
Gene Electrotransfer Efficiency in 2D and 3D Cancer Cell Models Using Different Electroporation Protocols: A Comparative Study

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Spheroids Formation
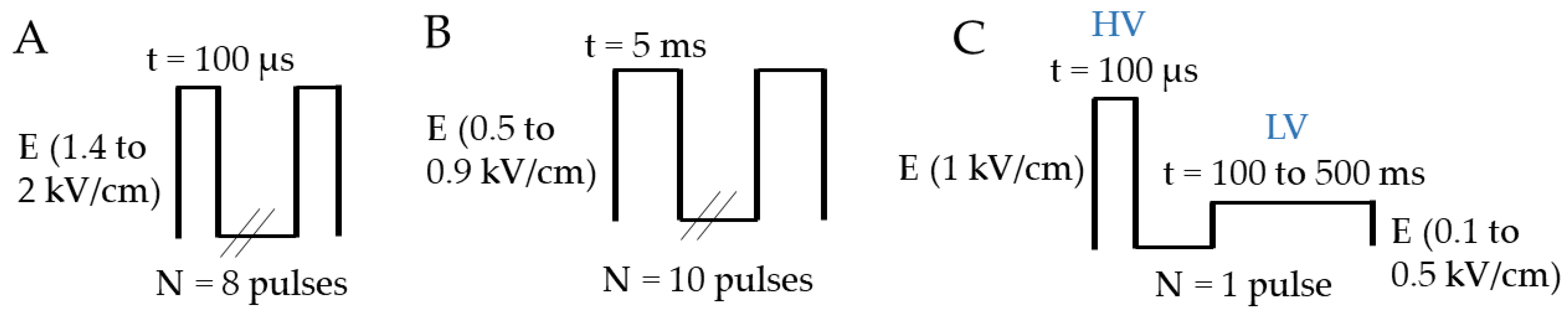
2.2. Electropulsation Device and Electrical Parameters
2.2.1. ECT and GET Device
2.2.2. HV-LV Device
2.3. Electropulsation
2.3.1. Cells in Suspension
2.3.2. Spheroids
2.4. Spheroids Clearing
2.5. Results Analysis
2.5.1. Flow Cytometry
2.5.2. Cell Viability
2.5.3. Fluorescence Microscopy
2.5.4. Biphotonic Microscopy
2.6. Statistics Analysis
3. Results
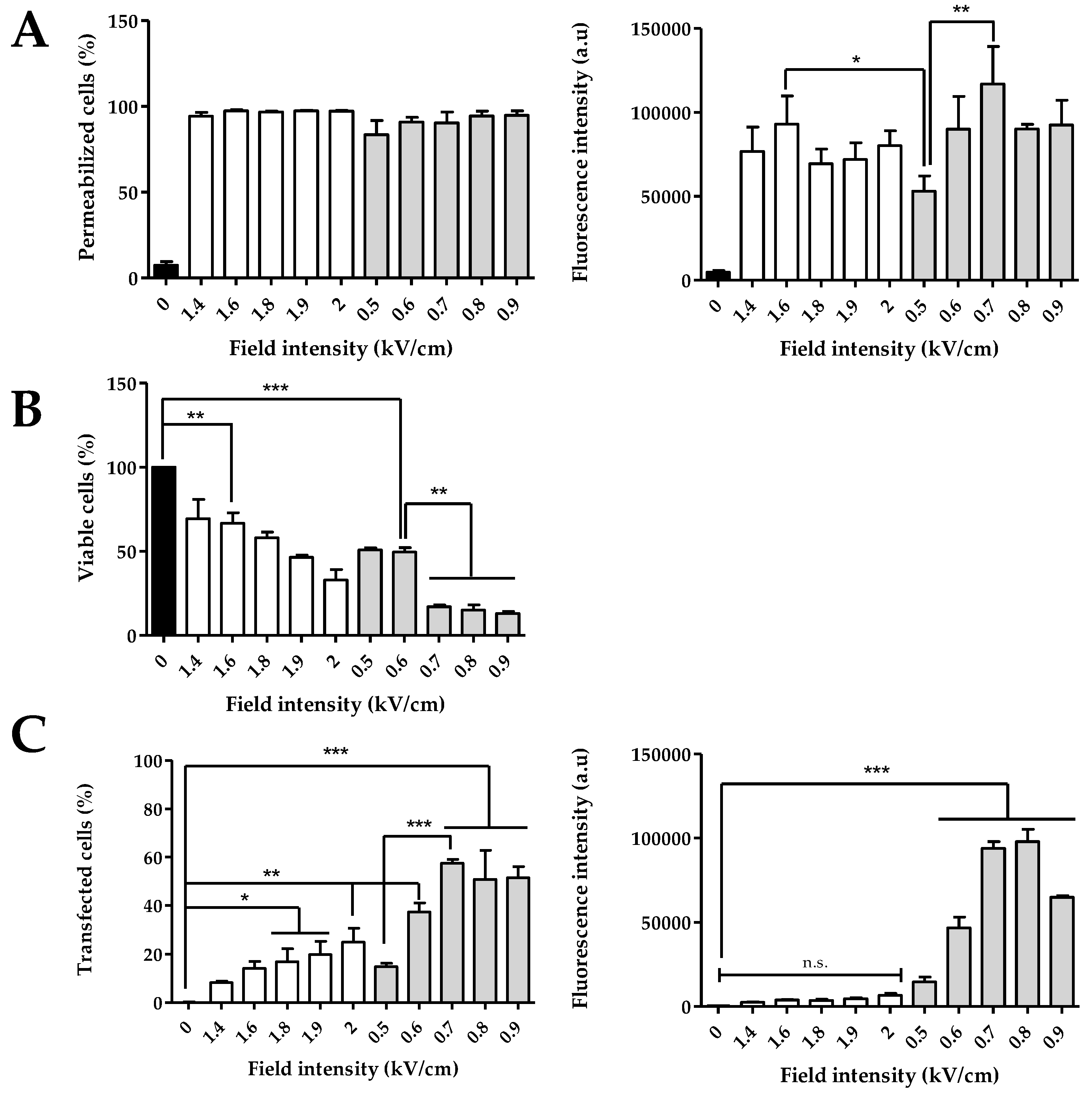
3.1. Effects of ECT and GET Parameters on Cell Permeabilization, Cell Viability and Plasmid Transfection in Cells Suspensions
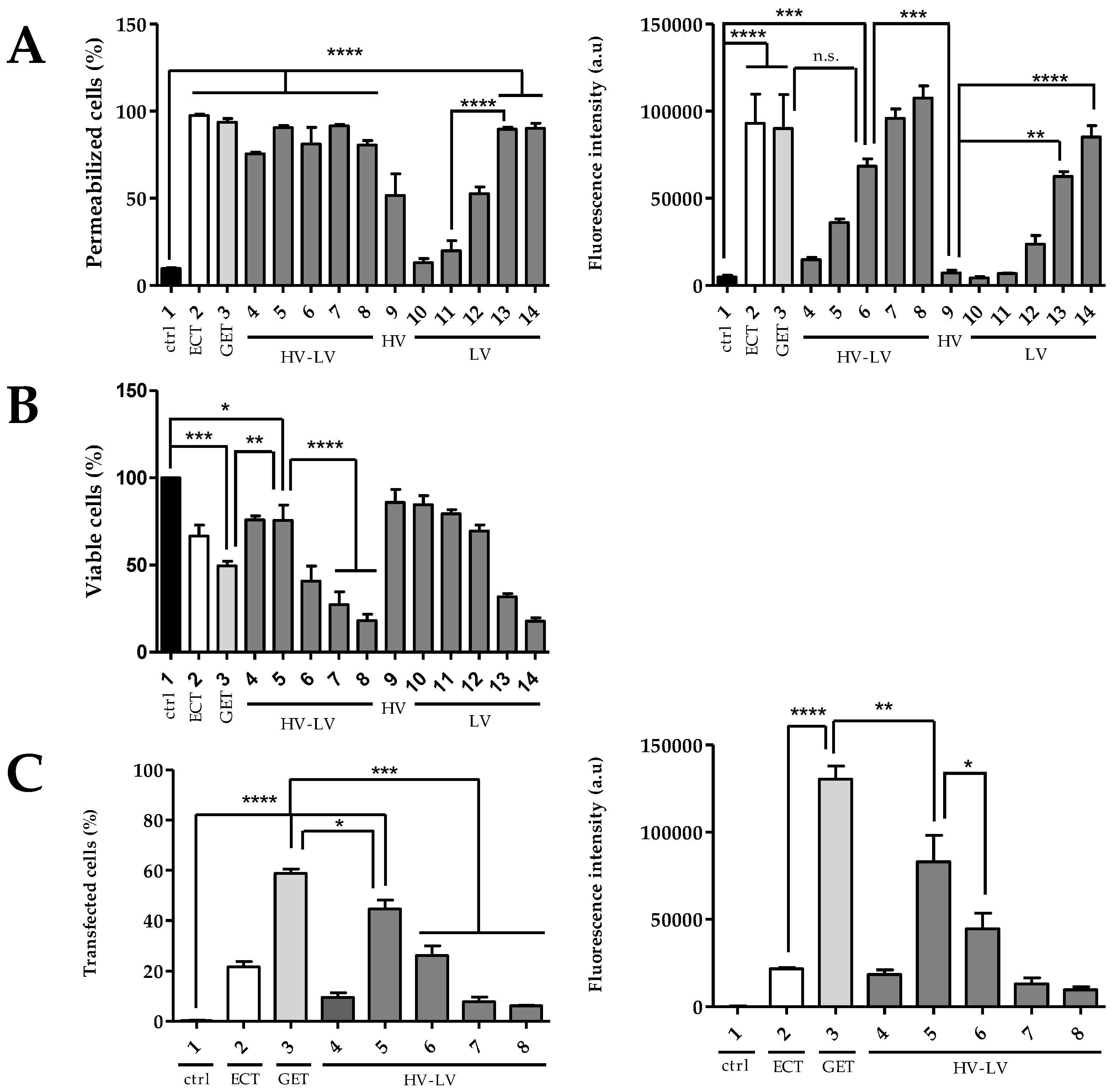
3.2. Effects of HV-LV Parameters on Cell Permeabilization, Cell Viability and Plasmid Transfection in Cells Suspensions
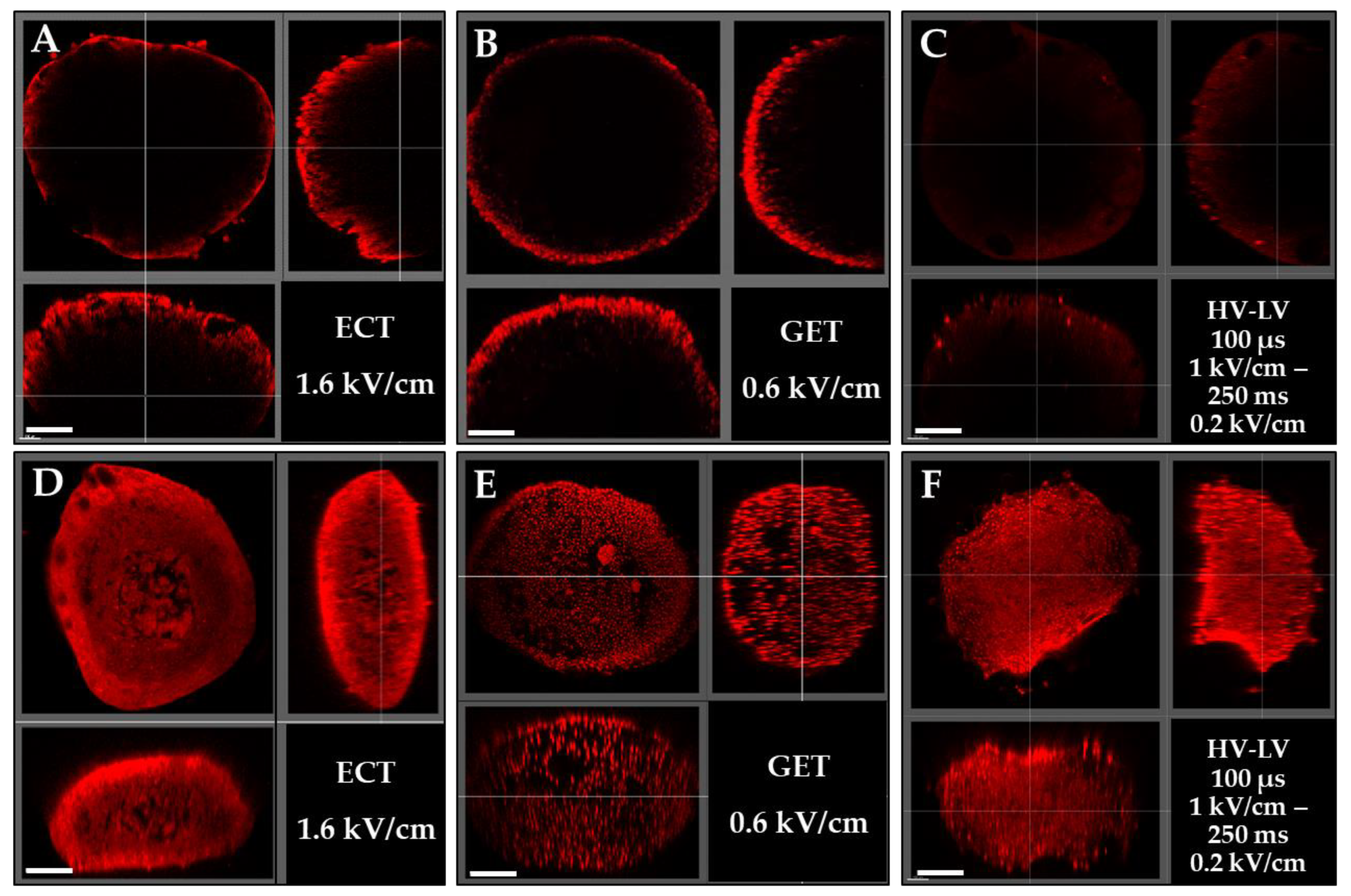
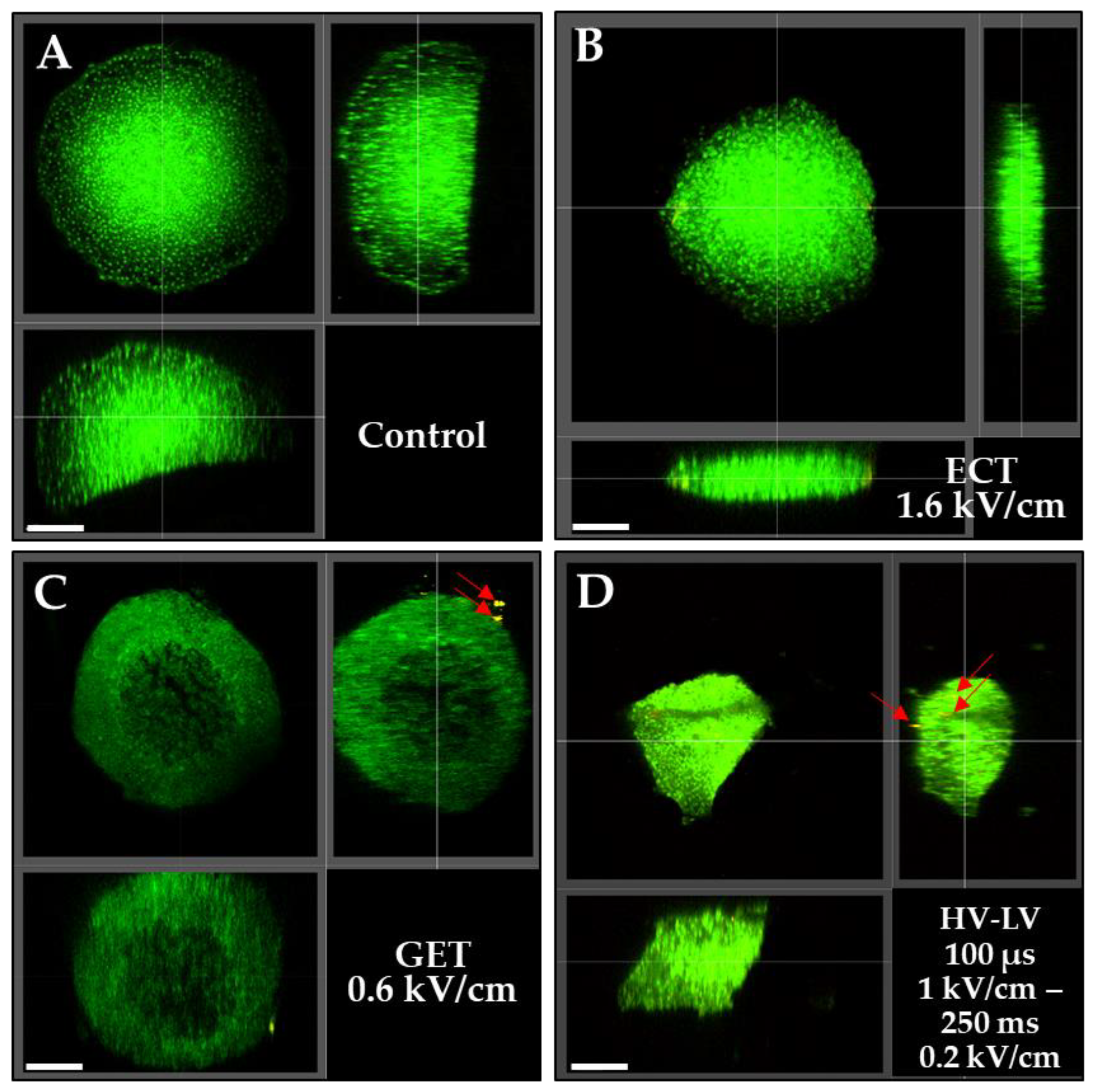
3.3. Effects of ECT, GET, HV-LV Parameters on Permeabilization and Growth of Spheroids Models
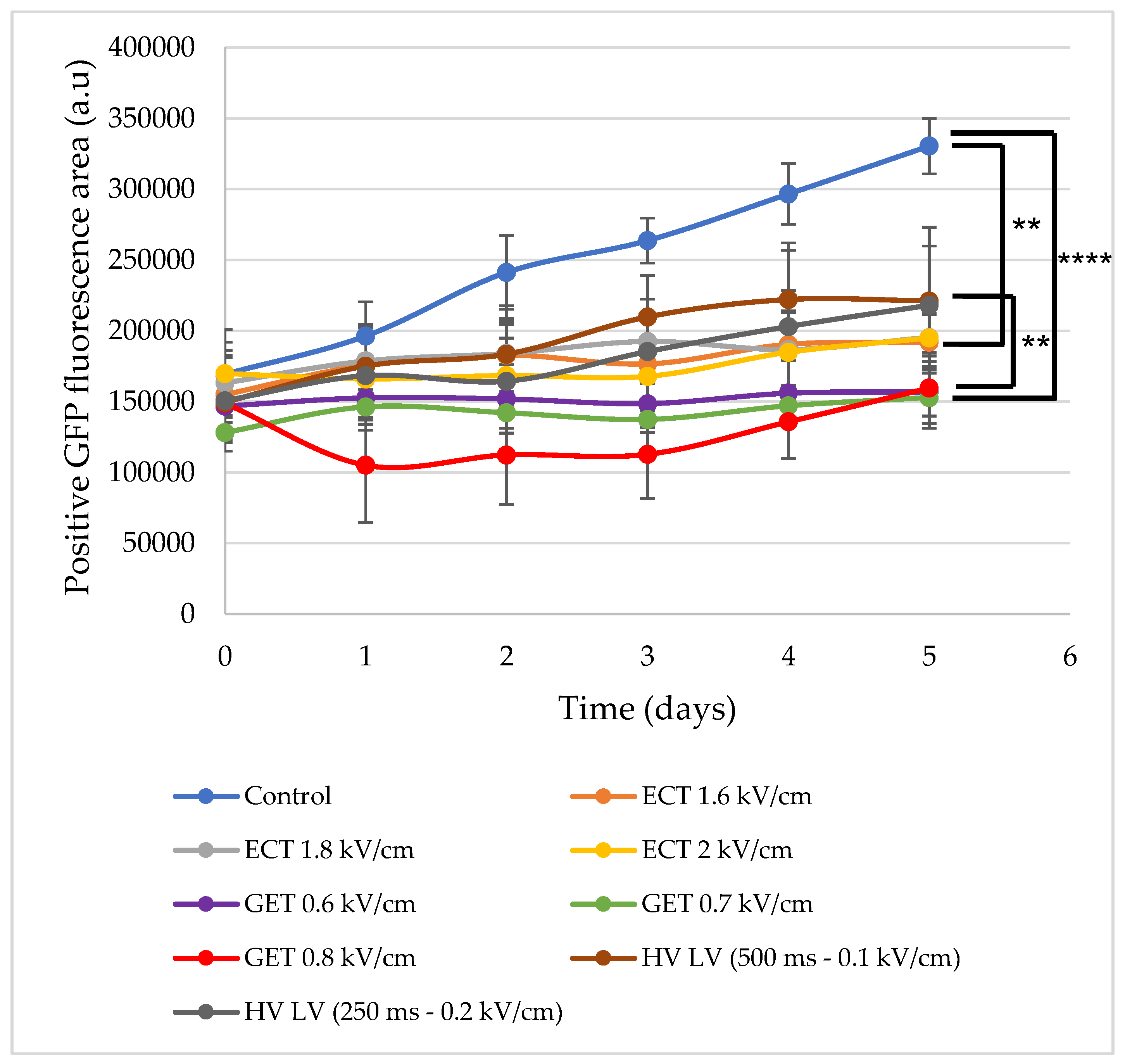
3.4. Effects of ECT, GET, HV-LV Parameters on Plasmid Transfection in Spheroid Models
4. Discussion
5. Conclusions
- (i)
- Transfection of suspended cells is efficient after GET parameters at field values between 0.6 and 0.8 kV/cm, as well as with HV-LV parameters with a LV of 0.2 kV/cm at 250 ms to avoid excessive cell mortality.
- (ii)
- Transfection of spheroids is much less effective and occurs on proliferative external cells. Spheroids are difficult to transfect and are, as such, a relevant model for predicting in vivo outcomes.
- (iii)
- Clearing of spheroids shows that inner cells are efficiently electropermeabilized under all the tested protocols, but not transfected, potentially indicating the problem of plasmid DNA diffusion in tissues and solid tumors. This technique opens new perspectives to study and quantify the delivery of molecules in tissues.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neumann, E.; Schaefer-Ridder, M.; Wang, Y.; Hofschneider, P.H. Gene transfer into mouse lymphoma cells by electroporation in high electric fields. EMBO J. 1982, 1, 841–845. [Google Scholar] [CrossRef]
- Titomirov, A.V.; Sukharev, S.; Kistanova, E. In vivo electroporation and stable transformation of skin cells of newborn mice by plasmid DNA. Biochim. Biophys. Acta 1991, 1088, 131–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, R.; Jaroszeski, M.; Atkin, A.; Moradpour, D.; Gilbert, R.; Wands, J.; Nicolau, C. In vivo gene electroinjection and expression in rat liver. FEBS Lett. 1996, 389, 225–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, L.M.; Bureau, M.F.; Gehl, J.; Rangara, R.; Rouy, D.; Caillaud, J.M.; Delaere, P.; Branellec, D.; Schwartz, B.; Scherman, D. High-efficiency gene transfer into skeletal muscle mediated by electric pulses. Proc. Natl. Acad. Sci. USA 1999, 96, 4262–4267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rols, M.P.; Delteil, C.; Golzio, M.; Dumond, P.; Cros, S.; Teissie, J. In vivo electrically mediated protein and gene transfer in murine melanoma. Nat. Biotechnol. 1998, 16, 168–171. [Google Scholar] [CrossRef]
- Delteil, C.; Teissie, J.; Rols, M.P. Effect of serum on in vitro electrically mediated gene delivery and expression in mammalian cells. Biochim. Biophys. Acta 2000, 1467, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golzio, M.; Teissie, J.; Rols, M.P. Cell synchronization effect on mammalian cell permeabilization and gene delivery by electric field. Biochim. Biophys. Acta 2002, 1563, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faurie, C.; Phez, E.; Golzio, M.; Vossen, C.; Lesbordes, J.C.; Delteil, C.; Teissie, J.; Rols, M.P. Effect of electric field vectoriality on electrically mediated gene delivery in mammalian cells. Biochim. Biophys. Acta 2004, 1665, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Golzio, M.; Teissie, J.; Rols, M.P. Direct visualization at the single-cell level of electrically mediated gene delivery. Proc. Natl. Acad. Sci. USA 2002, 99, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Escoffre, J.M.; Portet, T.; Favard, C.; Teissie, J.; Dean, D.S.; Rols, M.P. Membrane perturbation by an external electric field: A mechanism to permit molecular uptake. Eur. Biophys. J. 2007, 36, 973–983. [Google Scholar] [CrossRef]
- Rols, M.P.; Golzio, M.; Gabriel, B.; Teissie, J. Factors Controlling Electropermeabilisation of Cell Membranes. Technol. Cancer Res. Treat. 2002, 1, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Rosazza, C.; Escoffre, J.M.; Zumbusch, A.; Rols, M.P. The Actin Cytoskeleton Has an Active Role in the Electrotransfer of Plasmid DNA in Mammalian Cells. Mol. Ther. 2011, 19, 913–921. [Google Scholar] [CrossRef]
- Takabatake, Y.; Isaka, Y.; Mizui, M.; Kawachi, M.; Schimizu, F.; Ito, T.; Hori, M.; Emai, E. Exploring RNA interference as a therapeutic strategy for renal disease. Gene Ther. 2005, 12, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Paganin-Gioanni, A.; Bellard, E.; Couderc, B.; Teissei, J.; Golzio, M. Tracking in vitro and in vivo siRNA electrotransfer in tumor cells. J. RNAi Gene Silencing 2008, 4, 281–288. [Google Scholar] [PubMed]
- Paganin-Gioanni, A.; Bellard, E.; Escoffre, J.M.; Rols, M.P.; Teissie, J.; Golzio, M. Direct visualization at the single-cell level of siRNA electrotransfer into cancer cells. Proc. Natl. Acad. Sci. USA 2011, 108, 10443–10447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golzio, M.; Mazzolini, L.; Ledoux, A.; Paganin, A.; Izard, M.; Hellaudais, L.; Bieth, A.; Pillaire, M.J.; Cazaux, C.; Hoffmann, J.C.; et al. In vivo gene silencing in solid tumors by targeted electrically mediated siRNA delivery. Gene Ther. 2007, 14, 752–759. [Google Scholar] [CrossRef] [Green Version]
- Paganin-Gioanni, A.; Rols, M.P.; Teissie, J.; Golzio, M. Cyclin B1 knockdown mediated by clinically approved pulsed electric fields siRNA delivery induces tumor regression in murine melanoma. Int. J. Pharm. 2020, 573, 118732. [Google Scholar] [CrossRef] [PubMed]
- Chabot, S.; Pelofy, S.; Paganin-Gioani, A.; Teissie, J.; Golzio, M. Electrotransfer of RNAi-based Oligonucleotides for Oncology. Anticancer Res. 2011, 31, 4083–4090. [Google Scholar] [PubMed]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.J.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nat. Lett. 2010, 464, 1067–1070. [Google Scholar] [CrossRef] [Green Version]
- Daud, A.I.; DeConti, R.C.; Andrews, S.; Urbas, P.; Riker, A.I.; Sondak, V.K.; Munster, P.N.; Sullivan, D.M.; Ugen, K.E.; Messina, J.L.; et al. Phase I Trial of Interleukin-12 Plasmid Electroporation in Patients With Metastatic Melanoma. J. Clin. Oncol. 2008, 26, 5896–5903. [Google Scholar] [CrossRef] [Green Version]
- Marty, M.; Sersa, G.; Garbay, J.M.; Gehl, J.; Collins, C.G.; Snoj, M.; Billard, V.; Geertsen, P.F.; Larkin, J.O.; Miklavcic, D.; et al. Electrochemotherapy—An easy, highly effective and safe treatment of cutaneous and subcutaneous metastases: Results of ESOPE (European Standard Operating Procedures of Electrochemotherapy) study. EJC Suppl. 2006, 4, 3–13. [Google Scholar] [CrossRef]
- Gehl, J.; Sersa, G.; Matthiessen, L.W.; Muir, T.; Soden, D.; Occhini, A.; Quaglino, P.; Curatolo, P.; Campana, L.G.; Kunte, C.; et al. Updated standard operating procedures for electrochemotherapy of cutaneous tumours and skin metastases. Acta Oncol. 2018, 57, 874–882. [Google Scholar] [CrossRef]
- Suzuki, T.; Shin, B.; Fujikura, K.; Matsuzaki, T.; Takata, K. Direct gene transfer into rat liver cells by in vivo electroporation. FEBS Lett. 1998, 425, 436–440. [Google Scholar] [CrossRef] [Green Version]
- Chopinet, L.; Wasungu, L.; Rols, M.P. First explanations for differences in electrotransfection efficiency in vitro and in vivo using spheroid model. Int. J. Pharm. 2012, 423, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Kandušer, M.; Miklavčic, D.; Pavlin, M. Mechanisms involved in gene electrotransfer using high- and low-voltage pulses—An in vitro study. Bioelectrochemistry 2009, 74, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Pavlin, M.; Flisar, K.; Kanduser, M. The Role of Electrophoresis in Gene Electrotransfer. J. Membr. Biol. 2010, 236, 75–79. [Google Scholar] [CrossRef]
- Orio, J.; Coustets, M.; Mauroy, C.; Teissie, J. Electric Field Orientation for Gene Delivery Using High-Voltage and Low-Voltage Pulses. Membr. Biol. J. 2012, 245, 661–666. [Google Scholar] [CrossRef]
- Pavselj, N.; Preat, V. DNA electrotransfer into the skin using a combination of one high- and one low-voltage pulse. J. Control. Release 2005, 106, 407–415. [Google Scholar] [CrossRef]
- Bureau, M.F.; Gehl, J.; Deleuze, V.; Mir, L.M.; Scherman, D. Importance of association between permeabilization and electrophoretic forces for intramuscular DNA electrotransfer. Biochim. Biophys. Acta 2000, 1474, 353–359. [Google Scholar] [CrossRef]
- Satkauskas, S.; Bureau, M.F.; Puc, M.; Mahfoudi, A.; Scherman, D.; Miklavcic, D.; Mir, L.M. Mechanisms of in Vivo DNA Electrotransfer: Respective Contributions of Cell Electropermeabilization and DNA Electrophoresis. Mol. Ther. 2002, 5, 133–140. [Google Scholar] [CrossRef]
- Tevz, G.; Pavlin, D.; Kamensek, U.; Kranjc, S.; Mesojednik, S.; Coer, A.; Sersa, G.; Cemazar, M. Gene Electrotransfer into Murine Skeletal Muscle: A Systematic Analysis of Parameters for Long-term Gene Expression. Technol. Cancer Res. Treat. 2008, 7, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Meglič, S.H.; Pavlin, M. The impact of impaired DNA mobility on gene electrotransfer efficiency: Analysis in 3D model. BioMed. Eng. OnLine 2021, 20, 85. [Google Scholar] [CrossRef] [PubMed]
- Canatella, P.J.; Black, M.M.; Bonnichsen, D.M.; McKenna, C.; Prausnitz, M.R. Tissue Electroporation: Quantification and Analysis of Heterogeneous Transport in Multicellular Environments. Biophys. J. 2004, 86, 3260–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasungu, L.; Escoffre, J.M.; Valette, A.; Teissie, J.; Rols, M.P. A 3D in vitro spheroid model as a way to study the mechanisms of electroporation. Int. J. Pharm. 2009, 379, 278–284. [Google Scholar] [CrossRef]
- Pelofy, S.; Bousquet, H.; Gibot, L.; Rols, M.P.; Golzio, M. Transfer of small interfering RNA by electropermeabilization in tumor spheroids. Bioelectrochemistry 2021, 141, 107848. [Google Scholar] [CrossRef]
- Pelofy, S.; Teissie, J.; Golzio, M.; Chabot, S. Chemically Modified Oligonucleotide–Increased Stability Negatively Correlates with Its Efficacy Despite Efficient Electrotransfer. J. Membr. Biol. 2012, 245, 565–571. [Google Scholar] [CrossRef]
- Griseti, E.; Kolosnjaj-Tabi, J.; Gibot, L.; Fourquaux, I.; Rols, M.P.; Yousfi, M.; Merbahi, N.; Golzio, M. Pulsed Electric Field Treatment Enhances the Cytotoxicity of Plasma-Activated Liquids in a Three-Dimensional Human Colorectal Cancer Cell Model. Sci. Rep. 2019, 9, 7583. [Google Scholar] [CrossRef] [Green Version]
- Jing, D.; Zhang, S.; Luo, W.; Gao, X.; Men, Y.; Ma, C.; Lui, X.; Yi, Y.; Budge, A.; Zhou, B.O.; et al. Tissue clearing of both hard and soft tissue organs with the PEGASOS method. Cell Res. 2018, 28, 803–818. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.S.; Lichtman, J.W. Clarifying Tissue Clearing. Cell 2015, 162, 246–257. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, R.M. Cell and Environment Interactions in Tumor Microregions: The Multicell Spheroid Model. Science 1998, 240, 177–184. [Google Scholar] [CrossRef]
- Costa, E.C.; Moreira, A.F.; de Melo-Diogo, D.; Correiaa, I.J. Polyethylene glycol molecular weight influences the ClearT2 optical clearing method for spheroids imaging by confocal laser scanning microscopy. J. Biomed. Opt. 2018, 23, 055003. [Google Scholar]
- Santisteban, T.S.; Rabajania, O.; Kalinina, I.; Robinson, S.; Meier, M. Rapid spheroid clearing on a microfluidic chip. Lab Chip 2019, 18, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, E.; Orehov, N.; Tischenko, K.; Schwob, O.; Zamir, G.; Hubert, A.; Manevitch, Z.; Beny, O. Rapid Clearing for High Resolution 3D Imaging of Ex Vivo Pancreatic Cancer Spheroids. Int. J. Mol. Sci. 2020, 21, 7703. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Silva, P.N.; Syed, A.M.; Sindhawani, S.; Rocheleau, J.V.; Chan, W.C. Clarifying intact 3D tissues on a microfluidic chip for high-throughput structural analysis. Proc. Natl. Acad. Sci. USA 2016, 113, 14915–14920. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essay | Number of Pulses | HV | LV | ||
|---|---|---|---|---|---|
| Field Intensity (kV/cm) | Pulse Duration (µs) | Field Intensity (kV/cm) | Pulse Duration (ms) | ||
| 1 | 0 | 0 | 0 | 0 | 0 |
| 2 | 8 | 1.6 | 100 | 0 | 0 |
| 3 | 10 | 0 | 0 | 0.6 | 5 |
| 4 | 1 | 1 | 100 | 0.1 | 500 |
| 5 | 1 | 1 | 100 | 0.2 | 250 |
| 6 | 1 | 1 | 100 | 0.3 | 170 |
| 7 | 1 | 1 | 100 | 0.4 | 125 |
| 8 | 1 | 1 | 100 | 0.5 | 100 |
| 9 | 1 | 1 | 100 | 0 | 0 |
| 10 | 1 | 0 | 0 | 0.1 | 500 |
| 11 | 1 | 0 | 0 | 0.2 | 250 |
| 12 | 1 | 0 | 0 | 0.3 | 170 |
| 13 | 1 | 0 | 0 | 0.4 | 125 |
| 14 | 1 | 0 | 0 | 0.5 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Caro, A.; Bellard, E.; Kolosnjaj-Tabi, J.; Golzio, M.; Rols, M.-P. Gene Electrotransfer Efficiency in 2D and 3D Cancer Cell Models Using Different Electroporation Protocols: A Comparative Study. Pharmaceutics 2023, 15, 1004. https://doi.org/10.3390/pharmaceutics15031004
de Caro A, Bellard E, Kolosnjaj-Tabi J, Golzio M, Rols M-P. Gene Electrotransfer Efficiency in 2D and 3D Cancer Cell Models Using Different Electroporation Protocols: A Comparative Study. Pharmaceutics. 2023; 15(3):1004. https://doi.org/10.3390/pharmaceutics15031004
Chicago/Turabian Stylede Caro, Alexia, Elisabeth Bellard, Jelena Kolosnjaj-Tabi, Muriel Golzio, and Marie-Pierre Rols. 2023. "Gene Electrotransfer Efficiency in 2D and 3D Cancer Cell Models Using Different Electroporation Protocols: A Comparative Study" Pharmaceutics 15, no. 3: 1004. https://doi.org/10.3390/pharmaceutics15031004