

Granulocyte Colony-Stimulating Factor Improves Endothelial Progenitor Cell-Mediated Neovascularization in Mice with Chronic Kidney Disease
 and
and 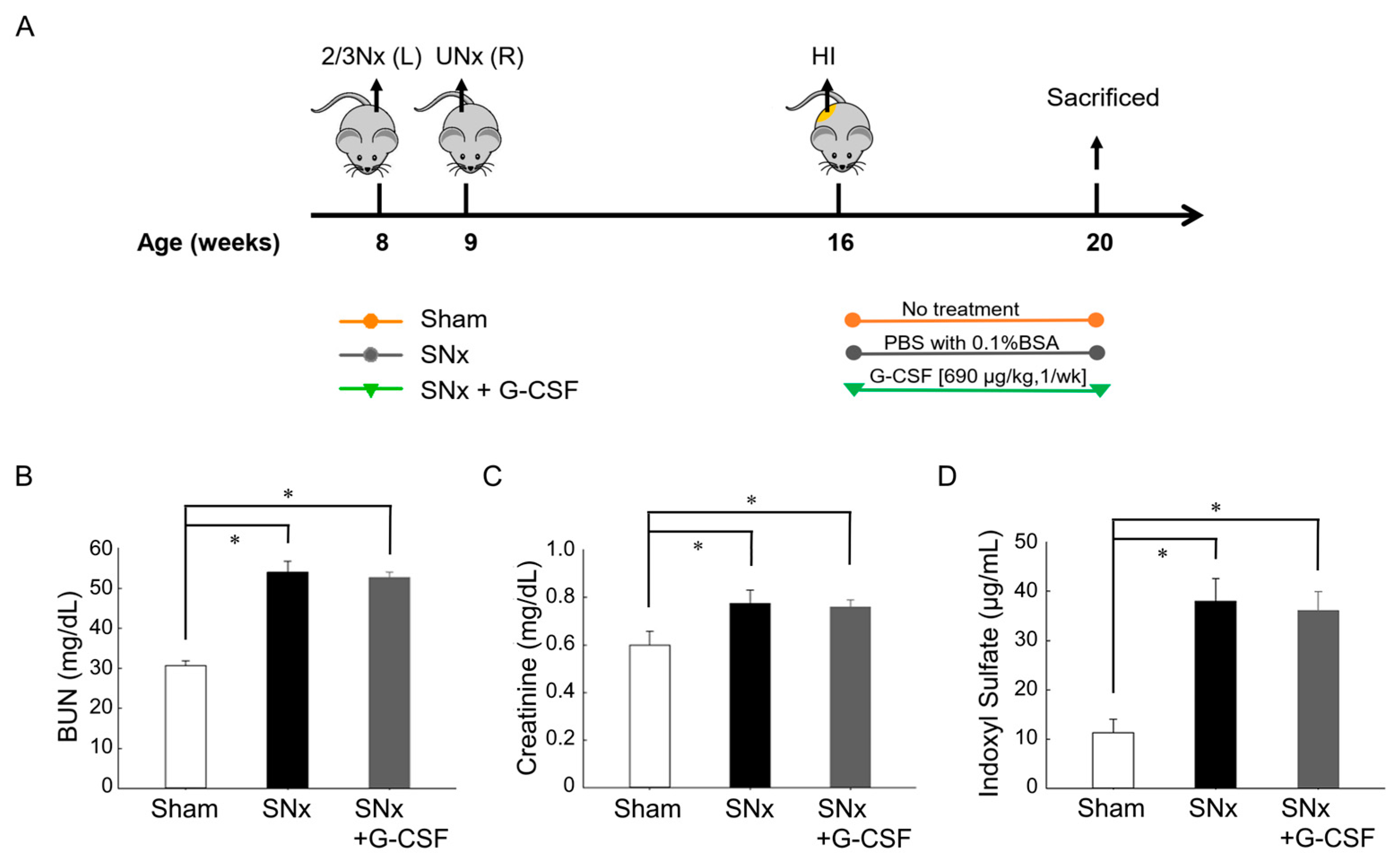{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Animals
2.3. Subtotal Nephrectomy
2.4. Hindlimb Ischemia Surgery
2.5. Measurement of Renal Function and Indoxyl Sulfate
2.6. Laser Doppler Imaging
2.7. Flow Cytometry
2.8. ELISA
2.9. Histological Analysis
2.10. Western Blotting
2.11. Human EPC Isolation and Cultivation
2.12. MTT Assay
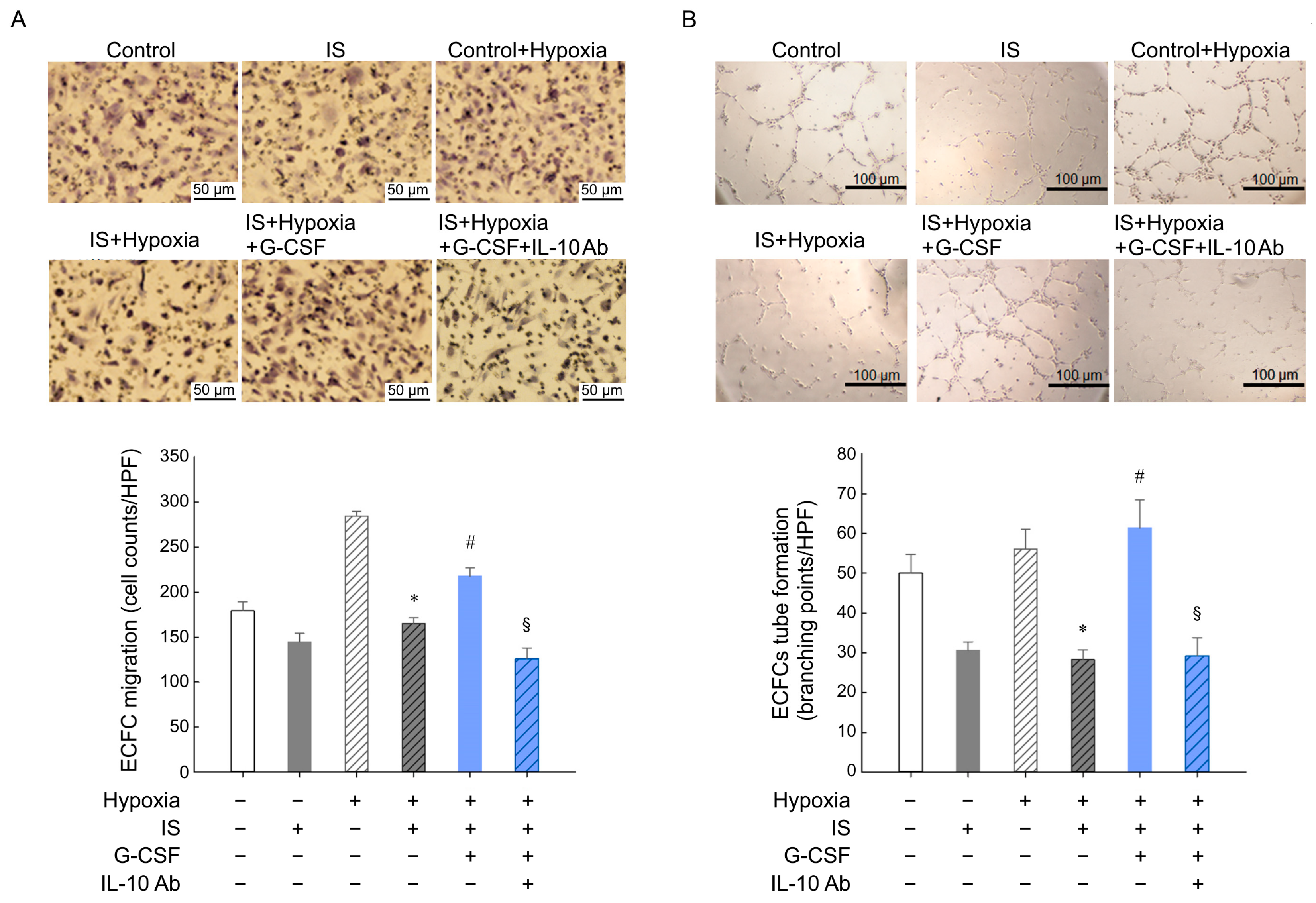
2.13. ECFC Migration and Tube Formation Assays
2.14. Statistical Analysis
3. Results
3.1. Renal Function Decline and Increase in Plasma IS Levels after Subtotal Nephrectomy
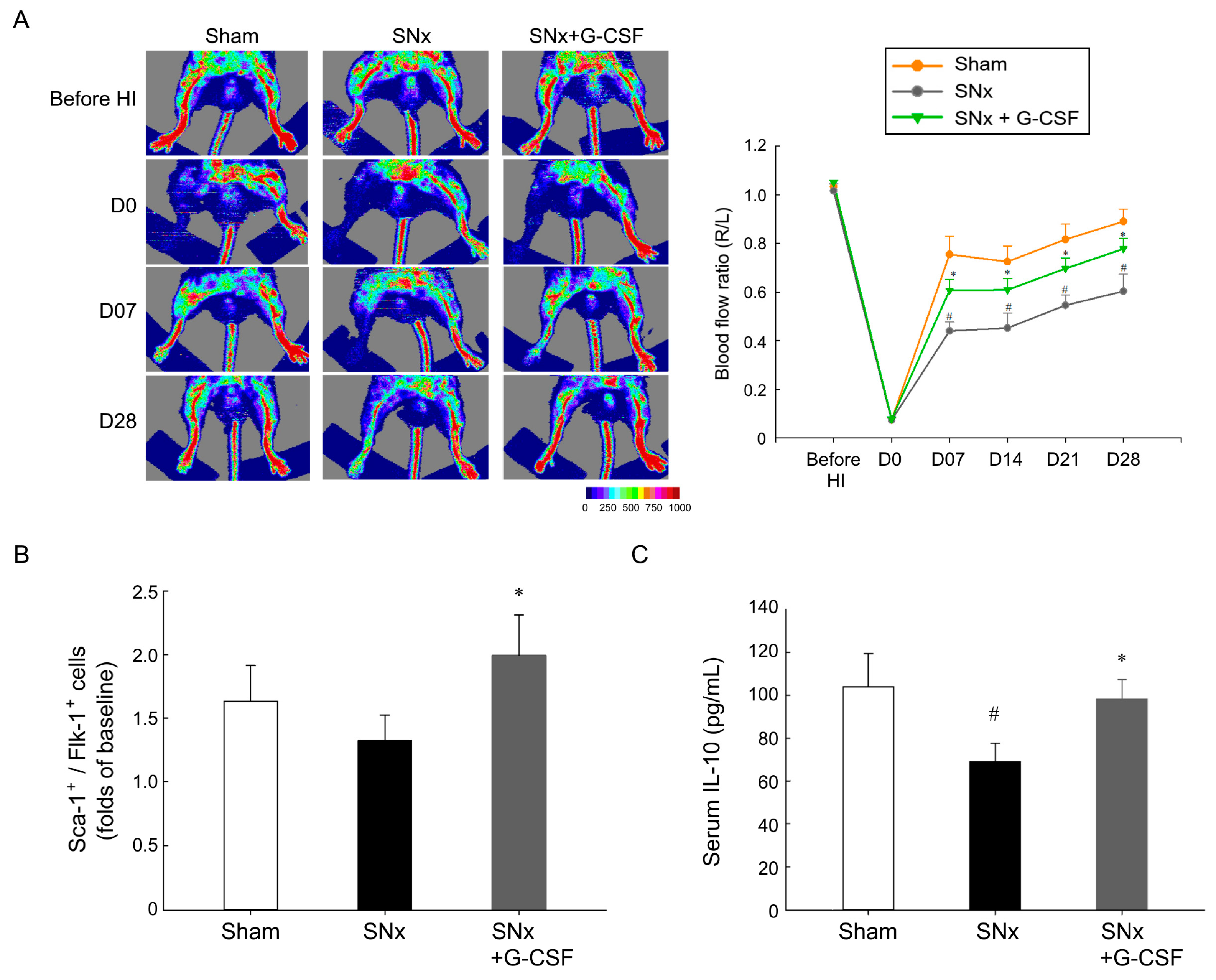
3.2. G-CSF Reinstated Blood Flow Reperfusion in Ischemic Hindlimbs, Mobilized EPCs, and Elevated Plasma IL-10 Levels in SNx Mice
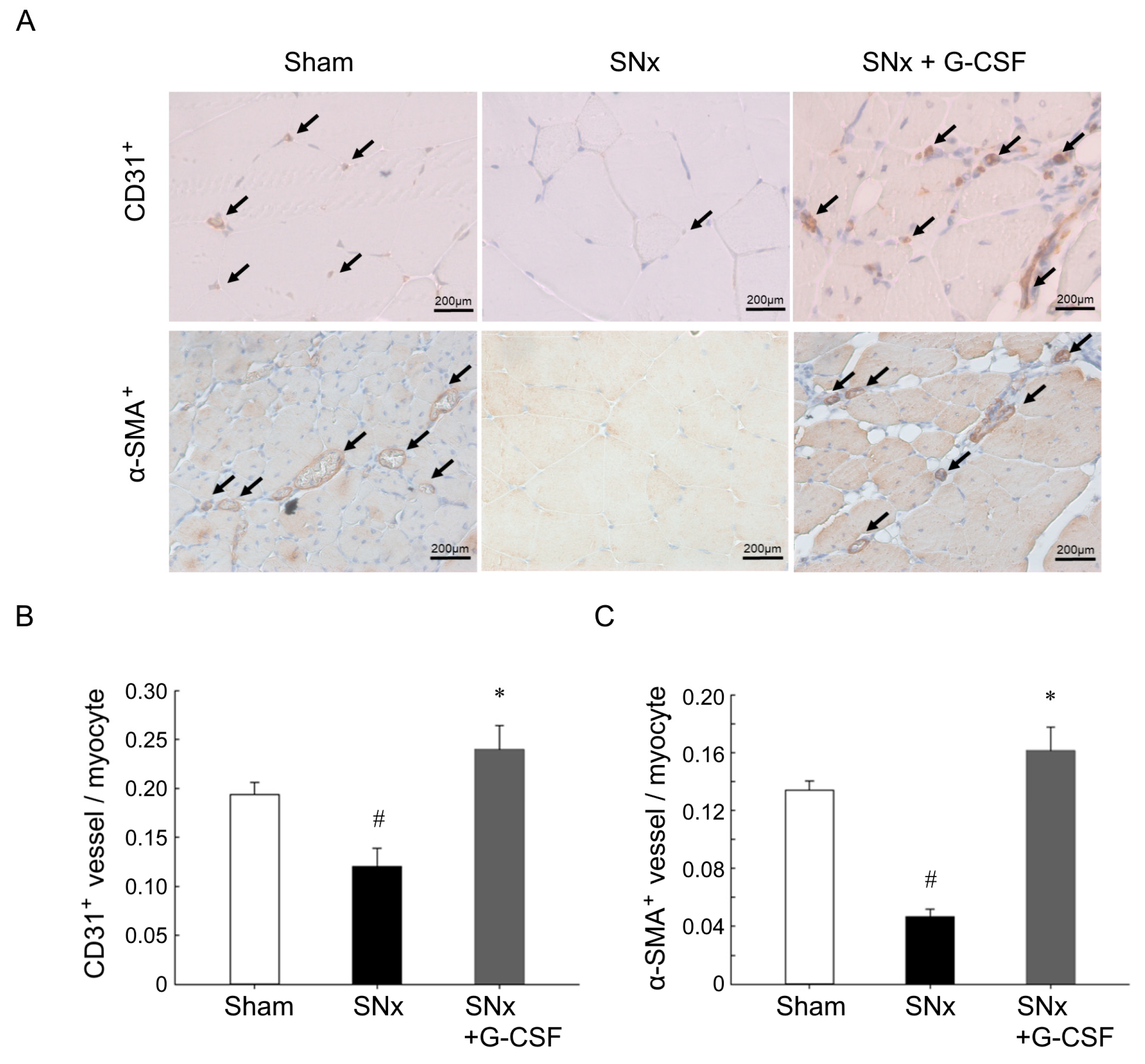
3.3. G-CSF Restored Impaired Neovascularization Induced by Ischemia in SNx Mice
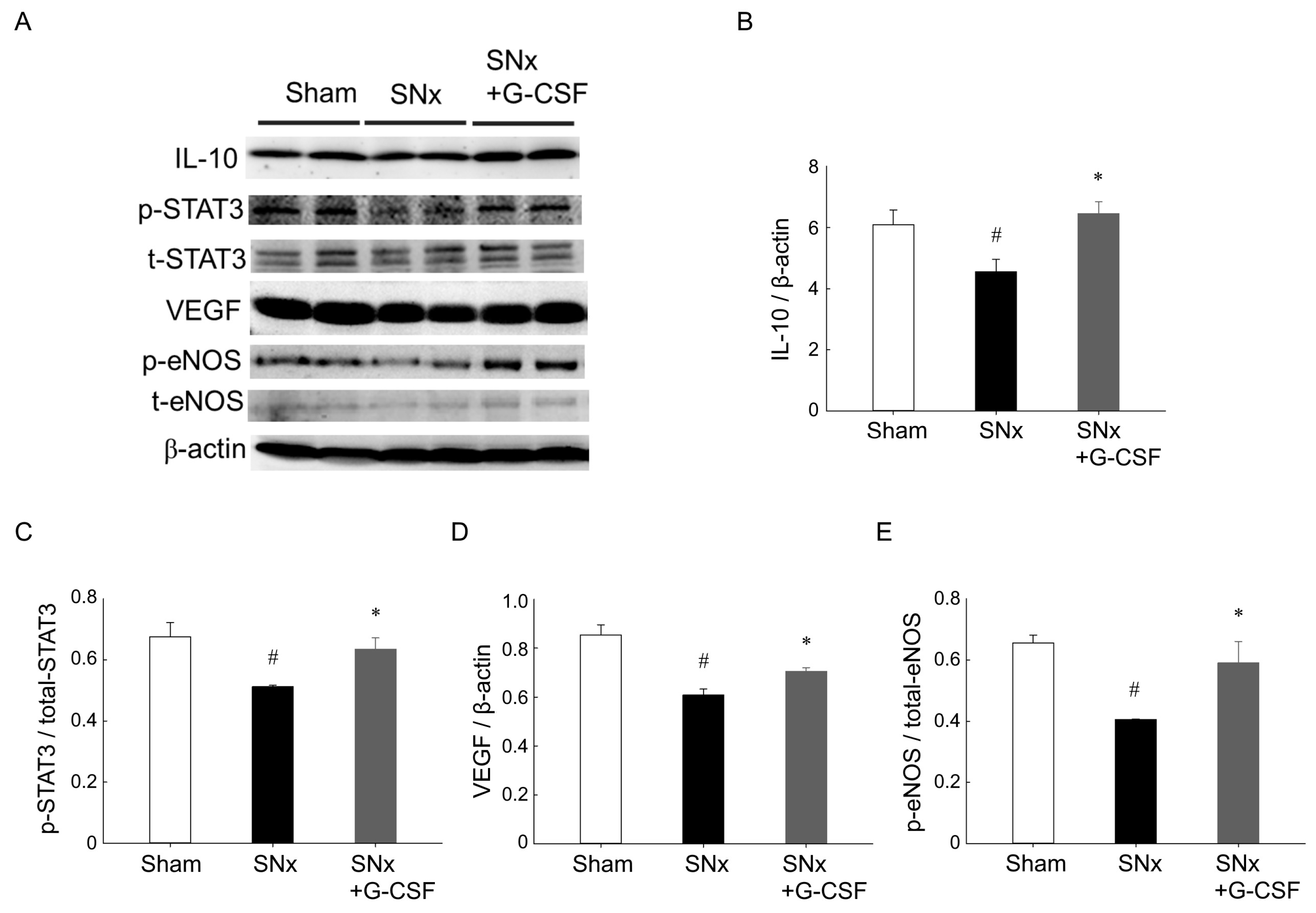
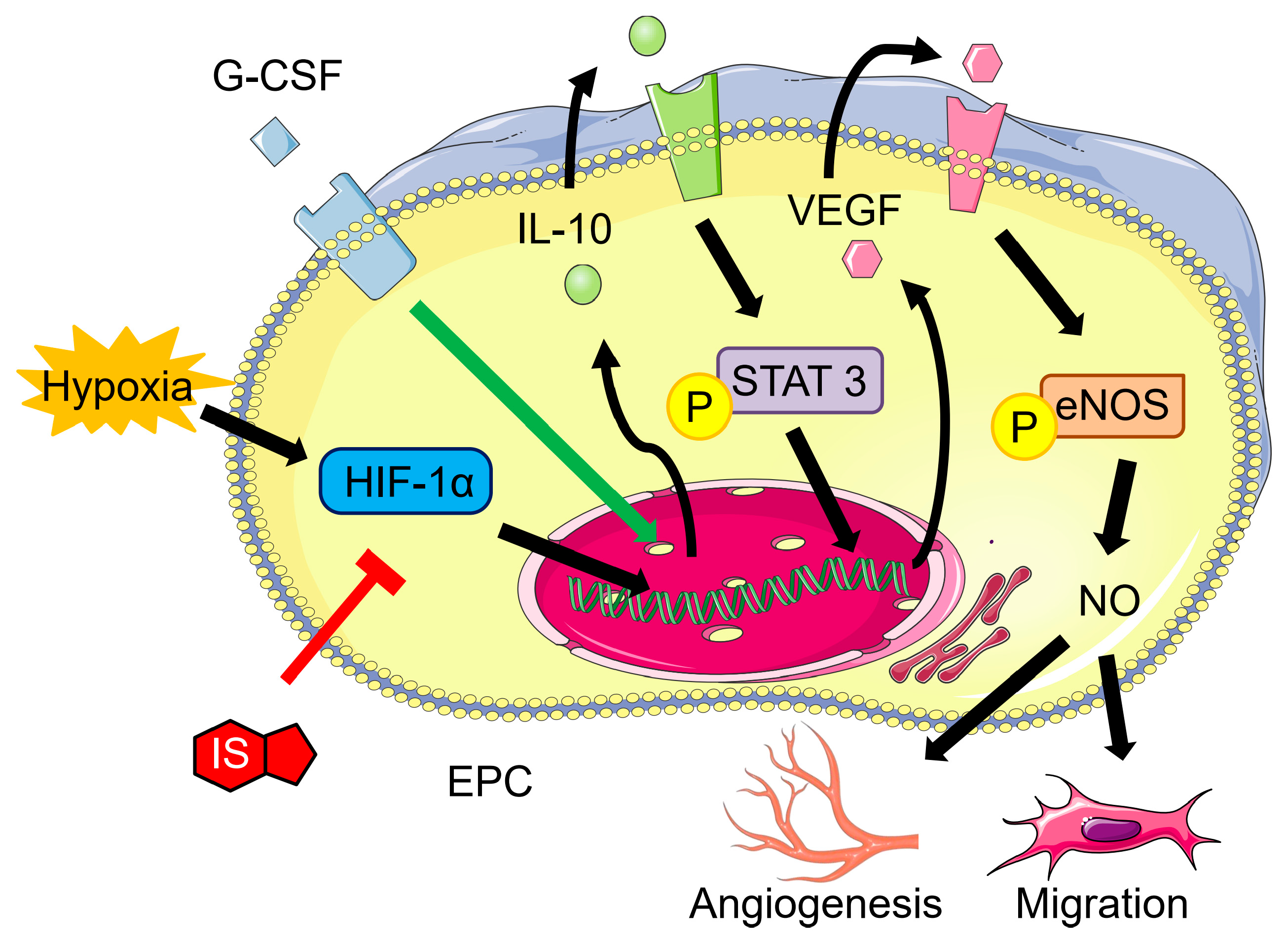
3.4. G-CSF Modulates Protein Expression in Ischemic Hindlimbs from SNx Mice
3.5. G-CSF Modulates Protein Expression in Ischemic Hindlimbs from SNx Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wattanakit, K.; Folsom, A.R.; Selvin, E.; Coresh, J.; Hirsch, A.T.; Weatherley, B.D. Kidney function and risk of peripheral arterial disease: Results from the atherosclerosis risk in communities (ARIC) study. J. Am. Soc. Nephrol. JASN 2007, 18, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Lash, J.P.; Go, A.S.; Appel, L.J.; He, J.; Ojo, A.; Rahman, M.; Townsend, R.R.; Xie, D.; Cifelli, D.; Cohan, J.; et al. Chronic renal insufficiency cohort (cric) study: Baseline characteristics and associations with kidney function. Clin. J. Am. Soc. Nephrol. CJASN 2009, 4, 1302–1311. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.K.; Sarnak, M.J.; Yan, G.; Dwyer, J.T.; Heyka, R.J.; Rocco, M.V.; Teehan, B.P.; Levey, A.S. Atherosclerotic cardiovascular disease risks in chronic hemodialysis patients. Kidney Int. 2000, 58, 353–362. [Google Scholar] [CrossRef]
- Selvin, E.; Erlinger, T.P. Prevalence of and risk factors for peripheral arterial disease in the united states: Results from the national health and nutrition examination survey, 1999–2000. Circulation 2004, 110, 738–743. [Google Scholar] [CrossRef]
- Eggers, P.W.; Gohdes, D.; Pugh, J. Nontraumatic lower extremity amputations in the medicare end-stage renal disease population. Kidney Int. 1999, 56, 1524–1533. [Google Scholar] [CrossRef]
- Fishbane, S.; Youn, S.; Flaster, E.; Adam, G.; Maesaka, J.K. Ankle-arm blood pressure index as a predictor of mortality in hemodialysis patients. Am. J. Kidney Dis. Off. J. Natl. Kidney Found. 1996, 27, 668–672. [Google Scholar] [CrossRef]
- Duranton, F.; Cohen, G.; De Smet, R.; Rodriguez, M.; Jankowski, J.; Vanholder, R.; Argiles, A. Normal and pathologic concentrations of uremic toxins. J. Am. Soc. Nephrol. JASN 2012, 23, 1258–1270. [Google Scholar] [CrossRef]
- Kuo, K.L.; Zhao, J.F.; Huang, P.H.; Guo, B.C.; Tarng, D.C.; Lee, T.S. Indoxyl sulfate impairs valsartan-induced neovascularization. Redox Biol. 2020, 30, 101433. [Google Scholar] [CrossRef]
- Hung, S.C.; Kuo, K.L.; Huang, H.L.; Lin, C.C.; Tsai, T.H.; Wang, C.H.; Chen, J.W.; Lin, S.J.; Huang, P.H.; Tarng, D.C. Indoxyl sulfate suppresses endothelial progenitor cell-mediated neovascularization. Kidney Int. 2016, 89, 574–585. [Google Scholar] [CrossRef]
- Adusumilli, A.; Krothapalli, S.R. The clinical applications of hematopoietic growth factor-GCSF. Asian J. Biomed. Pharm. Sci. 2012, 2, 1–10. [Google Scholar]
- Tsai, R.K.; Lin, K.L.; Huang, C.T.; Wen, Y.T. Transcriptomic analysis reveals that granulocyte colony-stimulating factor trigger a novel signaling pathway (taf9-p53-triap1-casp3) to protect retinal ganglion cells after ischemic optic neuropathy. Int. J. Mol. Sci. 2022, 23, 8359. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, J.F.; Souza, B.S.; Lins, T.F.; Garcia, L.M.; Kaneto, C.M.; Sampaio, G.P.; de Alcântara, A.C.; Meira, C.S.; Macambira, S.G.; Ribeiro-dos-Santos, R.; et al. Administration of granulocyte colony-stimulating factor induces immunomodulation, recruitment of t regulatory cells, reduction of myocarditis and decrease of parasite load in a mouse model of chronic chagas disease cardiomyopathy. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 4691–4702. [Google Scholar] [CrossRef] [PubMed]
- Perobelli, S.M.; Mercadante, A.C.; Galvani, R.G.; Gonçalves-Silva, T.; Alves, A.P.; Pereira-Neves, A.; Benchimol, M.; Nóbrega, A.; Bonomo, A. G-csf-induced suppressor il-10+ neutrophils promote regulatory t cells that inhibit graft-versus-host disease in a long-lasting and specific way. J. Immunol. 2016, 197, 3725–3734. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; Lin, L.Y.; Chen, J.W. Inhibition of macrophage inflammatory protein-1β improves endothelial progenitor cell function and ischemia-induced angiogenesis in diabetes. Angiogenesis 2019, 22, 53–65. [Google Scholar] [CrossRef]
- Huang, P.H.; Lin, C.P.; Wang, C.H.; Chiang, C.H.; Tsai, H.Y.; Chen, J.S.; Lin, F.Y.; Leu, H.B.; Wu, T.C.; Chen, J.W.; et al. Niacin improves ischemia-induced neovascularization in diabetic mice by enhancement of endothelial progenitor cell functions independent of changes in plasma lipids. Angiogenesis 2012, 15, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.H.; Chen, Y.H.; Wang, C.H.; Chen, J.S.; Tsai, H.Y.; Lin, F.Y.; Lo, W.Y.; Wu, T.C.; Sata, M.; Chen, J.W.; et al. Matrix metalloproteinase-9 is essential for ischemia-induced neovascularization by modulating bone marrow-derived endothelial progenitor cells. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1179–1184. [Google Scholar] [CrossRef]
- Béland, S.; Désy, O.; Bouchard-Boivin, F.; Gama, A.; De Serres, S.A. Endothelial colony forming cells generated from cryopreserved peripheral blood mononuclear cells. Hum. Immunol. 2021, 82, 309–314. [Google Scholar] [CrossRef]
- Chen, J.Z.; Zhu, J.H.; Wang, X.X.; Zhu, J.H.; Xie, X.D.; Sun, J.; Shang, Y.P.; Guo, X.G.; Dai, H.M.; Hu, S.J. Effects of homocysteine on number and activity of endothelial progenitor cells from peripheral blood. J. Mol. Cell. Cardiol. 2004, 36, 233–239. [Google Scholar] [CrossRef]
- Bevan, G.H.; White Solaru, K.T. Evidence-based medical management of peripheral artery disease. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 541–553. [Google Scholar] [CrossRef]
- Tran, B. Assessment and management of peripheral arterial disease: What every cardiologist should know. Heart 2021, 107, 1835–1843. [Google Scholar] [CrossRef]
- Serra, R.; Bracale, U.M.; Ielapi, N.; Del Guercio, L.; Di Taranto, M.D.; Sodo, M.; Michael, A.; Faga, T.; Bevacqua, E.; Jiritano, F.; et al. The impact of chronic kidney disease on peripheral artery disease and peripheral revascularization. Int. J. Gen. Med. 2021, 14, 3749–3759. [Google Scholar] [CrossRef] [PubMed]
- Shokane, L.L.; Bezuidenhout, S.; Lundie, M. Use of granulocyte colony-stimulating factor in patients with chemotherapy-induced neutropaenia. Health Gesondheid 2023, 28, 2221. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.H.; Tsai, H.Y.; Wang, C.H.; Chen, Y.H.; Chen, J.S.; Lin, F.Y.; Lin, C.P.; Wu, T.C.; Sata, M.; Chen, J.W.; et al. Moderate intake of red wine improves ischemia-induced neovascularization in diabetic mice—Roles of endothelial progenitor cells and nitric oxide. Atherosclerosis 2010, 212, 426–435. [Google Scholar] [CrossRef]
- Del Papa, N.; Pignataro, F. The role of endothelial progenitors in the repair of vascular damage in systemic sclerosis. Front. Immunol. 2018, 9, 1383. [Google Scholar] [CrossRef]
- Asahara, T.; Takahashi, T.; Masuda, H.; Kalka, C.; Chen, D.; Iwaguro, H.; Inai, Y.; Silver, M.; Isner, J.M. Vegf contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells. EMBO J. 1999, 18, 3964–3972. [Google Scholar] [CrossRef]
- Askari, A.T.; Unzek, S.; Popovic, Z.B.; Goldman, C.K.; Forudi, F.; Kiedrowski, M.; Rovner, A.; Ellis, S.G.; Thomas, J.D.; DiCorleto, P.E.; et al. Effect of stromal-cell-derived factor 1 on stem-cell homing and tissue regeneration in ischaemic cardiomyopathy. Lancet 2003, 362, 697–703. [Google Scholar] [CrossRef]
- Ceradini, D.J.; Kulkarni, A.R.; Callaghan, M.J.; Tepper, O.M.; Bastidas, N.; Kleinman, M.E.; Capla, J.M.; Galiano, R.D.; Levine, J.P.; Gurtner, G.C. Progenitor cell trafficking is regulated by hypoxic gradients through hif-1 induction of sdf-1. Nat. Med. 2004, 10, 858–864. [Google Scholar] [CrossRef]
- Aicher, A.; Zeiher, A.M.; Dimmeler, S. Mobilizing endothelial progenitor cells. Hypertension 2005, 45, 321–325. [Google Scholar] [CrossRef]
- Kalka, C.; Masuda, H.; Takahashi, T.; Gordon, R.; Tepper, O.; Gravereaux, E.; Pieczek, A.; Iwaguro, H.; Hayashi, S.I.; Isner, J.M.; et al. Vascular endothelial growth factor(165) gene transfer augments circulating endothelial progenitor cells in human subjects. Circ. Res. 2000, 86, 1198–1202. [Google Scholar] [CrossRef]
- Moore, M.A.; Hattori, K.; Heissig, B.; Shieh, J.H.; Dias, S.; Crystal, R.G.; Rafii, S. Mobilization of endothelial and hematopoietic stem and progenitor cells by adenovector-mediated elevation of serum levels of SDF-1, VEGF, and angiopoietin-1. Ann. N. Y. Acad. Sci. 2001, 938, 36–45, discussion 45–37. [Google Scholar] [CrossRef]
- Hattori, K.; Dias, S.; Heissig, B.; Hackett, N.R.; Lyden, D.; Tateno, M.; Hicklin, D.J.; Zhu, Z.; Witte, L.; Crystal, R.G.; et al. Vascular endothelial growth factor and angiopoietin-1 stimulate postnatal hematopoiesis by recruitment of vasculogenic and hematopoietic stem cells. J. Exp. Med. 2001, 193, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Yancopoulos, G.D.; Davis, S.; Gale, N.W.; Rudge, J.S.; Wiegand, S.J.; Holash, J. Vascular-specific growth factors and blood vessel formation. Nature 2000, 407, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Omura, T.; Yoshiyama, M.; Matsumoto, R.; Enomoto, S.; Kusuyama, T.; Nishiya, D.; Akioka, K.; Iwao, H.; Takeuchi, K.; et al. Granulocyte-colony stimulating factor augments neovascularization induced by bone marrow transplantation in rat hindlimb ischemia. J. Pharmacol. Sci. 2005, 99, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.; Bradley, L.; Smith, A.; Foxwell, B. Signal transducer and activator of transcription 3 is the dominant mediator of the anti-inflammatory effects of il-10 in human macrophages. J. Immunol. 2004, 172, 567–576. [Google Scholar] [CrossRef]
- Hilfiker-Kleiner, D.; Limbourg, A.; Drexler, H. Stat3-mediated activation of myocardial capillary growth. Trends Cardiovasc. Med. 2005, 15, 152–157. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, S.-Y.; Lee, Y.-C.; Tseng, C.-W.; Huang, P.-H.; Kuo, K.-L.; Tarng, D.-C. Granulocyte Colony-Stimulating Factor Improves Endothelial Progenitor Cell-Mediated Neovascularization in Mice with Chronic Kidney Disease. Pharmaceutics 2023, 15, 2380. https://doi.org/10.3390/pharmaceutics15102380
Tang S-Y, Lee Y-C, Tseng C-W, Huang P-H, Kuo K-L, Tarng D-C. Granulocyte Colony-Stimulating Factor Improves Endothelial Progenitor Cell-Mediated Neovascularization in Mice with Chronic Kidney Disease. Pharmaceutics. 2023; 15(10):2380. https://doi.org/10.3390/pharmaceutics15102380
Chicago/Turabian StyleTang, Shao-Yu, Yi-Chin Lee, Chien-Wei Tseng, Po-Hsun Huang, Ko-Lin Kuo, and Der-Cherng Tarng. 2023. "Granulocyte Colony-Stimulating Factor Improves Endothelial Progenitor Cell-Mediated Neovascularization in Mice with Chronic Kidney Disease" Pharmaceutics 15, no. 10: 2380. https://doi.org/10.3390/pharmaceutics15102380