
A Gelatin Methacrylate-Based Hydrogel as a Potential Bioink for 3D Bioprinting and Neuronal Differentiation


 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Bioink
2.3. Characterization of the Mechanical Properties of Scaffolds
2.4. Degradation Test
2.5. Rheological Analysis
2.6. Microstructural Analysis
2.7. Biological Characterization
2.8. Extraction and Cultivation of Murine Cortical Astrocytes
2.9. Human Induced Pluripotent Stem Cell (hiPSC) Culture and Human Neural Progenitor Cell (hNPC) Differentiation
2.10. Neuroblastoma Cells (SH-SY5Y)
2.11. hNPC and SH-SY5Y Viability in Bioink
2.12. hNPC and SH-SY5Y Gene Expression in Bioink
2.13. 3D Bioprinting of Murine Cortical Astrocytes and hNPCs
2.14. Neuronal Induction and Maturation
2.15. Immunofluorescence Analysis of Cell Markers
2.16. Statistical Analysis
3. Results
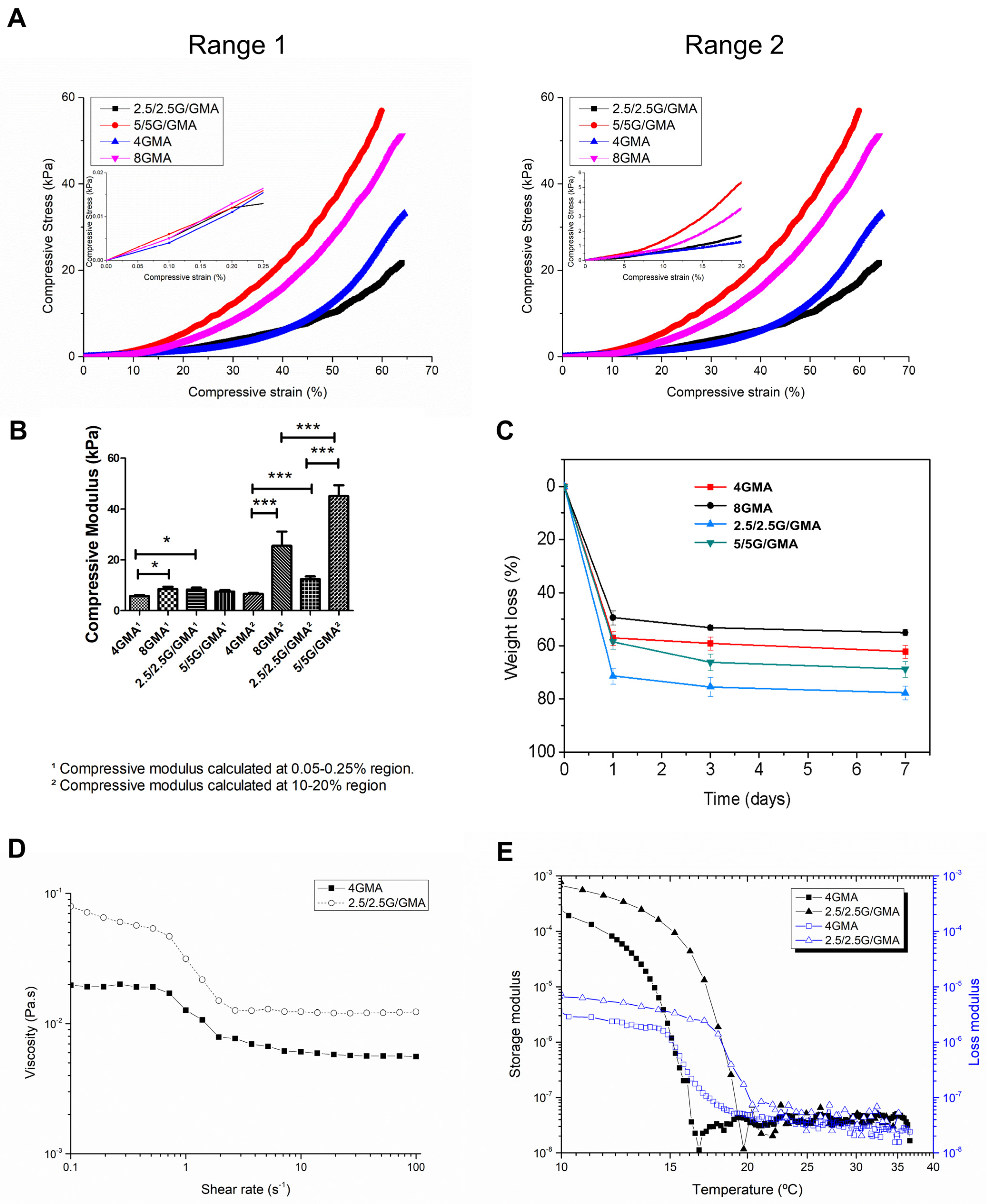
3.1. Mechanical Properties of the Bioink
3.2. Degradation Test
3.3. Rheology Behavior
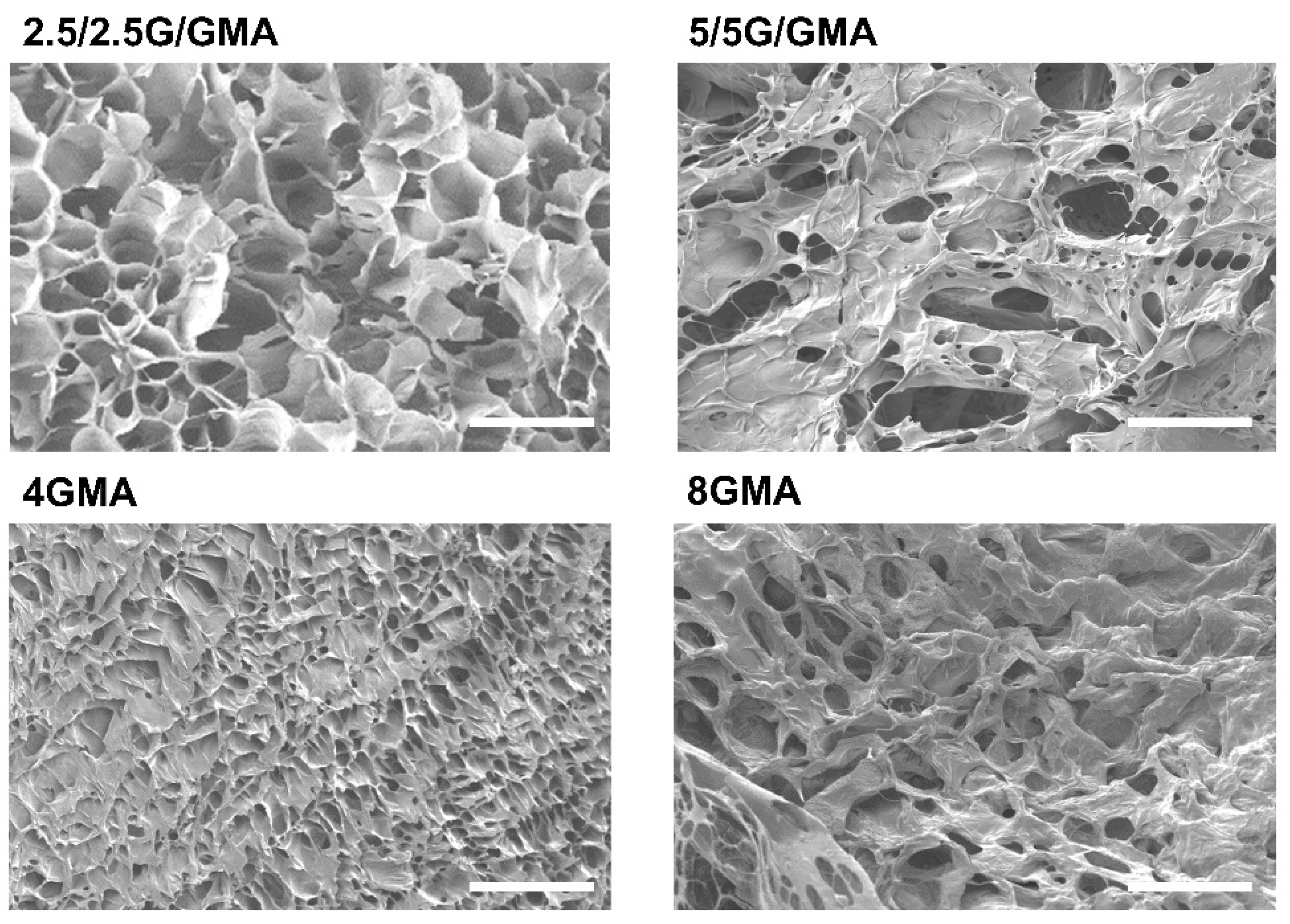
3.4. Bioink Microstructural Analysis by SEM
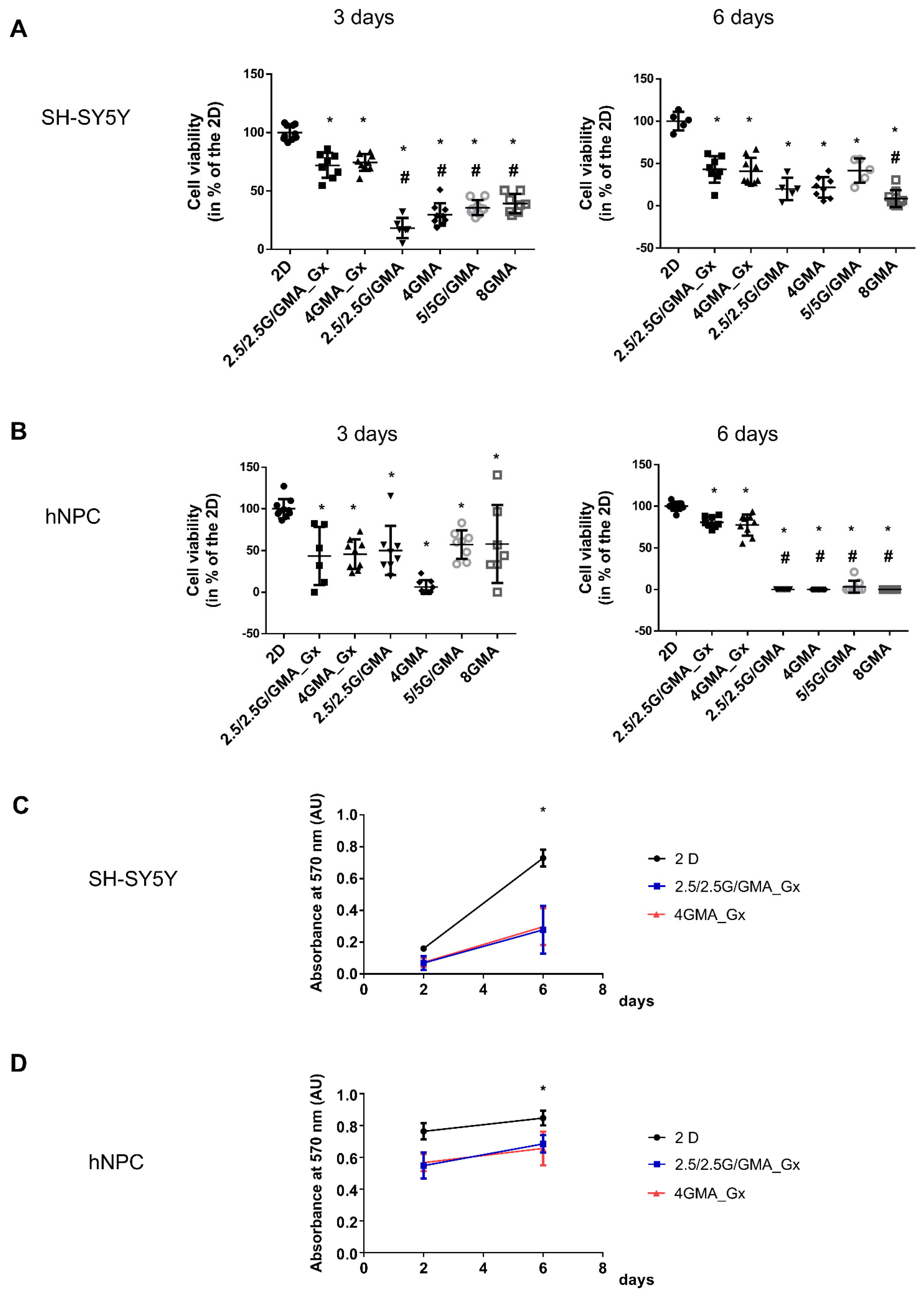
3.5. The Addition of GeltrexTM to the Bioink Is Essential for Maintaining hNPC Viability
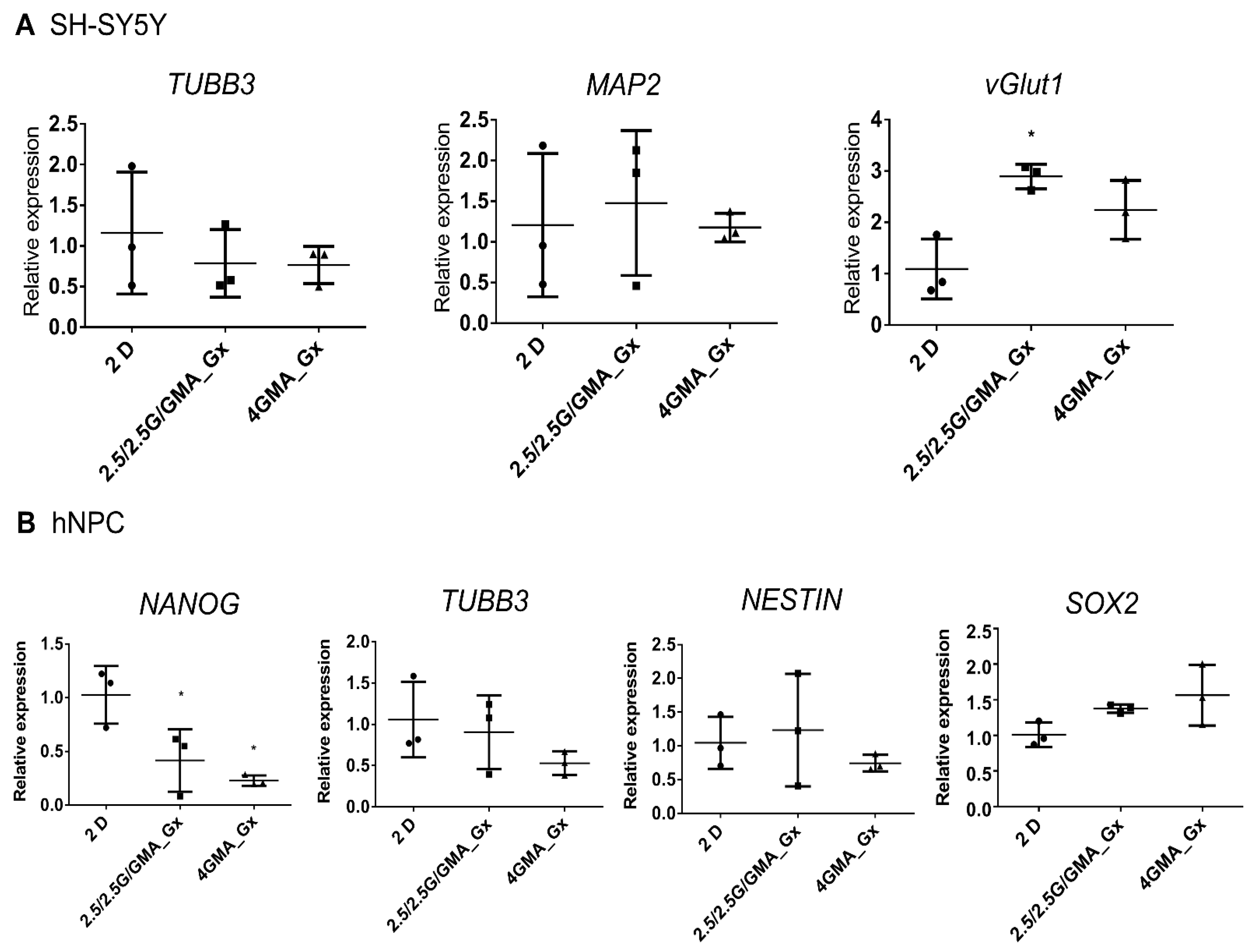
3.6. Cells Cultured in Bioink for Six Days Present Minor Changes in Their Gene Expression Profile
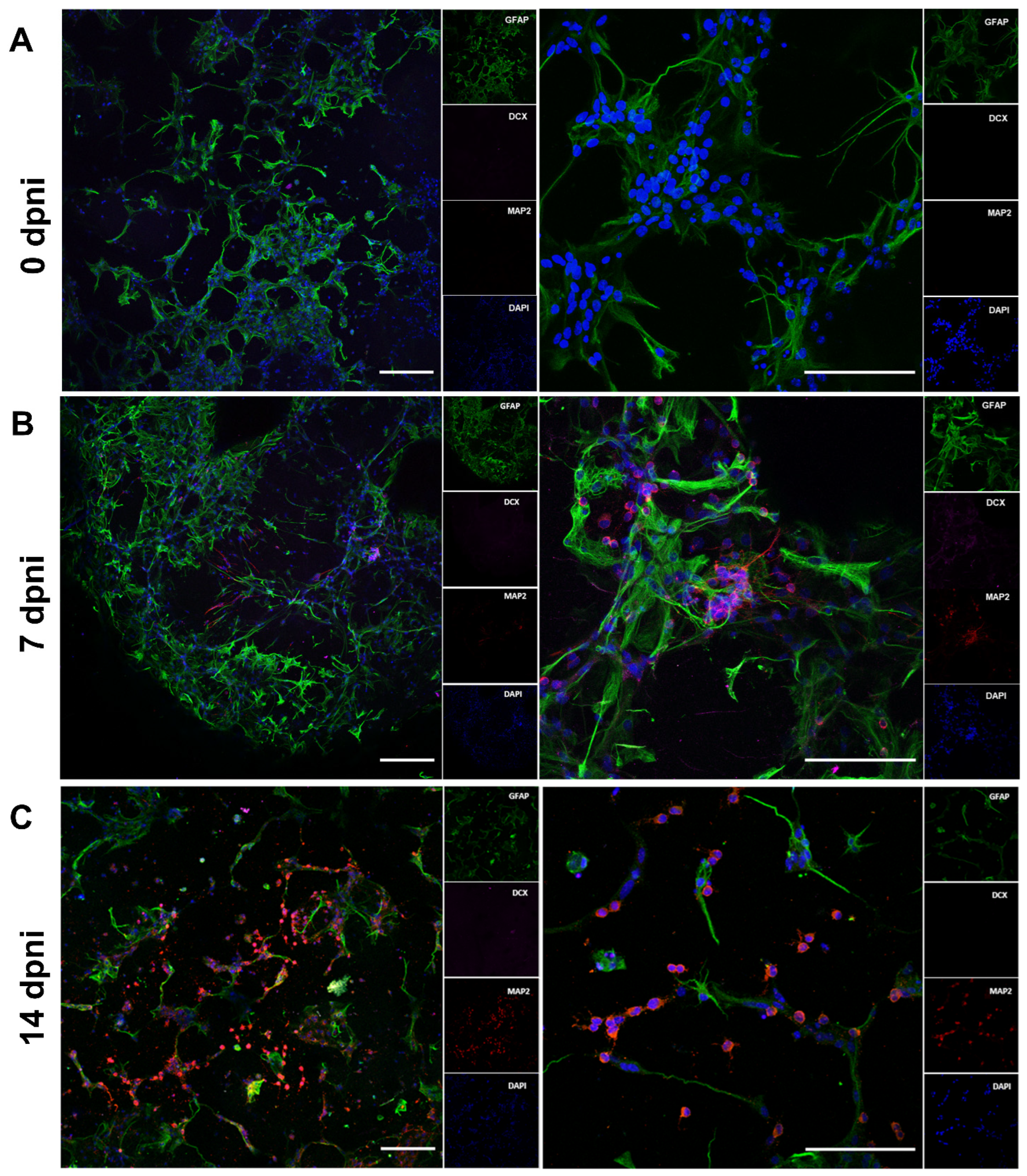
3.7. Bioprinted hNPCs Successfully Differentiated in Immature Neurons
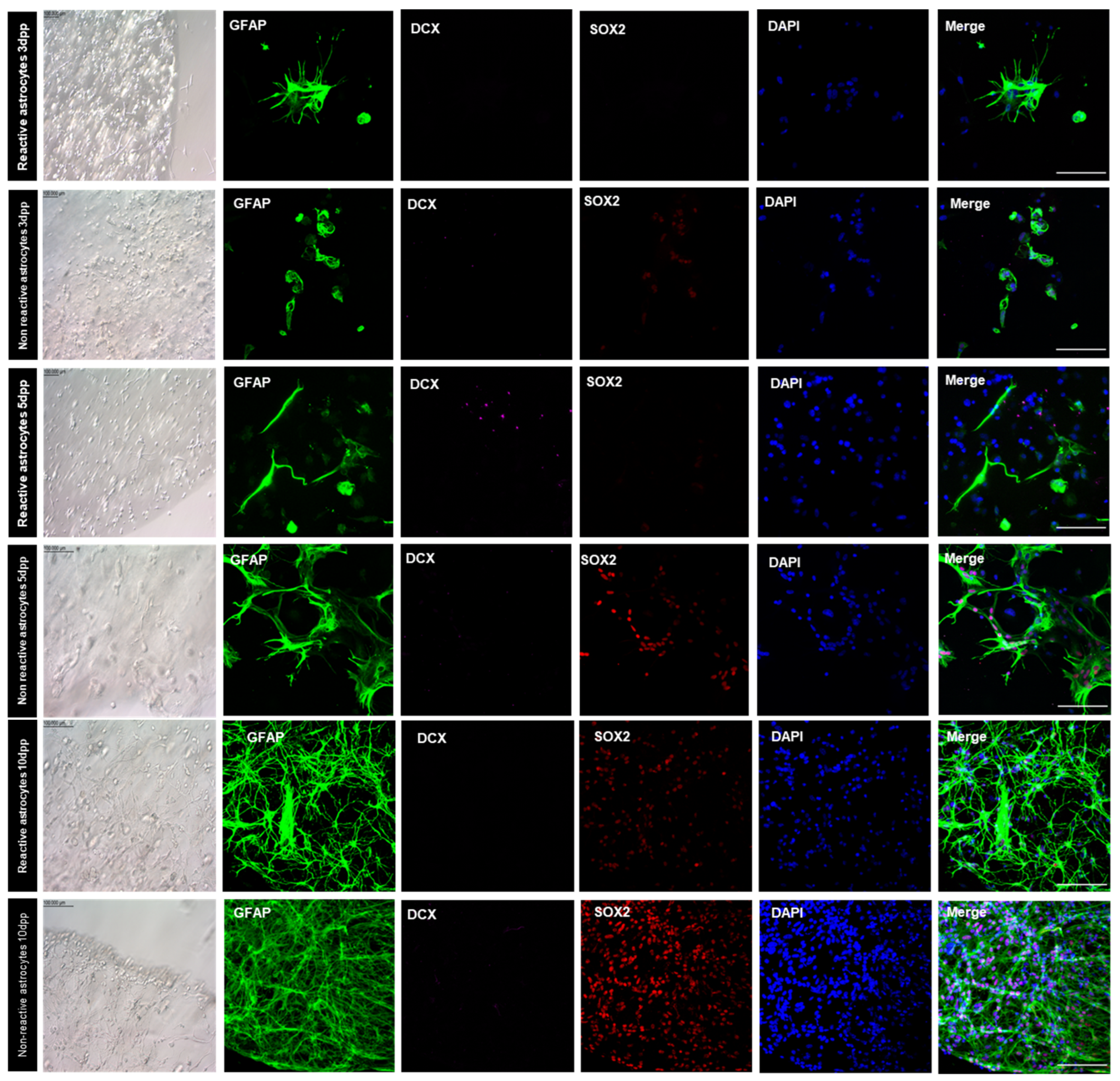
3.8. Bioprinted Murine Astrocytes De-Differentiated in NSC and Originate New Neurons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zamproni, L.N.; Mundim, M.; Porcionatto, M.A. Neurorepair and Regeneration of the Brain: A Decade of Bioscaffolds and Engineered Microtissue. Front. Cell Dev. Biol. 2021, 9, 649891. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, I.; Wood, R.L.; Phillips, C.J.; Macey, S. The costs of traumatic brain injury: A literature review. Clin. Outcomes Res. 2013, 5, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Riggio, S. Traumatic brain injury and its neurobehavioral sequelae. Neurol. Clin. 2011, 29, 35–47. [Google Scholar] [CrossRef]
- Sulhan, S.; Lyon, K.A.; Shapiro, L.A.; Huang, J.H. Neuroinflammation and blood-brain barrier disruption following traumatic brain injury: Pathophysiology and potential therapeutic targets. J. Neurosci. Res. 2020, 98, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Shimazaki, T.; Okano, H. Current understanding of adult neurogenesis in the mammalian brain: How does adult neurogenesis decrease with age? Inflamm. Regen. 2020, 40, 10. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, L. Neural stem cell therapies and hypoxic-ischemic brain injury. Prog. Neurobiol. 2019, 173, 1–17. [Google Scholar] [CrossRef]
- Cheng, A.Y.; García, A.J. Engineering the matrix microenvironment for cell delivery and engraftment for tissue repair. Curr. Opin. Biotechnol. 2013, 24, 864–871. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, C.; Li, J.; Han, J.; Liu, X.; Yang, H. The physical microenvironment of hematopoietic stem cells and its emerging roles in engineering applications. Stem Cell Res. Ther. 2019, 10, 327. [Google Scholar] [CrossRef]
- Krieger, D.W. Therapeutic drug approach to stimulate clinical recovery after brain injury. Front. Neurol. Neurosci. 2013, 32, 76–87. [Google Scholar] [CrossRef]
- Seidi, S.; Eftekhari, A.; Khusro, A.; Heris, R.S.; Sahibzada, M.U.K.; Gajdács, M. Simulation and modeling of physiological processes of vital organs in organ-on-a-chip biosystem. J. King Saud Univ. Sci. 2022, 34, 101710. [Google Scholar] [CrossRef]
- Qiu, B.; Bessler, N.; Figler, K.; Buchholz, M.-B.; Rios, A.C.; Malda, J.; Levato, R.; Caiazzo, M. Bioprinting Neural Systems to Model Central Nervous System Diseases. Adv. Funct. Mater. 2020, 30, 1910250. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.; Guvendiren, M. Recent Advances in Bioink Design for 3D Bioprinting of Tissues and Organs. Front. Bioeng. Biotechnol. 2017, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, F.Y.; Hsu, S.H. 3D bioprinting: A new insight into the therapeutic strategy of neural tissue regeneration. Organogenesis 2015, 11, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Willerth, S.M. 3-D Bioprinting of Neural Tissue for Applications in Cell Therapy and Drug Screening. Front. Bioeng. Biotechnol. 2017, 5, 69. [Google Scholar] [CrossRef]
- Lee, S.-J.; Esworthy, T.; Stake, S.; Miao, S.; Zuo, Y.Y.; Harris, B.T.; Zhang, L.G. Advances in 3D Bioprinting for Neural Tissue Engineering. Adv. Biosyst. 2018, 2, 1700213. [Google Scholar] [CrossRef]
- Huang, C.T.; Kumar Shrestha, L.; Ariga, K.; Hsu, S.H. A graphene-polyurethane composite hydrogel as a potential bioink for 3D bioprinting and differentiation of neural stem cells. J. Mater. Chem. B 2017, 5, 8854–8864. [Google Scholar] [CrossRef]
- Liu, X.; Hao, M.; Chen, Z.; Zhang, T.; Huang, J.; Dai, J.; Zhang, Z. 3D bioprinted neural tissue constructs for spinal cord injury repair. Biomaterials 2021, 272, 120771. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, Y.; Ferracci, G.; Zheng, J.; Cho, N.J.; Lee, B.H. Gelatin methacryloyl and its hydrogels with an exceptional degree of controllability and batch-to-batch consistency. Sci. Rep. 2019, 9, 6863. [Google Scholar] [CrossRef]
- Nichol, J.W.; Koshy, S.T.; Bae, H.; Hwang, C.M.; Yamanlar, S.; Khademhosseini, A. Cell-laden microengineered gelatin methacrylate hydrogels. Biomaterials 2010, 31, 5536–5544. [Google Scholar] [CrossRef]
- Yue, K.; Trujillo-de Santiago, G.; Alvarez, M.M.; Tamayol, A.; Annabi, N.; Khademhosseini, A. Synthesis, properties, and biomedical applications of gelatin methacryloyl (GelMA) hydrogels. Biomaterials 2015, 73, 254–271. [Google Scholar] [CrossRef] [Green Version]
- Discher, D.E.; Janmey, P.; Wang, Y.-l. Tissue Cells Feel and Respond to the Stiffness of Their Substrate. Science 2005, 310, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Kothapalli, C.; Mahajan, G.; Farrell, K. Substrate stiffness induced mechanotransduction regulates temporal evolution of human fetal neural progenitor cell phenotype, differentiation, and biomechanics. Biomater. Sci. 2020, 8, 5452–5464. [Google Scholar] [CrossRef]
- Lv, H.; Li, L.; Sun, M.; Zhang, Y.; Chen, L.; Rong, Y.; Li, Y. Mechanism of regulation of stem cell differentiation by matrix stiffness. Stem Cell Res. Ther. 2015, 6, 103. [Google Scholar] [CrossRef] [PubMed]
- de Melo, B.A.G.; Cruz, E.M.; Ribeiro, T.N.; Mundim, M.V.; Porcionatto, M.A. 3D Bioprinting of Murine Cortical Astrocytes for Engineering Neural-Like Tissue. J. Vis. Exp. 2021, 173, e62691. [Google Scholar] [CrossRef]
- Schildge, S.; Bohrer, C.; Beck, K.; Schachtrup, C. Isolation and culture of mouse cortical astrocytes. J. Vis. Exp. 2013, 71, e50079. [Google Scholar] [CrossRef]
- Ribeiro, T.N.; Delgado-García, L.M.; Porcionatto, M.A. Notch1 and Galectin-3 Modulate Cortical Reactive Astrocyte Response After Brain Injury. Front. Cell Dev. Biol. 2021, 9, 649854. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cheng, X.P.; Li, J.W.; Yao, Q.; Ju, G. De-differentiation response of cultured astrocytes to injury induced by scratch or conditioned culture medium of scratch-insulted astrocytes. Cell Mol. Neurobiol. 2009, 29, 455–473. [Google Scholar] [CrossRef]
- Chambers, S.M.; Fasano, C.A.; Papapetrou, E.P.; Tomishima, M.; Sadelain, M.; Studer, L. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling. Nat. Biotechnol. 2009, 27, 275–280. [Google Scholar] [CrossRef]
- Machado, L.S.; Pieri, N.C.G.; Botigelli, R.C.; de Castro, R.V.G.; de Souza, A.F.; Bridi, A.; Lima, M.A.; Fantinato Neto, P.; Pessôa, L.V.F.; Martins, S.; et al. Generation of neural progenitor cells from porcine-induced pluripotent stem cells. J. Tissue Eng. Regen. Med. 2020, 14, 1880–1891. [Google Scholar] [CrossRef]
- Rasmussen, M.A.; Holst, B.; Tümer, Z.; Johnsen, M.G.; Zhou, S.; Stummann, T.C.; Hyttel, P.; Clausen, C. Transient p53 suppression increases reprogramming of human fibroblasts without affecting apoptosis and DNA damage. Stem Cell Rep. 2014, 3, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Kirwan, P.; Livesey, F.J. Directed differentiation of human pluripotent stem cells to cerebral cortex neurons and neural networks. Nat. Protoc. 2012, 7, 1836–1846. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Samanipour, R.; Tahmooressi, H.; Rezaei Nejad, H.; Hirano, M.; Shin, S.R.; Hoorfar, M. A review on 3D printing functional brain model. Biomicrofluidics 2022, 16, 011501. [Google Scholar] [CrossRef]
- Boularaoui, S.; Al Hussein, G.; Khan, K.A.; Christoforou, N.; Stefanini, C. An overview of extrusion-based bioprinting with a focus on induced shear stress and its effect on cell viability. Bioprinting 2020, 20, e00093. [Google Scholar] [CrossRef]
- Li, Y.-C.E.; Jodat, Y.A.; Samanipour, R.; Zorzi, G.; Zhu, K.; Hirano, M.; Chang, K.; Arnaout, A.; Hassan, S.; Matharu, N.; et al. Toward a neurospheroid niche model: Optimizing embedded 3D bioprinting for fabrication of neurospheroid brain-like co-culture constructs. Biofabrication 2020, 13, 015014. [Google Scholar] [CrossRef] [PubMed]
- Man, A.J.; Davis, H.E.; Itoh, A.; Leach, J.K.; Bannerman, P. Neurite outgrowth in fibrin gels is regulated by substrate stiffness. Tissue Eng. Part A 2011, 17, 2931–2942. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xiang, Y.; Fang, J.; Li, X.; Lin, Z.; Dai, G.; Yin, J.; Wei, P.; Zhang, D. The influence of the stiffness of GelMA substrate on the outgrowth of PC12 cells. Biosci. Rep. 2019, 39, BSR20181748. [Google Scholar] [CrossRef]
- Xu, Z.; Li, Y.; Li, P.; Sun, Y.; Lv, S.; Wang, Y.; He, X.; Xu, J.; Li, L. Soft substrates promote direct chemical reprogramming of fibroblasts into neurons. Acta Biomater. 2022, 152, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Huang, G.; Tian, J.; Qiu, J.; Jia, Y.; Feng, D.; Wei, Z.; Li, S.; Xu, F. Matrix stiffness changes affect astrocyte phenotype in an in vitro injury model. NPG Asia Mater. 2021, 13, 35. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Xu, Q.; Ding, L.; Xia, Y.; Zhang, C.; Lai, H.; Liu, C.; Deng, D.Y.B. Cell Infiltrative Inner Connected Porous Hydrogel Improves Neural Stem Cell Migration and Differentiation for Functional Repair of Spinal Cord Injury. ACS Biomater. Sci. Eng. 2022, 8, 5307–5318. [Google Scholar] [CrossRef]
- Zhou, L.; Wolfes, A.C.; Li, Y.; Chan, D.C.W.; Ko, H.; Szele, F.G.; Bayley, H. Lipid-Bilayer-Supported 3D Printing of Human Cerebral Cortex Cells Reveals Developmental Interactions. Adv. Mater. 2020, 32, e2002183. [Google Scholar] [CrossRef]
- Herrera-Ruiz, A.; Tovar, B.B.; García, R.G.; Tamez, M.F.L.; Mamidi, N. Nanomaterials-Incorporated Chemically Modified Gelatin Methacryloyl-Based Biomedical Composites: A Novel Approach for Bone Tissue Engineering. Pharmaceutics 2022, 14, 2645. [Google Scholar] [CrossRef] [PubMed]
- Sakr, M.A.; Sakthivel, K.; Hossain, T.; Shin, S.R.; Siddiqua, S.; Kim, J.; Kim, K. Recent trends in gelatin methacryloyl nanocomposite hydrogels for tissue engineering. J. Biomed. Mater. Res. A 2022, 110, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Rajabi, N.; Rezaei, A.; Kharaziha, M.; Bakhsheshi-Rad, H.R.; Luo, H.; RamaKrishna, S.; Berto, F. Recent Advances on Bioprinted Gelatin Methacrylate-Based Hydrogels for Tissue Repair. Tissue Eng. Part A 2021, 27, 679–702. [Google Scholar] [CrossRef]
- Lopez-Suarez, L.; Awabdh, S.A.; Coumoul, X.; Chauvet, C. The SH-SY5Y human neuroblastoma cell line, a relevant in vitro cell model for investigating neurotoxicology in human: Focus on organic pollutants. NeuroToxicology 2022, 92, 131–155. [Google Scholar] [CrossRef] [PubMed]
- Salaris, F.; Colosi, C.; Brighi, C.; Soloperto, A.; Turris, V.; Benedetti, M.C.; Ghirga, S.; Rosito, M.; Di Angelantonio, S.; Rosa, A. 3D Bioprinted Human Cortical Neural Constructs Derived from Induced Pluripotent Stem Cells. J. Clin. Med. 2019, 8, 1595. [Google Scholar] [CrossRef] [PubMed]
- Keung, A.J.; Dong, M.; Schaffer, D.V.; Kumar, S. Pan-neuronal maturation but not neuronal subtype differentiation of adult neural stem cells is mechanosensitive. Sci. Rep. 2013, 3, 1817. [Google Scholar] [CrossRef]
- Du, X.; Li, J.; Li, M.; Yang, X.; Qi, Z.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. Research progress on the role of type I vesicular glutamate transporter (VGLUT1) in nervous system diseases. Cell Biosci. 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; George, J.K.; Sorger, V.J.; Grace Zhang, L. 3D printing scaffold coupled with low level light therapy for neural tissue regeneration. Biofabrication 2017, 9, 025002. [Google Scholar] [CrossRef]
- Gu, Y.; Cheng, X.; Huang, X.; Yuan, Y.; Qin, S.; Tan, Z.; Wang, D.; Hu, X.; He, C.; Su, Z. Conditional ablation of reactive astrocytes to dissect their roles in spinal cord injury and repair. Brain Behav. Immun. 2019, 80, 394–405. [Google Scholar] [CrossRef]
- Sharma, R.; Smits, I.P.M.; De La Vega, L.; Lee, C.; Willerth, S.M. 3D Bioprinting Pluripotent Stem Cell Derived Neural Tissues Using a Novel Fibrin Bioink Containing Drug Releasing Microspheres. Front. Bioeng. Biotechnol. 2020, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Cui, H.; Nowicki, M.; Miao, S.; Lee, S.J.; Masood, F.; Harris, B.T.; Zhang, L.G. Three-Dimensional-Bioprinted Dopamine-Based Matrix for Promoting Neural Regeneration. ACS Appl. Mater. Interfaces 2018, 10, 8993–9001. [Google Scholar] [CrossRef]
- Yang, H.; Qian, X.H.; Cong, R.; Li, J.W.; Yao, Q.; Jiao, X.Y.; Ju, G.; You, S.W. Evidence for heterogeneity of astrocyte de-differentiation in vitro: Astrocytes transform into intermediate precursor cells following induction of ACM from scratch-insulted astrocytes. Cell Mol. Neurobiol. 2010, 30, 483–491. [Google Scholar] [CrossRef]
- Zhang, S.; Cui, W. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.L.; Tian, F.; Zhang, S.Q.; Zhang, H.; Yin, Z.S. Epidermal growth factor increases the expression of Nestin in rat reactive astrocytes through the Ras-Raf-ERK pathway. Neurosci. Lett. 2014, 562, 54–59. [Google Scholar] [CrossRef]
- Kleiderman, S.; Gutbier, S.; Ugur Tufekci, K.; Ortega, F.; Sá, J.V.; Teixeira, A.P.; Brito, C.; Glaab, E.; Berninger, B.; Alves, P.M.; et al. Conversion of Nonproliferating Astrocytes into Neurogenic Neural Stem Cells: Control by FGF2 and Interferon-γ. Stem Cells 2016, 34, 2861–2874. [Google Scholar] [CrossRef]
- Puschmann, T.B.; Zandén, C.; Lebkuechner, I.; Philippot, C.; de Pablo, Y.; Liu, J.; Pekny, M. HB-EGF affects astrocyte morphology, proliferation, differentiation, and the expression of intermediate filament proteins. J. Neurochem. 2014, 128, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.-D.; He, B.-R.; Lu, F.; Liu, L.-H.; Zhang, L.; Chen, B.; He, Z.-P.; Hao, D.-J.; Yang, H. Fibroblast Growth Factor 4 Is Required but not Sufficient for the Astrocyte Dedifferentiation. Mol. Neurobiol. 2014, 50, 997–1012. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioink | Acronym |
|---|---|
| 2.5/2.5 wt.% Gelatin/GelMA | 2.5/2.5G/GMA |
| 5/5 wt.% Gelatin/GelMA | 5/5G/GMA |
| 4.0 wt.% GelMA | 4GMA |
| 8.0 wt.% GelMA | 8GMA |
| 1:1 (v/v) GeltrexTM + 5/5 wt.% Gelatin/GelMA * | 2.5/2.5G/GMA_Gx |
| 1:1 (v/v) GeltrexTM + 8.0 wt.% GelMA ** | 4GMA_Gx |
| Gene | Sequence | Forward | Reverse |
|---|---|---|---|
| GAPDH | NM_001256799.3 | GTGGTCTCCTCTGACTTCAAC | CCTGTTGCTGTAGCCAAATTC |
| ACTB | NM_001101.5 | TCCACGAAACTACCTTCAACTC | CAGTGATCTCCTTCTGCATCC |
| SOX2 | NM_003106.4 | TACAGCATGTCCTACTCGCAG | GAGGAAGAGGTAACCACAGGG |
| NESTIN | NM_006617.2 | AGAGAGCGTAGAGGCAGTAA | GGTGCTTGAGTTTCTGGAGAT |
| NANOG | NM_001297698.2 | GCAAATGTCTTCTGCTGAGATG | CTTTGGGACTGGTGGAAGAA |
| TUBB3 | NM_001197181.2 | AGTATCCCGACCGCATCAT | AGTAGGTCTCATCCGTGTTCTC |
| MAP2 | NM_001039538.2 | TGGTGCCGAGTGAGAAGAAG | AGTGGTTGGTTAATAAGCCGAAG |
| vGlut1 | NM_020309.4 | CGACGACAGCCTTTTGTGGT | GCCGTAGACGTAGAAAACAGAG |
| Bioink | Compressive Modulus (kPa) |
|---|---|
| 4.0 wt.% GelMA 1 | 5.8 ± 0.9 |
| 8.0 wt.% GelMA 1 | 8.5 ± 1.8 |
| 2.5/2.5 wt.% Gelatin/GelMA 1 | 8.3 ± 1.6 |
| 5/5 wt.% Gelatin/GelMA 1 | 7.5 ± 1.3 |
| 4.0 wt.% GelMA 2 | 6.6 ± 0.9 |
| 8.0 wt.% GelMA 2 | 25.6 ± 2.3 |
| 2.5/2.5 wt.% Gelatin/GelMA 2 | 12.4 ± 2.3 |
| 5/5 wt.% Gelatin/GelMA 2 | 45.2 ± 8.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, E.M.; Machado, L.S.; Zamproni, L.N.; Bim, L.V.; Ferreira, P.S.; Pinto, L.A.; Pessan, L.A.; Backes, E.H.; Porcionatto, M.A. A Gelatin Methacrylate-Based Hydrogel as a Potential Bioink for 3D Bioprinting and Neuronal Differentiation. Pharmaceutics 2023, 15, 627. https://doi.org/10.3390/pharmaceutics15020627
Cruz EM, Machado LS, Zamproni LN, Bim LV, Ferreira PS, Pinto LA, Pessan LA, Backes EH, Porcionatto MA. A Gelatin Methacrylate-Based Hydrogel as a Potential Bioink for 3D Bioprinting and Neuronal Differentiation. Pharmaceutics. 2023; 15(2):627. https://doi.org/10.3390/pharmaceutics15020627
Chicago/Turabian StyleCruz, Elisa Marozzi, Lucas Simões Machado, Laura Nicoleti Zamproni, Larissa Valdemarin Bim, Paula Scanavez Ferreira, Leonardo Alves Pinto, Luiz Antonio Pessan, Eduardo Henrique Backes, and Marimélia Aparecida Porcionatto. 2023. "A Gelatin Methacrylate-Based Hydrogel as a Potential Bioink for 3D Bioprinting and Neuronal Differentiation" Pharmaceutics 15, no. 2: 627. https://doi.org/10.3390/pharmaceutics15020627