Xu, X.; Zhang, B.; Wu, Y.; Yin, D.; Zhou, S.; Sun, D.; Huang, Y. Endothelial colony-forming cell-derived exosomes restore blood-brain barrier continuity in mice subjected to traumatic brain injury. Exp. Neurol. 2018, 307, 99–108. [Google Scholar] [CrossRef]
- Huang, S.; Ge, X.; Yu, J.; Han, Z.; Yin, Z.; Li, Y.; Chen, F.; Wang, H.; Zhang, J.; Lei, P. Increased miR-124-3p in microglial exosomes following traumatic brain injury inhibits neuronal inflammation and contributes to neurite outgrowth via their transfer into neurons. FASEB J. 2018, 32, 512–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, J.; Ma, B.; Li, N.; Wang, S.; Sun, Z.; Xue, C.; Han, Q.; Wei, J.; Zhao, R.C. MSC-derived exosomes promote recovery from traumatic brain injury via microglia/macrophages in rat. Aging 2020, 12, 18274. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, H.; Yang, Y.; Ma, Y. Exosomal microRNAs have great potential in the neurorestorative therapy for traumatic brain injury. Exp. Neurol. 2022, 352, 114026. [Google Scholar] [CrossRef] [PubMed]
- Guedes, V.A.; Devoto, C.; Leete, J.; Sass, D.; Acott, J.D.; Mithani, S.; Gill, J.M. Extracellular vesicle proteins and microRNAs as biomarkers for traumatic brain injury. Front. Neurol. 2020, 11, 663. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, Y.; Zhu, Z.; Gu, C.; Waqas, A.; Chen, L. Emerging exosomes and exosomal MiRNAs in spinal cord injury. Front. Cell Dev. Biol. 2021, 9, 703989. [Google Scholar] [CrossRef]
- Harman, D. Aging: Overview. Ann. New York Acad. Sci. 2001, 928, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Bonafede, R.; Brandi, J.; Manfredi, M.; Scambi, I.; Schiaffino, L.; Merigo, F.; Turano, E.; Bonetti, B.; Marengo, E.; Cecconi, D. The anti-apoptotic effect of ASC-exosomes in an in vitro ALS model and their proteomic analysis. Cells 2019, 8, 1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassew, N.G.; Charish, J.; Shabanzadeh, A.P.; Luga, V.; Harada, H.; Farhani, N.; D’Onofrio, P.; Choi, B.; Ellabban, A.; Nickerson, P.E. Exosomes mediate mobilization of autocrine Wnt10b to promote axonal regeneration in the injured CNS. Cell Rep. 2017, 20, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liang, Y.; Xu, Z.; Liu, J.; Liu, X.; Ma, J.; Sun, C.; Yang, Y. Exosomal miR-673-5p from fibroblasts promotes Schwann cell-mediated peripheral neuron myelination by targeting the TSC2/mTORC1/SREBP2 axis. J. Biol. Chem. 2022, 298, 101718. [Google Scholar] [CrossRef]
- Zhang, Y.; Chopp, M.; Liu, X.S.; Katakowski, M.; Wang, X.; Tian, X.; Wu, D.; Zhang, Z.G. Exosomes derived from mesenchymal stromal cells promote axonal growth of cortical neurons. Mol. Neurobiol. 2017, 54, 2659–2673. [Google Scholar] [CrossRef] [Green Version]
- Feng, N.; Jia, Y.; Huang, X. Exosomes from adipose-derived stem cells alleviate neural injury caused by microglia activation via suppressing NF-kB and MAPK pathway. J. Neuroimmunol. 2019, 334, 576996. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Chen, Y.; Xue, C.; Ma, B.; Shen, Y.; Guan, J.; Bao, X.; Wu, H.; Han, Q.; Wang, R. Protection of nerve injury with exosome extracted from mesenchymal stem cell. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. Acta Acad. Med. Sin. 2016, 38, 33–36. [Google Scholar]
- Cui, Y.; Liu, C.; Huang, L.; Chen, J.; Xu, N. Protective effects of intravitreal administration of mesenchymal stem cell-derived exosomes in an experimental model of optic nerve injury. Exp. Cell Res. 2021, 407, 112792. [Google Scholar] [CrossRef]
- Sarko, D.K.; McKinney, C.E. Exosomes: Origins and therapeutic potential for neurodegenerative disease. Front. Neurosci. 2017, 11, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Chen, W.; Ye, J.; Wang, Y. Potential role of exosomes in ischemic stroke treatment. Biomolecules 2022, 12, 115. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Corbett, A.L.; Taatizadeh, E.; Tasnim, N.; Little, J.P.; Garnis, C.; Daugaard, M.; Guns, E.; Hoorfar, M.; Li, I.T. Challenges and opportunities in exosome research—Perspectives from biology, engineering, and cancer therapy. APL Bioeng. 2019, 3, 011503. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Source of Exosomes | Cargo or Intermediate Molecule | Disease | Method (In Vitro/In Vivo) | Dose/Route of Administration | Mechanism of Actions and Outcomes | References |
|---|---|---|---|---|---|---|
| ADSC | _____ | AD | in vitro: PC12 cells | 10, 50 and 100 μg/mL | ↑p-PI3K, p-Akt, CD29, CD44, CD73, CD105, cell proliferation, and migration; ↓CD45, HLA-DR, and cell apoptosis | [44] |
| BMSCs | GDF-15 | AD | in vitro: SH-SY5Y cells | _____ | ↑Cell viability, Akt/GSK-3β/β-catenin pathway; ↓Apoptosis, TNF-α, IL-6, IL-1β, and IL-8 | [45] |
| Macrophage cell line (RAW 264.7) | Curcumin | AD | in vivo: Okadaic acid-induced AD in C57BL/6 mice; in vitro: hCMEC/D3 cells | 0.4 mg/kg, single i.v. dose, and 100 μg/mL, i.p. for 7 days | ↑Curcumin solubility, stability, bioavailability, cellular uptake and BBB-crossing, p-Akt, p-Ser9 GSK-3β; ↓Learning deficiencies, cognitive decline, escape latency, cell apoptosis, tau hyperphosphorylation, p-Ser396 tau, neuronal injury, Bax and c-caspase 3; | [46] |
| HucMSCs | miR-223 | AD | in vitro: Aβ1–40-induced injury in SH-SY5Y cells under hypoxic conditions | 2 μg per 1 × 105 recipient cells | ↑p-Akt; ↓Apoptosis, scratch area, IL-6, IL-1β, TNF-α, CRP, and PTEN | [47] |
| NSC | _____ | HFD-related cognitive decline | in vivo: male C57BL/6 mice | 1.5 μg per nostril, 3 times per week | ↑BDNF, nNOS, Sirt1, Egr3, RelA, and pTrkB; ↓Memory impairment | [48] |
| MSCs | miR-132-3p | VD | in vivo: VD-induced in male C57BL/6 mice; in vitro: OGD-injured neurons | 1 × 1010 particles/100 μL, via the tail vein once every 7 days for 21 days | ↑Cognitive function, neuron number, dendritic spine density, synaptic plasticity, Ras, p-Akt, p-GSK-3β, and neurite elongation; ↓Aβ, p-tau, RASA1, and apoptosis | [49] |
| MSCs | miR-17-92 | Stroke | in vivo: 2 h intraluminal filament-induced MCAO in Wistar rats; Ex-vivo rat organotypic brain slice culture model | 3 × 1011 particles/rat, i.v. | ↑GAP-43 immunoreactivity, cortical and intracortical axonal density, myelin density, neuronal plasticity, contralesional axon number and total length, CST axonal remodeling, and functional recovery ↓Time to remove the adhesive tabs, mNSS score, and lowest threshold value of ICMS; ↑MBP+ myelin and NFH+, PTEN/PI3K/Akt/mTOR pathway | [53] |
| MSCs | miR-17–92 | Stroke | Wistar rats were subjected to 2 h intraluminal filament-induced MCAO | 100 μg total exosome protein or 3 × 1011 particles per rat, i.v. | ↑Functional recovery, axonal density, p-NF-H immunoreactive area, synaptophysin immunoreactivity, primary and secondary neurite branching, spine density, dendritic plasticity, neurogenesis, oligodendrogenesis, p-Akt, p-mTOR, and p-GSK-3β; ↓mNSS score, number of Foot-fault and PTEN level | [54] |
| Healthy rat serum-derived exosomes | _____ | Stroke | in vivo: Focal cerebral ischemia induced by the intraluminal suture MCAO method in SD rats in vitro: OGD/R injury model in bEnd.3 immortalized mouse brain endothelial cells | 800 μg/kg, i.v. 50 μg/mL | ↑Neurobehavioral scores, total moving distance, neuronal spine density, claudin-5, ZO-1, Bcl-2/Bax ratio, p-Akt/Akt and SQSTM1/p62 expression, ↓Infarct volumes, %distance and %time in center, BBB leakage, Evans blue dye extravasation, MMP-9, cleaved caspase-3 and LC3B-II/LC3B-I ratio; ↑SQSTM1/p62 expression, ↓Apoptotic cells, TUNEL+/CD31+ cells, cleaved caspase-3 and LC3B-II/LC3B-I ratio | [55] |
| MSCs | miR-132-3p | Ischemic stroke | in vivo: Focal ischemic stroke induced by transient MCAO in C57BL/6 mice in vitro: H/R injury model in mouse brain microvascular endothelial cells | 1 × 1010 particles/100 μL in PBS via the tail vein, i.v. 50 μg/mL | ↑cMVD and CBF;↓ROS, apoptosis, Evans blue dye extravasation, brain water content, infarct volume, NDS, and BBB disruption; ↑Ras, p-PI3K/PI3K, p-Akt/Akt and p-eNOS/eNOS, ZO-1, and Claudin-5; ↓RASA1, ROS, paracelluar permeability and apoptosis | [56] |
| BMSCs | miR-146a-5p | ICH | in vivo: Collagenase type IV-induced ICH in male SD rats, by an intrastriatal injection | 100 μg/mL, 100 μg via the tail vein, i.v. | ↑SOD and neurological function; ↓Microglia M1 polarization, iNOS, COX-2, MCP-1, IRAK1, NFAT5, TNF-α, IL-1β, IL-6, MPO-positive cells, MDA, OX42-positive cells, Iba-1+/MHC-II+, apoptotic, and degenerative neurons | [57] |
| MSCs | miR-133b | ICH | in vivo: An autologous arterial blood ICH model in adult male SD rats | 100 μg via the tail vein, i.v., 72 h after ICH | ↑p-ERK1/2/ERK1/2 and p-CREB/CREB; ↓RhoA expression, neuronal apoptosis and neurodegenerative neurons | [58] |
| ADSCs | miR-140-5p | SAH | in vivo: SAH-induced neurological dysfunction in rat in vitro: TDP-43-induced neuronal injury | _____ | ↑Cell viability and PI3K/Akt activation; ↓IGFBP5 expression and apoptosis | [59] |
| HucMSCs | miR-206-knockdown | SAH | in vivo: SAH-induced EBI in double blood injection model in SD rats | 200 μL PBS containing 400 μg exosomes, i.v. injected into the femoral vein 1 h after SAH | ↑Bcl-2, BDNF, TrkB, and p-CREB; ↓Bax, caspase-8, neurological deficit, brain edema, and neuronal apoptosis | [60] |
| SC | _____ | Cerebral I/R injury | in vivo: Focal cerebral I/R induced by the improved Longa method in rats | _____ | ↑Latency for the novel arm, IFN-γ, and Bcl-2; ↓Novel entries, IL-1α, IL-2, TNF-α, Bax, cleaved caspase-3, cleaved caspase-9, CytC, PI3K, Akt, and neural cell apoptosis | [61] |
| ATCs | miR-34c | Cerebral I/R injury | in vivo: Wistar rats MCAO model in vitro: OGD/R model in N2a mouse neuroblastoma cells | 20 and 30 μg/mL via the tail vein after ischemia, 20 μM | ↑Nissl bodies and c-fos positive cell numbers; ↓Neuronal injury, NDS, infarct volume, brain water content, IL-6, IL-8, and TNF-α; ↑Cell proliferation, EdU positive cell index, and TLR7; ↓Bax, cleaved caspase-3, cleaved PARP, apoptosis, and NF-κB/MAPK axis | [62] |
| BMSCs | _____ | Cerebral I/R injury | in vitro: OGD/R model in PC12 cells | 10 μg/mL | ↑Cell viability, autophagic flux, p-AMPK/AMPK; ↓LDH, morphological changes, pyroptosis, NLRP3, ROS, cleaved caspase-1, IL-1β, GSDMD-N, p-mTOR/mTOR, and P62 | [63] |
| ATCs | miR-361 | Cerebral I/R injury | in vivo: Wistar rats reversible MCAO model in vitro: OGD/R model in PC12 mouse neuroblastoma cells | 2 mL exosomes (30 μg/mL) via the caudal vein, twice a week for 2 weeks 30 μg/mL | ↑Nissl bodies, C-fos and neuronal viability; ↓Nerve damage, NDS, brain water content, infarct volume, cerebral edema, apoptosis, AMPK and mTOR mRNA and protein levels; ↑Cell activity, cell proliferation, and EdU positive cell index; ↓Apoptosis, Bax, cleaved caspase-3, cleaved PARP, CTSB, AMPK and mTOR mRNA and protein levels | [64] |
| BMSCs | miR-29b-3p | Hypoxic-ischemic brain injury | in vivo: Cerebral ischemia induced by MCAO method in SD rats in vitro: OGD/R injury model in rat primary cortical neurons and BMECs | 100 μg/kg/day intracerebroventricular stereotactic injection 2 h after the MCAO model, every day for 3 days | ↑Bcl-2, VEGFA, VEGFR2, angiogenesis, and p-Akt/Akt; ↓Apoptotic cells, Bax, cleaved caspase-3, infarct volume, MVD, and PTEN; | [65] |
| Neuron, EC, NPC and ATC differentiated from H9 hES | _____ | Ischemia | OGD-induced injury in H9 hES derived neurons | 100 μg/mL | ↑Neuronal survival rate, p-PI3K p85, p-Akt, p-mTOR, Bcl-2, and basal neuronal synaptic transmission; ↓Neuronal damage, p-AMPK, COX-2, iNOS, TNF-α, Bax, and cleaved caspase-3 | [66] |
| EPCs | miR-126 | Diabetic ischemic stroke | H/R and HG-induced injury in human astrocytes | 3 × 109 particles/mL | ↓Cytotoxicity, ROS and lipid peroxidation | [67] |
| Source of Exosomes | Cargo or Intermediate Molecule | Disease | Method (In Vitro/In Vivo) | Dose/Route of Administration | Mechanism of Actions and Outcomes | References |
|---|---|---|---|---|---|---|
| BMSCs | miR-26a | SCI | in vivo: SCI induced in SD rats in vitro: PC12 cells | 200 μg in 200 μL PBS via tail vein injection 20 μg/mL for 48 h | ↑Axonal regeneration, neurogenesis, functional recovery, BBB scores, MEP amplitudes, neurofilament density, Tuj-1, p-Akt, p-PI3K, p-mTOR, p-S6K, and p62; ↓Glial scarring, GFAP, astrocyte inflammation, autophagy, p-AMPK, p-ULK1, p-IKB, and p-p65; ↑Neurofilament generation, nerve regeneration, p-Akt, p-PI3K, and p-mTOR, ↓PTEN, autophagy, p-AMPK, p-ULK1, p-IKB and p-p65 | [84] |
| BMSCs | miR-338-5p | SCI | in vivo: SCI induced in SD rats in vitro: H2O2-induced oxidative stress injury in PC12 cells | 100 µg (50 µg microinjected to injured site +50 µg via the tail vein) in PBS at 5 min and 1 h after SCI, 100 μg of total protein | ↑NF-M and GAP43; ↓MAG and GFAP; ↑Cell viability, SOD, NF-M, GAP43, Bcl-2, cAMP, Rap1, p-Akt, and p-PI3K; ↓Apoptosis, ROS, MAG, GFAP, Bax, cleaved caspase-3, and Cnr1 | [85] |
| BMSCs under hypoxia | miR-216a-5p | SCI | in vivo: SCI induced in C57BL/6 mice in vitro: LPS-stimulated BV2 microglial cells and primary microglia | 200 μg of total protein via tail vein injection 200 μg/mL | ↑Functional recovery, BMS score, gait recovery, motor coordination, NeuN-positive neurons, MEP amplitudes, IL-4, IL-10, TGF-β, Arg1, CD206, YM1/2, and M1 to M2 polarization; ↓Neurofilament 200, lesion volume, iNOS, TNF-α, IL-1β, and IL-6; ↑IL-4, IL-10, Arg1, CD206, YM1/2, M1 to M2 polarization, TGF-β, p-Akt, and p-PI3K; ↓TNF-α, IL-1β, IL-6, iNOS, TLR4, p-P65, and MyD88; | [86] |
| GIT1-BMSCs | _____ | SCI | in vivo: SCI induced in SD rats in vitro: Glutamate-induced injury model in neuronal cells | 200 μg of total protein via tail vein injection 100 µg/mL | ↑Nissl bodies, neural regeneration, BBB score, motor function, Bcl-2, and P-Akt; ↓Apoptosis, glial scar formation, TNF-α, IL-1β, IL-6, Bax, and cleaved caspase-3; ↑Bcl-2; ↓TUNEL-positive cells, neural apoptosis, Bax, cleaved caspase-3, and caspase-9 | [87] |
| MSCs | _____ | SCI | in vivo: SCI induced in SD rats in vitro: Astrocytes isolated from SCI rats | 1 × 106 in 200 μL PBS via tail vein injection, 5 × 104 | ↑Functional recovery, MBP, BBB scores, Syn, and NeuN; ↓Lesion size, morphological phenomena, p65+ nuclei, A1 astrocytes, TNF-α, IL-1α, IL-1β, C3, GFAP, TUNEL-positive cells, p-IKBα, and p-p65 | [88] |
| MSCs | miR-126 | SCI | in vivo: SCI induced in SD rats in vitro: OGD injury model in HUVECs | 100 µg of total protein in 0.5 mL PBS via tail vein injection 10 µg | ↑Functional recovery, VEGF, angiogenesis, neurogenesis, blood vessels number and connectivity value, Bcl-2, NeuN, Sox2, and Nestin positive cells; ↓Lesion volume, incorrect steps, apoptosis, SPRED1, PIK3R2, Bax and cleaved caspase-3; ↑Angiogenesis and HUVECs migration; ↓SPRED1 and PIK3R2 | [89] |
| Neuron-derived exosomes | miR-124-3p | SCI | in vivo: SCI induced in C57BL/6 mice in vitro: Primary microglial cultures and MCM, primary astrocyte cultures and primary neuronal cultures | 200 μg of total protein in 200 μL of PBS via tail vein injection 200 μg/mL | ↑Functional recovery, BMS score, gait recovery, motor coordination, MEP amplitudes, and hind limb alternation; ↓Forelimb dependence, axonal damage, TNF-α, IL-1α, IL-6, IL-1β, C1q, M1 microglia, iNOS, C3, and A1 astrocytes; ↑p-PI3K and p-Akt; ↓M1 microglia, iNOS, and p-P65 | [90] |
| NSCs | FTY720 | SCI | in vivo: SCI induced in SD rats in vitro: SCMECs hypoxic model | 20 μg in 0.3 mL PBS via tail vein injection 20 μg/mL | ↑Locomotor function, complete tissue structure, claudin-5, and Bcl-2; ↓Edema formation, inflammatory cell infiltration, SCI lesion, neuronal cell apoptosis, AQP4, and Bax; ↑ZO-1 and p-Akt ↓PTEN and SCMEC permeability | [91] |
| Primary SCDEs | _____ | SCI | in vivo: SCI induced in mice in vitro: H2O2-induced injury in spinal cord astrocytes | 0.1 μg/μL in 100 μL of DPBS via tail vein injection, three times a week for 4 weeks | ↑TLR2, functional recovery, GFAP, 5-HT, BMS score, motor function, and neuron survival; ↓CSPGs deposition, p-PI3K/PI3K, and NF-κB; ↓p-PI3K/PI3K and NF-κB | [92] |
| Primary SCDEs | _____ | SCI | in vivo: SCI induced in SD rats in vitro: H2O2-induced injury in PC12 cells | 250 µL (0.1 µg/µL) in DPBS via tail vein injection, three times a week for 4 weeks | ↑Autophagy, motor function, myelinated areas, NeuN, ChAT, LC3-1/2 and Beclin-1, P62; ↓Apoptosis, %cavity size and EGFR; ↑Neuronal survival, LC3-1/2, Beclin-1 and P62; ↓Apoptosis, EGFR, p-Akt, and p-mTOR | [93] |
| HUSC | ANGPTL3 | SCI | in vivo: SCI induced in mice in vitro: HUVECs | 200 μg in 200 μL of PBS via local intrathecal injection 200 μg | ↑Spinal cord regeneration, locomotor function, BMS scores, sensory improvement, MEP amplitudes, angiogenesis, vessel volume fraction, vascular segment and bifurcation numbers; ↓Latent period and lesion cavities area; ↑Proliferation rate, cell migration, tube formation, angiogenic activities, p-Akt, and p-PI3K | [94] |
| hUC-MSCs | miR-199a-3p/145-5p | SCI | in vivo: SCI induced in SD rats in vitro: LPS-induced injury in PC12 cells | 20 μg/mL | ↑TrkA, locomotor function, and BBB score; ↓Lesion size, inflammation, and apoptotic cells ↑Cell viability, NF-H, β-tubulin-III, Neu-N, p-Akt, p-Erk, neurite outgrowth, and TrkA; ↓Cblb and Cbl expression; | [95] |
| PMs | _____ | SCI | in vivo: SCI induced in SD rats in vitro: BV2 cells cultured in DMEM culture medium | 20 and 200 μg/mL via tail vein injection 20 and 200 μg/mL | ↑BBB score, inclined plate angle, Nissl-positive cells, IL-4, IL-10, and IL-13; ↓Tissue damage, IL-1β, IL-6, and TNF-α; ↑IL-10, CD206, CD163, Arg-1, LC3-II/Ⅰ, and Beclin-1; ↓p62, mTOR, and Akt protein level; | [96] |
| Primary microglia | Resveratrol | SCI | in vivo: SCI induced in SD rats in vitro: Mechanical injury model in primary spinal cord neurons | 0.2 mL of PBS suspension of 40 µM for 14 days | ↑Muscle tension, foot functional movements, BBB scores, motor function, neuron natural morphology and number, LC3B-positive cells, Beclin-1, and p-PI3K; ↓TUNEL-positive neurons, cleaved caspase-3, and apoptosis; | [97] |
| Pericytes | _____ | SCI | in vivo: SCI induced in ICR mice in vitro: SCMECs hypoxic model | 20 µg in 0.3 mL PBS via tail vein injection 20 µg/mL | ↑Locomotor function, complete tissue structure, myelin sheath, Nissl body morphology and number, blood flow, Bcl-2, local microvascular disturbances, mean flux, and claudin-5; ↓Inflammatory cell infiltration, TUNEL-positive cells, Bax, HIF-1α, BSCB disruption, edema formation, MMP-2, and AQP4; ↑ZO-1 and p-Akt; ↓PTEN; | [98] |
| ECFCs | _____ | TBI | in vivo: TBI induced in male C57BL/6 mice in vitro: RBMEC hypoxia injury model | 4×106 cell equivalents via the tail vein at 2 h after TBI,5 and 10 μg/mL | ↑Neurological functional recovery, TJ, p-Akt, CLN5, occludin, and ZO-1; ↓Brain edema, PTEN, MMP-9, BBB permeability, EB dye extravasation, and TJ degradation | [101] |
| Microglia | miR-124-3p | TBI | in vivo: (r)TBI male C57BL/6 mouse model in vitro: Scratch injury model in pure cortical neurons | 30 μg via tail vein, 3 × 108 exosomes | ↑Neurologic outcomes, neurite outgrowth and IL-10; ↓PDE4B, IL-1β, IL-6, TNF-α, mTOR signaling, neuronal inflammation, PDE4B, p-4E-BP1, and p-P70S6K; | [102] |
| Human ADSC | _____ | TBI | in vivo: Weight-drop-induced TBI in male SD rats in vitro: LPS-induced inflammatory model | 20 μg total protein per rat, 2.0 × 1010 particles/mL, intracerebroventricular injection | ↑Functional recovery, neurogenesis, M1 to M2 microglial polarization, IGF1, arginase1, CD206 and IL-10; ↓Neuroinflammation, neuronal apoptosis, hippocampal neurogenesis, CD68+ activated microglia/macrophages, mNSS score, TNF-α, iNOS, IL-1α, IL-1β, IL-6, MCP-1, CCL2, CCL3, CCL5, morphological transformation, p-P38, p-IKKαβ, p-IKBα, p-P68, NF-κB, and P38/MAPK activation | [103] |
| Source of Exosomes | Cargo or Intermediate Molecule | Disease | Method (In Vitro/In Vivo) | Dose/Route of Administration | Mechanism of Actions and Outcomes | References |
|---|---|---|---|---|---|---|
| ADSC | _____ | ALS | in vitro: H2O2-induced injury in NSC-34 (G93A) cells | 0.2 µg/mL, corresponding to 6–8 × 105 particles/mL | ↑Phospho-Akt, SOD1 gene, Bcl-2 α, and cell viability; ↓Cleaved caspase 3, Bax, and apoptosis; | [108] |
| Fibroblast | _____ | CNS injury | in vitro: Cultured adult rat DRGs and RGCs | 50 ng/mL | ↑Cell survival, neurite growth, axonal growth, and pS6K; ↓CSPG and Wnt10b; | [109] |
| Fibroblast | miR-673-5p | Peripheral neuron myelinati on | in vivo: One-day-old newborn rats in vitro: Schwann cells | 5 nmol/rat every 2 days via hypodermic injection | ↑Myelin gene expression, mTORC1, SREBP2, Hmgcr, phosphatidylcholine, phosphatidylethanolamine, phosphatidylserine, diacyl glycerol, cholesterol, myelin sheath, myelinated axons, and myelin lamellae; ↓Tsc2 expression; | [110] |
| MSCs | miR-17-92 | Axonal growth | in vitro: Primary cortical neurons under CSPG conditions | 3 × 108/300 µL and 3 × 109/300 µL | ↑Axonal growth, distal axons length, p-mTOR, and p-GSK-3β; ↓PTEN; | [111] |
| ADSCs | _____ | Neural injury | in vitro: LPS-induced injury in SH-SY5Y and BV-2 cells | 50 μg/mL | ↑Cell viability; ↓Neuroinflammation, microglia cells, TNF-α, IL-1β, IL-6, COX-2, iNOS, cytotoxicity, p-P38, p-P65, p-ERK, and p-JNK; | [112] |
| hAMS | _____ | Neural injury | in vitro: Neural injury induced by glutamate in PC12 cells | 100 ng/mL | ↑Cell survival and PI3K/Akt signaling activating; | [113] |
| MSCs | _____ | ONC injury | in vivo: ONC-induced injury in SD rats | 3 × 109/5 μL, intravitreal injection | ↑RGCs survival, IL-10, Bcl-2, and p-Akt; ↓IL-1β, IL-6, IL-8, MCP-1, Bax, TNF-α, cleaved caspase-3, and apoptosis; | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iranpanah, A.; Kooshki, L.; Moradi, S.Z.; Saso, L.; Fakhri, S.; Khan, H. The Exosome-Mediated PI3K/Akt/mTOR Signaling Pathway in Neurological Diseases. Pharmaceutics 2023, 15, 1006. https://doi.org/10.3390/pharmaceutics15031006
Iranpanah A, Kooshki L, Moradi SZ, Saso L, Fakhri S, Khan H. The Exosome-Mediated PI3K/Akt/mTOR Signaling Pathway in Neurological Diseases. Pharmaceutics. 2023; 15(3):1006. https://doi.org/10.3390/pharmaceutics15031006
Chicago/Turabian StyleIranpanah, Amin, Leila Kooshki, Seyed Zachariah Moradi, Luciano Saso, Sajad Fakhri, and Haroon Khan. 2023. "The Exosome-Mediated PI3K/Akt/mTOR Signaling Pathway in Neurological Diseases" Pharmaceutics 15, no. 3: 1006. https://doi.org/10.3390/pharmaceutics15031006