
RNAi-Mediated Knockdown of Cottontail Rabbit Papillomavirus Oncogenes Using Low-Toxicity Lipopolyplexes as a Paradigm to Treat Papillomavirus-Associated Cancers
 , , ,
, , ,  , , and
, , and 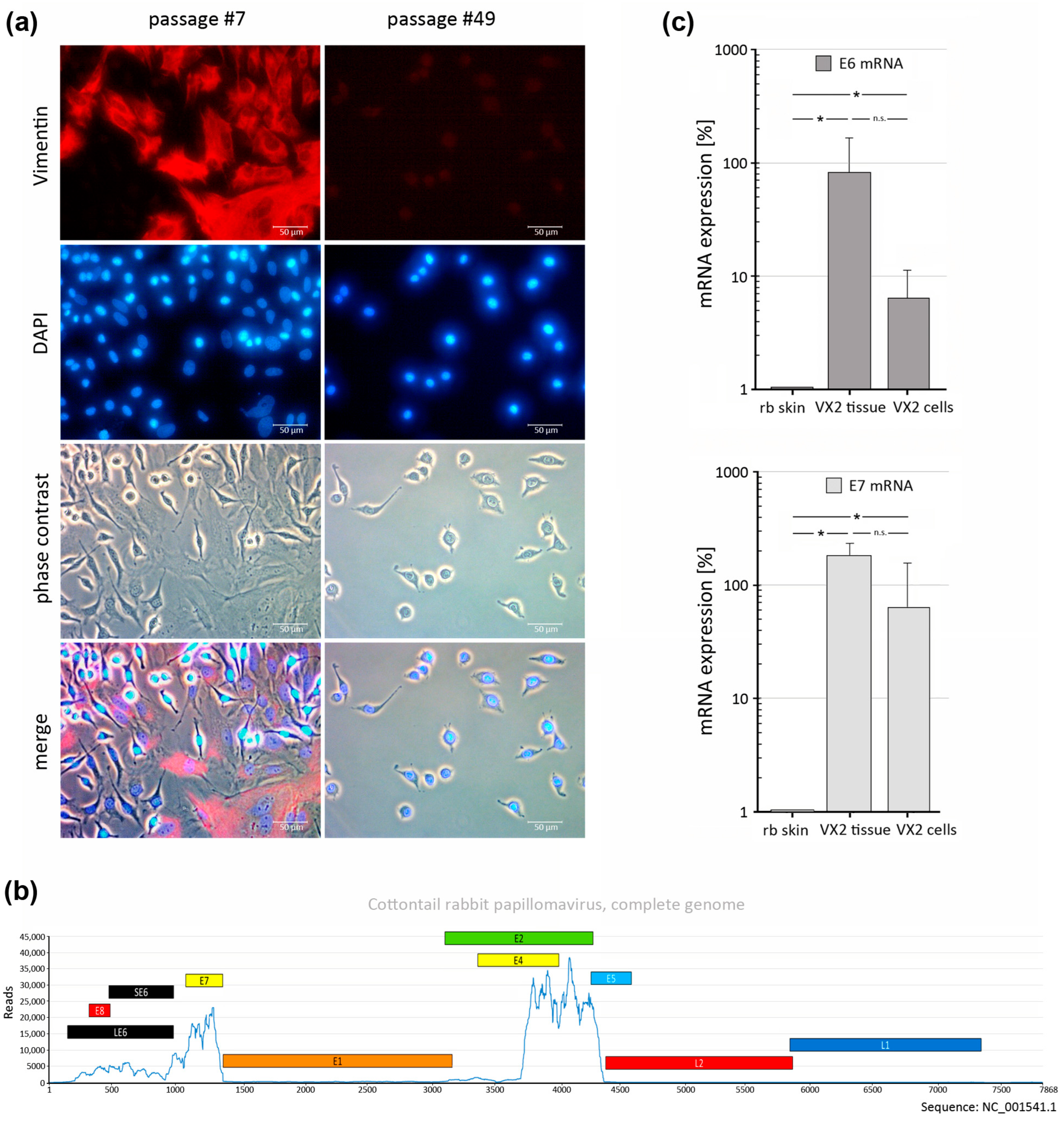{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Preparation of Lipopolyplexes
2.3. RT-qPCR Detection of CRPV E6 and E7 Transcripts
2.4. Western Blot Analysis
2.5. Generation of CRPV E6 and E7 Expression Plasmids
2.6. Expression of CRPV E6 and E7 in VX2 and COS-7 Cells
2.7. MTT Cell Viability Assay
2.8. RNAi Knockdown of CRPV E6 and E7 by siRNA-Loaded Lipopolyplexes
2.9. Measuring CRPV E6 and E7 Knockdown in COS-7 Cells by Flow Cytometry
2.10. Wound Healing Assay
2.11. Real Time Cellular Analysis (RTCA)
2.12. Flow Cytometry and Cell Cycle Analysis of VX2 Cells
2.13. Statistical Analysis
3. Results and Discussion
3.1. Generation of a VX2-Tumor-Derived Cell Line
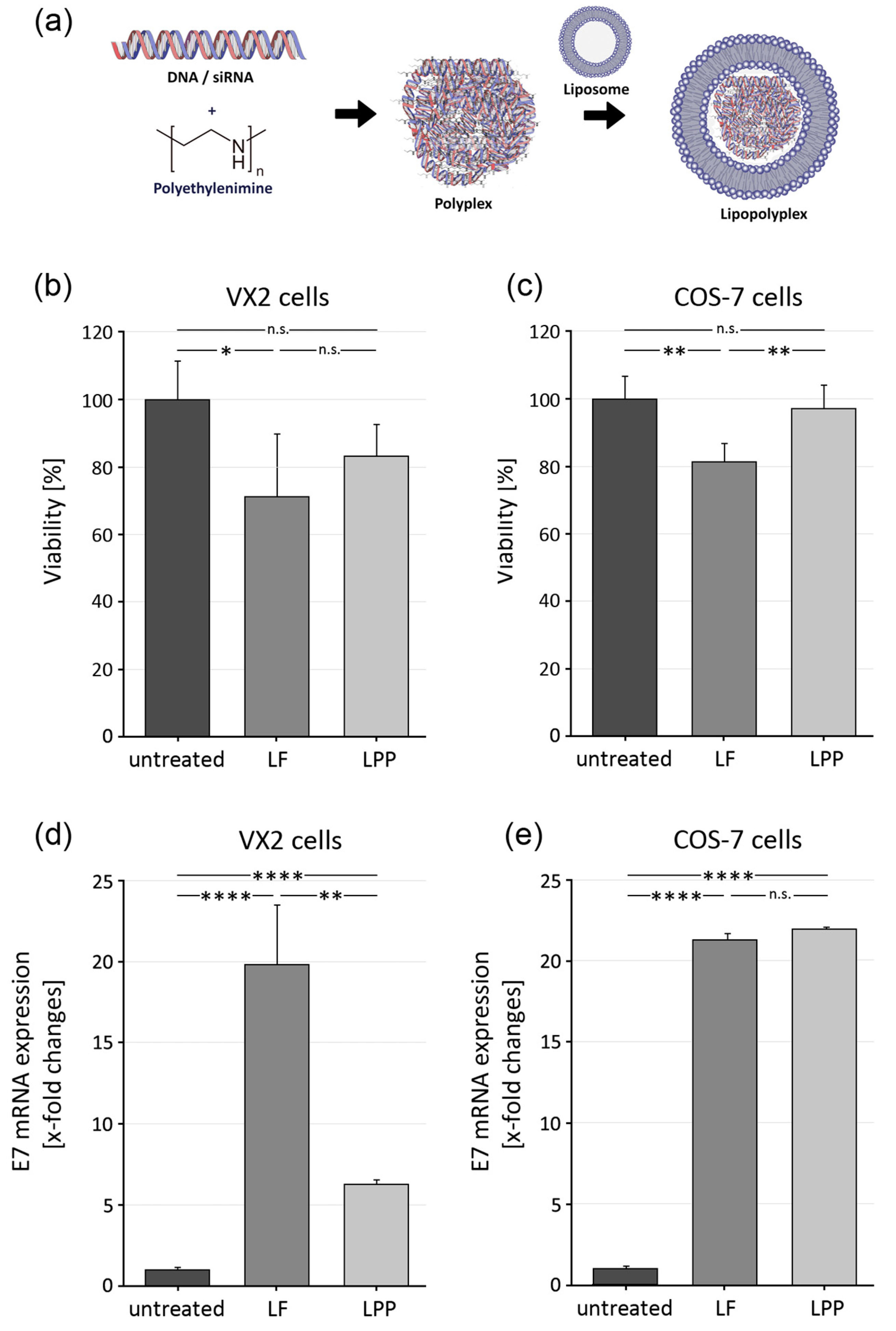
3.2. Generation of Lipopolyplexes and Evaluation of Cellular Toxicity
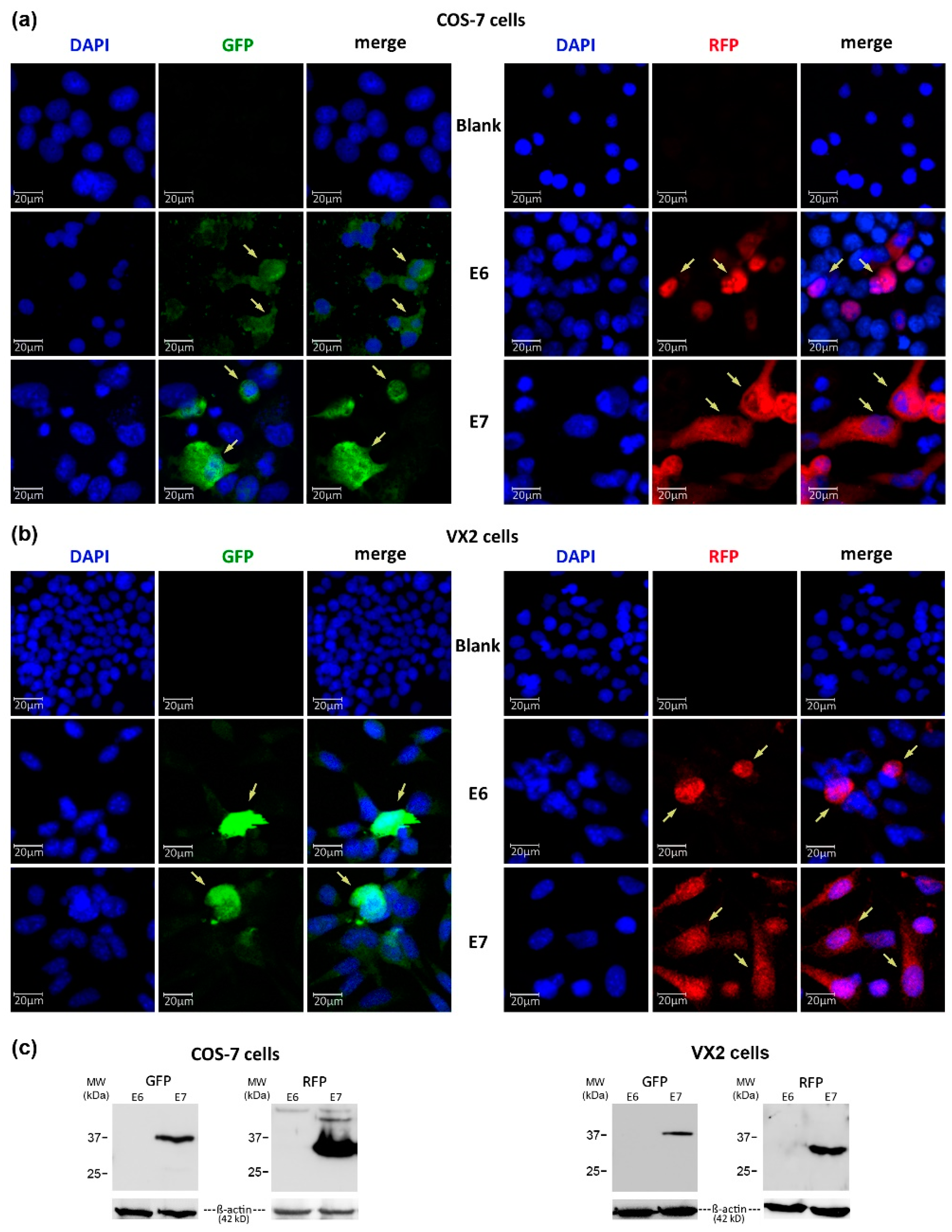
3.3. Transfection Efficiency of Lipopolyplexes in VX2 and COS-7 Cells
3.4. Expression of GFP- and RFP-Tagged CRPV-E6 and -E7 Oncoproteins in VX2 and COS-7 Cells
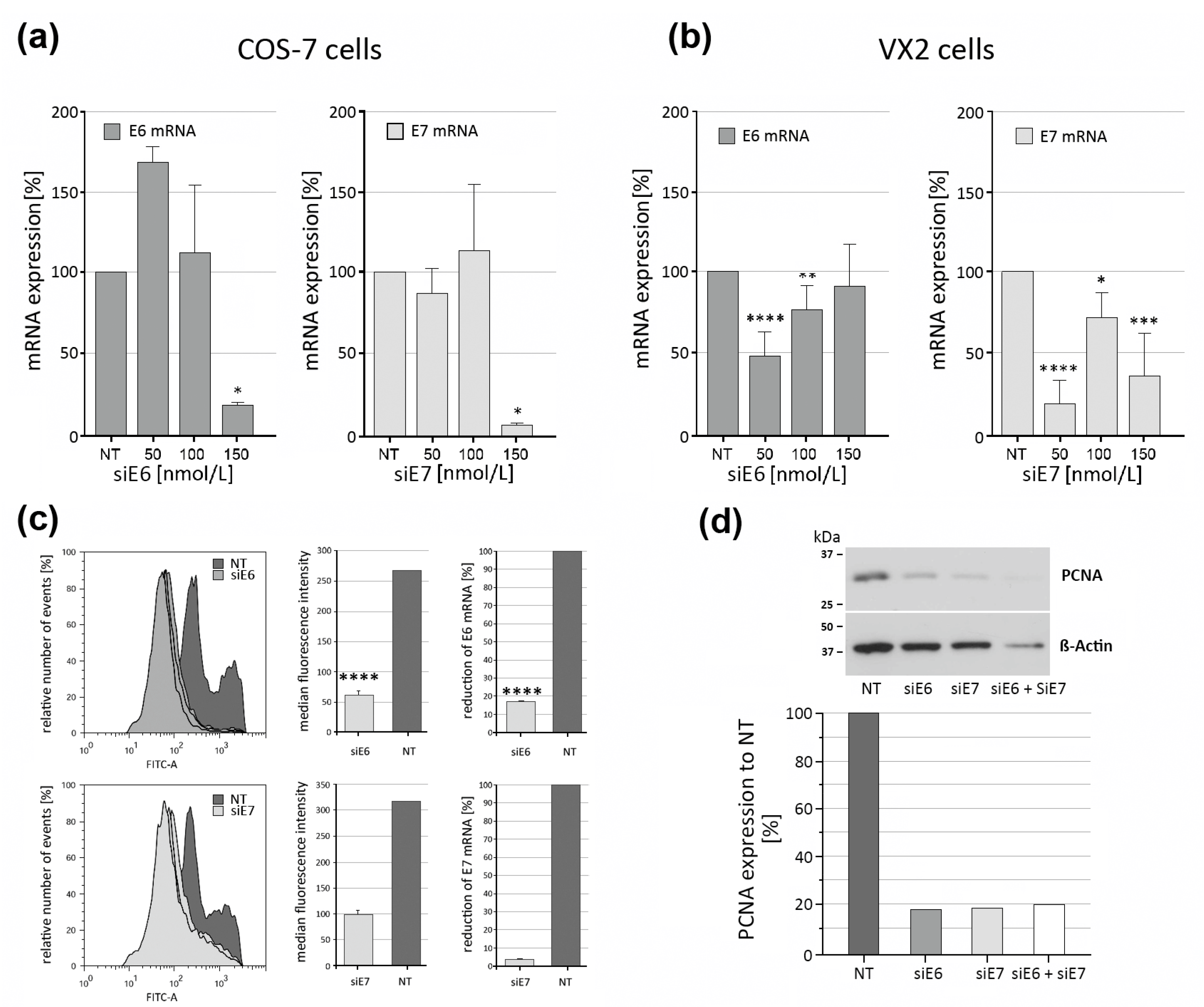
3.5. Effectiveness of CRPV E6 and E7 RNAi Knockdown Using siRNA-Loaded LPPs
3.6. Wound Closure Assay and Real-Time Cellular Analysis (RTCA)
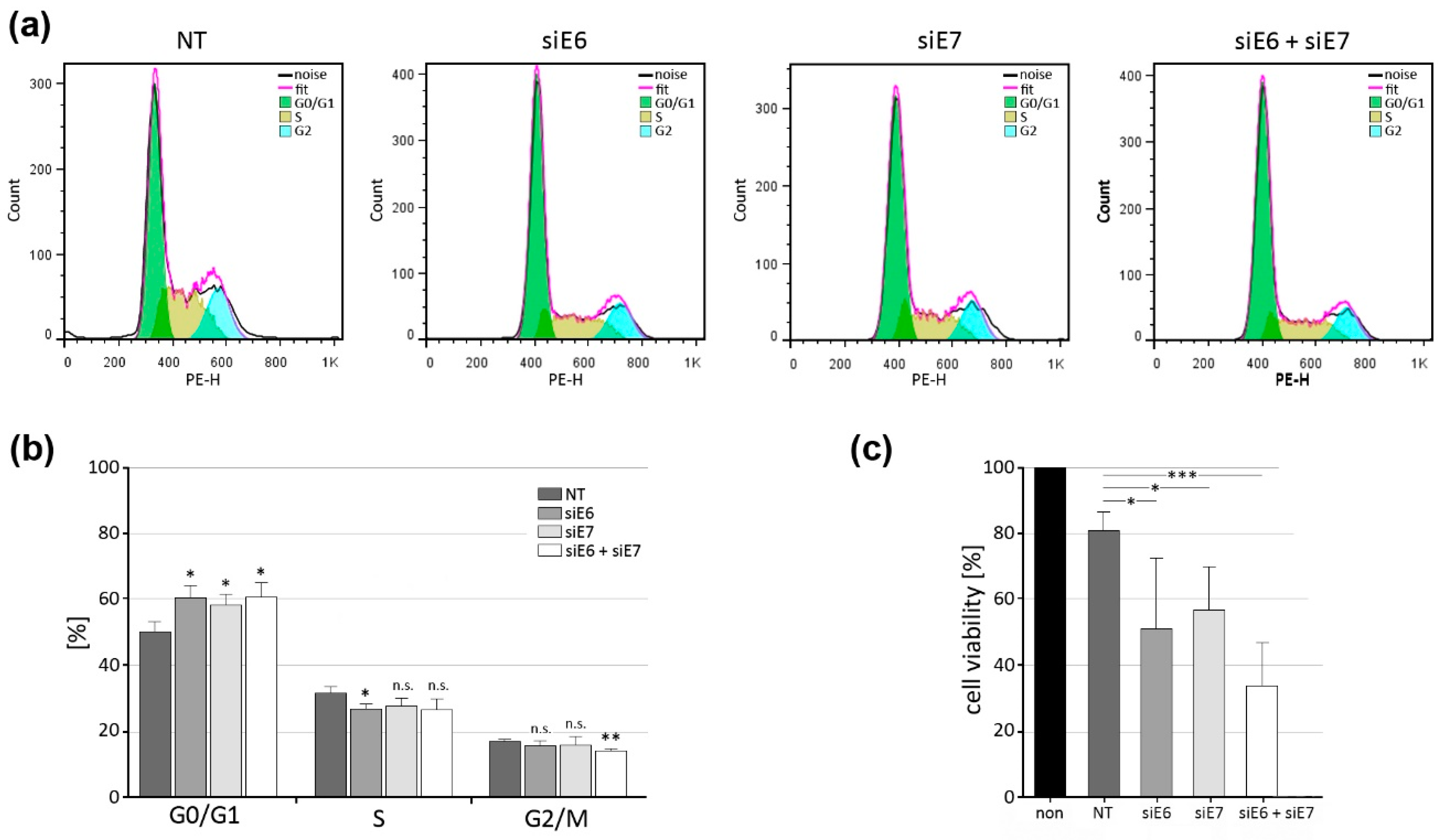
3.7. Indication of G1 Block after CRPV E6 and E7 Knockdown in VX2 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaikhoutdinov, I.; Goldenberg, D. Impact of genetic targets on therapy in head and neck squamous cell carcinoma. Adv. Exp. Med. Biol. 2013, 779, 165–177. [Google Scholar] [CrossRef]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef]
- Goldberg, H.I.; Lockwood, S.A.; Wyatt, S.W.; Crossett, L.S. Trends and differentials in mortality from cancers of the oral cavity and pharynx in the United States, 1973–1987. Cancer 1994, 74, 565–572. [Google Scholar] [CrossRef]
- van Es, R.J.; Franssen, O.; Dullens, H.F.; Bernsen, M.R.; Bosman, F.; Hennink, W.E.; Slootweg, P.J. The VX2 carcinoma in the rabbit auricle as an experimental model for intra-arterial embolization of head and neck squamous cell carcinoma with dextran microspheres. Lab. Anim. 1999, 33, 175–184. [Google Scholar] [CrossRef]
- Mandic, R.; Dunne, A.A.; Eikelkamp, N.; Ramaswamy, A.; Schulz, S.; Teymoortash, A.; Sesterhenn, A.; Moll, R.; Werner, J.A. Expression of MMP-3, MMP-13, TIMP-2 and TIMP-3 in the VX2 carcinoma of the New Zealand white rabbit. Anticancer Res. 2002, 22, 3281–3284. [Google Scholar]
- Amella, C.A.; Lofgren, L.A.; Ronn, A.M.; Nouri, M.; Shikowitz, M.J.; Steinberg, B.M. Latent infection induced with cottontail rabbit papillomavirus. A model for human papillomavirus latency. Am. J. Pathol. 1994, 144, 1167–1171. [Google Scholar]
- Shope, R.E.; Hurst, E.W. Infectious Papillomatosis of Rabbits: With a Note on the Histopathology. J. Exp. Med. 1933, 58, 607–624. [Google Scholar] [CrossRef]
- Shope, R.E. A Transmissible Tumor-Like Condition in Rabbits. J. Exp. Med. 1932, 56, 793–802. [Google Scholar] [CrossRef]
- Kreider, J.W.; Bartlett, G.L. Shope rabbit papilloma--carcinoma complex. A model system of HPV infections. Clin. Dermatol. 1985, 3, 20–26. [Google Scholar] [CrossRef]
- Spiotto, M.T.; Pytynia, M.; Liu, G.F.; Ranck, M.C.; Widau, R. Animal models to study the mutational landscape for oral cavity and oropharyngeal cancers. J. Oral Maxillofac. Res. 2013, 4, e1. [Google Scholar] [CrossRef]
- Hubrecht, R.C.; Carter, E. The 3Rs and Humane Experimental Technique: Implementing Change. Animals 2019, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Osato, T.; Ito, Y. In vitro cultivation and immunofluorescent studies of transplantable carcinomas Vx2 and Vx7. Persistence of a Shope virus-related antigenic substance in the cells of both tumors. J. Exp. Med. 1967, 126, 881–886. [Google Scholar] [CrossRef]
- Georges, E.; Breitburd, F.; Jibard, N.; Orth, G. Two Shope papillomavirus-associated VX2 carcinoma cell lines with different levels of keratinocyte differentiation and transplantability. J. Virol. 1985, 55, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Voelkel, E.F.; Tashjian, A.H., Jr.; Franklin, R.; Wasserman, E.; Levine, L. Hypercalcemia and tumor-prostaglandins: The VX2 carcinoma model in the rabbit. Metabolism 1975, 24, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Easty, D.M.; Easty, G.C. Establishment of an in vitro cell line from the rabbit VX2 carcinoma. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1982, 39, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.F.; Ren, L.R.; Su, G.Y.; Liu, Y.Q.; Gu, B.; Dong, J.H. Establishment and characterization of a rabbit tumor cell line VX2. Zhonghua Bing Li Xue Za Zhi 2005, 34, 661–663. [Google Scholar] [PubMed]
- Virmani, S.; Harris, K.R.; Szolc-Kowalska, B.; Paunesku, T.; Woloschak, G.E.; Lee, F.T.; Lewandowski, R.J.; Sato, K.T.; Ryu, R.K.; Salem, R.; et al. Comparison of two different methods for inoculating VX2 tumors in rabbit livers and hind limbs. J. Vasc. Interv. Radiol. 2008, 19, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Blitzer, G.C.; Smith, M.A.; Harris, S.L.; Kimple, R.J. Review of the clinical and biologic aspects of human papillomavirus-positive squamous cell carcinomas of the head and neck. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 761–770. [Google Scholar] [CrossRef]
- Gluzman, Y. SV40-transformed simian cells support the replication of early SV40 mutants. Cell 1981, 23, 175–182. [Google Scholar] [CrossRef]
- Muench, P.; Probst, S.; Schuetz, J.; Leiprecht, N.; Busch, M.; Wesselborg, S.; Stubenrauch, F.; Iftner, T. Cutaneous papillomavirus E6 proteins must interact with p300 and block p53-mediated apoptosis for cellular immortalization and tumorigenesis. Cancer Res. 2010, 70, 6913–6924. [Google Scholar] [CrossRef]
- Meyers, C.; Harry, J.; Lin, Y.L.; Wettstein, F.O. Identification of three transforming proteins encoded by cottontail rabbit papillomavirus. J. Virol. 1992, 66, 1655–1664. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2019, 10, 3116. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-viral vectors for gene-based therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Ewe, A.; Panchal, O.; Pinnapireddy, S.R.; Bakowsky, U.; Przybylski, S.; Temme, A.; Aigner, A. Liposome-polyethylenimine complexes (DPPC-PEI lipopolyplexes) for therapeutic siRNA delivery in vivo. Nanomedicine 2017, 13, 209–218. [Google Scholar] [CrossRef]
- Hadinoto, K.; Sundaresan, A.; Cheow, W.S. Lipid-polymer hybrid nanoparticles as a new generation therapeutic delivery platform: A review. Eur. J. Pharm. Biopharm. 2013, 85, 427–443. [Google Scholar] [CrossRef]
- Pinnapireddy, S.R.; Duse, L.; Strehlow, B.; Schafer, J.; Bakowsky, U. Composite liposome-PEI/nucleic acid lipopolyplexes for safe and efficient gene delivery and gene knockdown. Colloids Surf. B Biointerfaces 2017, 158, 93–101. [Google Scholar] [CrossRef]
- Tariq, I.; Pinnapireddy, S.R.; Duse, L.; Ali, M.Y.; Ali, S.; Amin, M.U.; Goergen, N.; Jedelska, J.; Schafer, J.; Bakowsky, U. Lipodendriplexes: A promising nanocarrier for enhanced gene delivery with minimal cytotoxicity. Eur. J. Pharm. Biopharm. 2019, 135, 72–82. [Google Scholar] [CrossRef]
- Hamidi, H.; Lilja, J.; Ivaska, J. Using xCELLigence RTCA Instrument to Measure Cell Adhesion. Bio Protoc. 2017, 7, e2646. [Google Scholar] [CrossRef]
- Franke, N.; Bette, M.; Marquardt, A.; Briese, T.; Lipkin, W.I.; Kurz, C.; Ehrenreich, J.; Mack, E.; Baying, B.; Benes, V.; et al. Virome Analysis Reveals No Association of Head and Neck Vascular Anomalies with an Active Viral Infection. In Vivo 2018, 32, 1323–1331. [Google Scholar] [CrossRef]
- Chen, J.; Xue, Y.; Poidinger, M.; Lim, T.; Chew, S.H.; Pang, C.L.; Abastado, J.P.; Thierry, F. Mapping of HPV transcripts in four human cervical lesions using RNAseq suggests quantitative rearrangements during carcinogenic progression. Virology 2014, 462–463, 14–24. [Google Scholar] [CrossRef]
- Gharaati-Far, N.; Tohidkia, M.R.; Dehnad, A.; Omidi, Y. Efficiency and cytotoxicity analysis of cationic lipids-mediated gene transfection into AGS gastric cancer cells. Artif. Cells Nanomed. Biotechnol. 2018, 46, 1001–1008. [Google Scholar] [CrossRef]
- Liu, Y.; Cherry, J.J.; Dineen, J.V.; Androphy, E.J.; Baleja, J.D. Determinants of stability for the E6 protein of papillomavirus type 16. J. Mol. Biol. 2009, 386, 1123–1137. [Google Scholar] [CrossRef]
- Zhu, D.; Ye, M.; Zhang, W. E6/E7 oncoproteins of high risk HPV-16 upregulate MT1-MMP, MMP-2 and MMP-9 and promote the migration of cervical cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 4981–4989. [Google Scholar]
- Ke, N.; Wang, X.; Xu, X.; Abassi, Y.A. The xCELLigence system for real-time and label-free monitoring of cell viability. Methods Mol. Biol. 2011, 740, 33–43. [Google Scholar] [CrossRef]
- Li, S.J.; Ren, G.X.; Jin, W.L.; Guo, W. Establishment and characterization of a rabbit oral squamous cell carcinoma cell line as a model for in vivo studies. Oral Oncol. 2011, 47, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Butz, K.; Ristriani, T.; Hengstermann, A.; Denk, C.; Scheffner, M.; Hoppe-Seyler, F. siRNA targeting of the viral E6 oncogene efficiently kills human papillomavirus-positive cancer cells. Oncogene 2003, 22, 5938–5945. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cladel, N.M.; Balogh, K.; Budgeon, L.; Christensen, N.D. Impact of genetic changes to the CRPV genome and their application to the study of pathogenesis in vivo. Virology 2007, 358, 384–390. [Google Scholar] [CrossRef]
- Tagalakis, A.D.; Maeshima, R.; Yu-Wai-Man, C.; Meng, J.; Syed, F.; Wu, L.P.; Aldossary, A.M.; McCarthy, D.; Moghimi, S.M.; Hart, S.L. Peptide and nucleic acid-directed self-assembly of cationic nanovehicles through giant unilamellar vesicle modification: Targetable nanocomplexes for in vivo nucleic acid delivery. Acta Biomater. 2017, 51, 351–362. [Google Scholar] [CrossRef]
- Linder, B.; Weirauch, U.; Ewe, A.; Uhmann, A.; Seifert, V.; Mittelbronn, M.; Harter, P.N.; Aigner, A.; Kogel, D. Therapeutic Targeting of Stat3 Using Lipopolyplex Nanoparticle-Formulated siRNA in a Syngeneic Orthotopic Mouse Glioma Model. Cancers 2019, 11, 333. [Google Scholar] [CrossRef]
- Kurosaki, T.; Kawakami, S.; Higuchi, Y.; Suzuki, R.; Maruyama, K.; Sasaki, H.; Yamashita, F.; Hashida, M. Development of anionic bubble lipopolyplexes for efficient and safe gene transfection with ultrasound exposure in mice. J. Control. Release 2014, 176, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Amin, M.U.; Tariq, I.; Sohail, M.F.; Ali, M.Y.; Preis, E.; Ambreen, G.; Pinnapireddy, S.R.; Jedelska, J.; Schafer, J.; et al. Lipoparticles for Synergistic Chemo-Photodynamic Therapy to Ovarian Carcinoma Cells: In vitro and in vivo Assessments. Int. J. Nanomed. 2021, 16, 951–976. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, U.; Bette, M.; Ambreen, G.; Pinnapireddy, S.R.; Tariq, I.; Marquardt, A.; Stuck, B.A.; Bakowsky, U.; Mandic, R. RNAi-Mediated Knockdown of Cottontail Rabbit Papillomavirus Oncogenes Using Low-Toxicity Lipopolyplexes as a Paradigm to Treat Papillomavirus-Associated Cancers. Pharmaceutics 2023, 15, 2379. https://doi.org/10.3390/pharmaceutics15102379
Ali U, Bette M, Ambreen G, Pinnapireddy SR, Tariq I, Marquardt A, Stuck BA, Bakowsky U, Mandic R. RNAi-Mediated Knockdown of Cottontail Rabbit Papillomavirus Oncogenes Using Low-Toxicity Lipopolyplexes as a Paradigm to Treat Papillomavirus-Associated Cancers. Pharmaceutics. 2023; 15(10):2379. https://doi.org/10.3390/pharmaceutics15102379
Chicago/Turabian StyleAli, Uzma, Michael Bette, Ghazala Ambreen, Shashank R. Pinnapireddy, Imran Tariq, André Marquardt, Boris A. Stuck, Udo Bakowsky, and Robert Mandic. 2023. "RNAi-Mediated Knockdown of Cottontail Rabbit Papillomavirus Oncogenes Using Low-Toxicity Lipopolyplexes as a Paradigm to Treat Papillomavirus-Associated Cancers" Pharmaceutics 15, no. 10: 2379. https://doi.org/10.3390/pharmaceutics15102379