
Recent Advances in Genomics-Based Approaches for the Development of Intracellular Bacterial Pathogen Vaccines
 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Traditional Vaccine Development

{kind=link}
{kind=link}
{kind=link}
| Bacteria | Illness | Intracellular Lifestyles | Bacterial Factors | Host Cells Localization | Incubation Period | Reference |
|---|---|---|---|---|---|---|
| L. pneumophila Non-spore forming, Gram-negative | Legionnaire’s disease | Intravacuolar pH 6 | Type IV secretion system | Macrophages | 2–10 days | [10] |
| M. tuberculosis Non-spore forming, Acid Fast | Tuberculosis | Intravacuolar pH 6.4 | Type VII secretion system | Macrophage, Cytosol, Phagosome | 4–6 weeks | [11] |
| S. typhi Rod-shaped, Gram-negative | Typhoid fever | Intravacuolar 5 | Type III secretion system | Macrophages | 10–14 days | [12] |
| Brucella spp. Gram-negative coccobacilli | Brucellosis, High fever | Intravacuolar 4 | Type IV secretion system | Neutrophils, Vacuole | 2–4 weeks | [13] |
| Listeria monocytogenes (L. monocytogenes) Gram-positive Bacillus | Listeriosis | Intracytosolic | In1A, In1B | Epithelial cells, Cytosol | 1–2 weeks | [14] |
| Rickettsia rickettsii (R. rickettsii) Gram-indeterminant coccobacilli | Rocky Mountain spotted fever | Intravacuolar | Type IV secretion system | Monocytes/Macrophages | 2–5 days | [15] |
| S. flexneri Non-spore forming, Gram-negative | Enteric disease | Intracytosolic | Type III secretion system | M Cells and Macrophages | 12–96 h | [16] |
| M. leprae Acid Fast Bacillus | Leprosy | Intravacuolar | Type VII secretion system | Schwann cells (SCs) | 4–5 years | [17] |
| S. typhimurium Non-spore forming, Gram-negative | Indigestion, food poisoning | Intravacuolar pH 5 | Type III secretion system | Macrophage, Vacuole | 12–72 h | [18] |
| Chlamydia spp. Gram-indeterminant | Genital and ocular infections | Intravacuolar pH ¼ 7.25 | Type III secretion system | Genital epithelium and conjunctiva, Vacuole, | 1–2 weeks | [19] |
| S. aureus Gram-positive | Dermal infection, osteomyelitis, mastitis | Intravascular | Type IV secretion system | Epithelial Cells, Osteoblast, Endosome, Cytosol | 16–18 h | [20] |
| S. enterica Gram-negative | Paratyphoid and typhoid | Intravacuolar | Type III secretion system | Macrophage, Modified phagosome, Vacuole | 12–72 h | [21] |
| Vaccine Type | Vaccine (Target Bacteria) | Vaccine Status | Reference |
|---|---|---|---|
| WCA | Q-Vax (C. burnetiid) (Dukoral, Shanchol (V. cholerae) R. rickettsia R. prowazekii O. tsutsugamushi | Licensed Licensed Experimental Experimental Experimental | [23] [24] [25] [26] [25] |
| LAVs | BCG (M. tuberculosis) S. enterica spp. BioThrax (B. anthracis) LVS (F. tularensis) Vaxchora (V. cholerae) O. tsutsugamushi R. prowazekii | Licensed Licensed Licensed Licensed Licensed Experimental Experimental | [27] [28] [29] [7] [30] [31] [32] |
| Live recombinant bacteria | M. tuberculosis R. rickettsii C. burnetii S. aureus | Experimental Experimental Experimental Experimental | [33] [34] [35] [36] |
| subunit vaccines | dTAP combined vaccine (C. tetani) dTAP combined vaccine (B. pertussis) Trumenba (N. meningitidis) rPA102 (B. anthracis) S. aureus Trumenba (N. meningitidis) | Licensed Licensed Licensed clinical trial Experimental Licensed | [37] [38] [39] [40] [41] [42] |
| Polysaccharide conjugates | PedvaxHIB, ActHIB, HibTITER (H. influenzae) Prevnar 13, Pneumovax 23 (S. pneumoniae) Menactra, Menveo, Menomune (N. meningitidis) | Licensed Licensed Licensed | [43] [44,45] [46] |
| Viral vectors | 85A antigen (M. tuberculosis) | Licensed | [47] |
| Bacterial vectors | H. pylori (S. typhimurium) encoding urease A and B subunits Y. enterolica encoding bacterioferritin (B. abortus) L. monocytogenes encoding antigen 85 complex and MPB7MpT51 antigen (M. tuberculosis) | Experimental Experimental Experimental | [48] [49] [50] |
| Plasmid DNA | M. tuberculosis (hsp65 from M. leprae) B. anthracis (PA antigen) | Experimental Experimental | [51] [52] |
| BGs | Y. pestis S. typhimurium enteritides (BGs expressing flagellin) BGs carrying DNA for N. ghonorhea antigens | Experimental Experimental Experimental | [22] [53] [54] |
| OMVs | Bexsero/4CMenB, VA-MENGOC-BC, MeNZB, MenBVac (N. meningitidis serogroup B) B. pertussis BCG C. trachomatis V. cholerae M. smegmatis T. pallidum | Licensed Experimental Experimental Experimental Experimental Experimental Experimental | [55,56,57] [22] [22] [58] [58] [58] [58] |
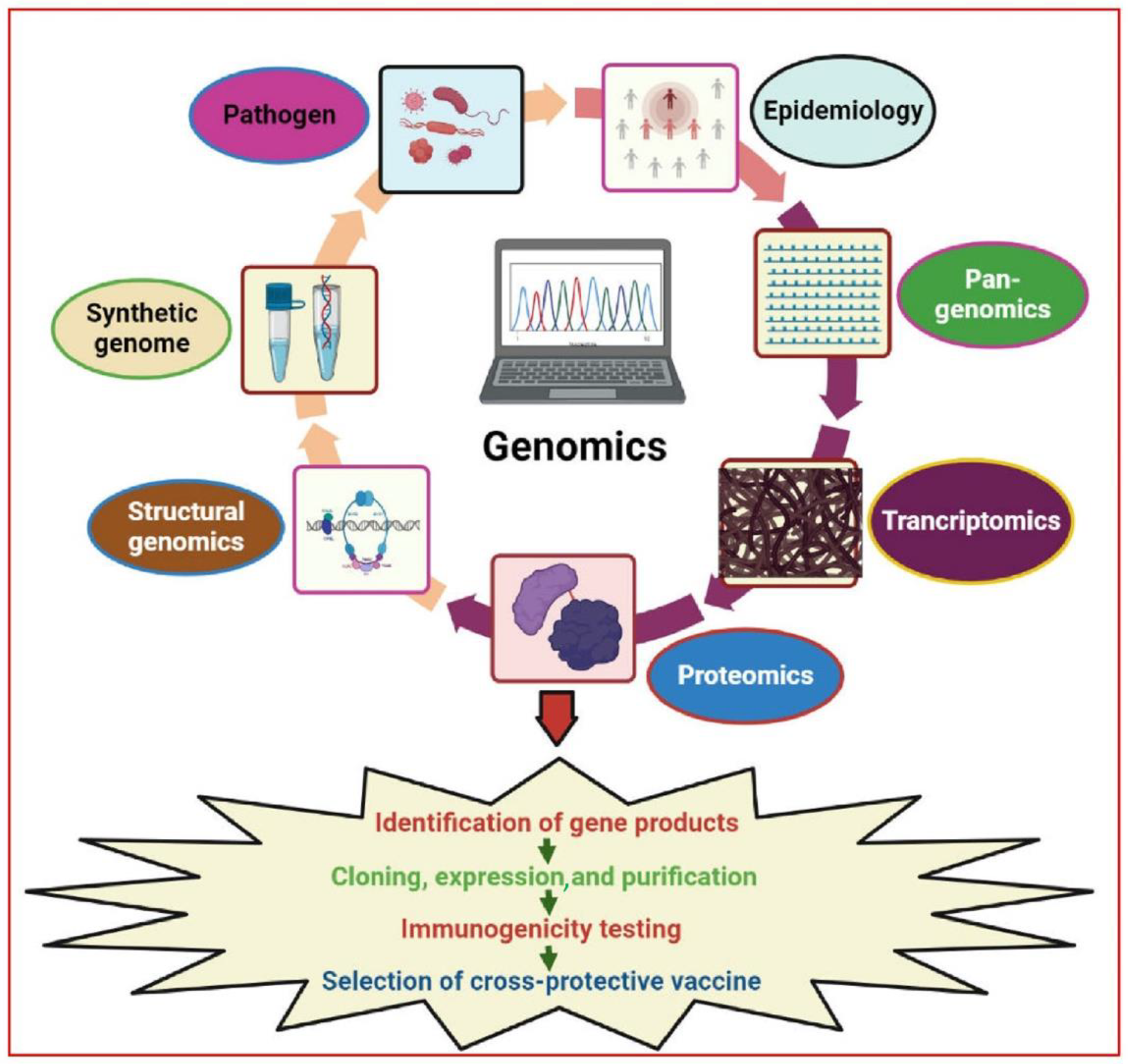
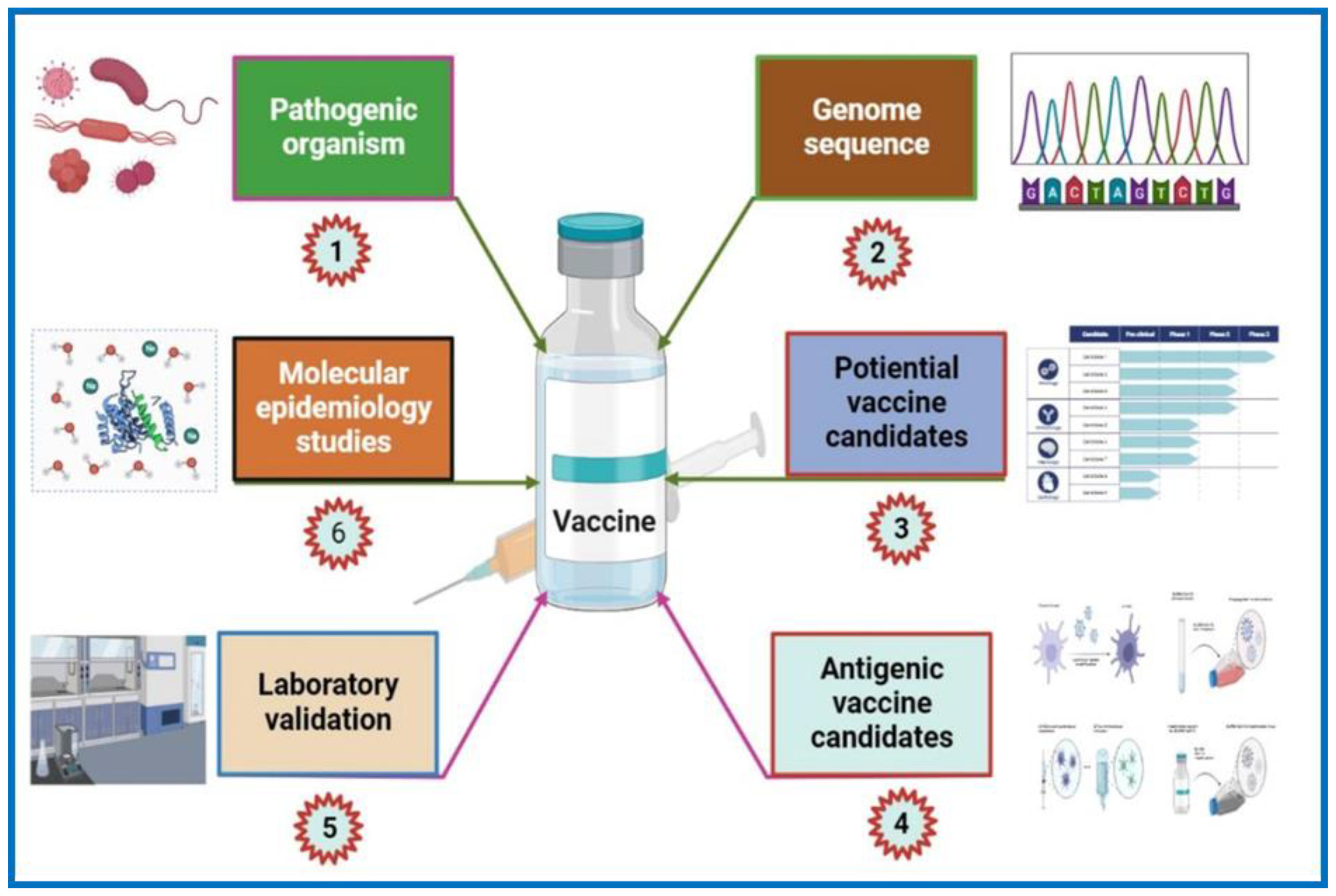
3. Genomics Revolution
4. Genome-Based Approaches
5. An In-Silico Approach: Reverse Vaccinology (RV)
6. Pan-Genomics Analysis or Comparative Genomics
7. Antigen Prioritization
8. Functional Genomics: Genes Essential for Vaccine Candidates
9. Transcriptomics: Expression Profile Identifies Potential Vaccines
10. Proteomics: A Genomic Complement for Vaccine Development
11. Next-Generation Epitope Mapping and Vaccine Design: Structural Genomics/Vaccinology
12. Synthetic Genomics: The Future of Vaccine Development
13. Conclusions and Future Direction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ribet, D.; Cossart, P. How Bacterial Pathogens Colonize Their Hosts and Invade Deeper Tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.T. Classical Labeling of Bacterial Pathogens According to Their Lifestyle in the Host: Inconsistencies and Alternatives. Front. Microbiol. 2012, 3, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurata, S. Extracellular and Intracellular Pathogen Recognition by Drosophila PGRP-LE and PGRP-LC. Int. Immunol. 2010, 22, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrand, J.; Ferrero, R.L. Recognition of Extracellular Bacteria by NLRs and Its Role in the Development of Adaptive Immunity. Front. Immunol. 2013, 4, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serruto, D.; Rappuoli, R. Post-Genomic Vaccine Development. FEBS Lett. 2006, 580, 2985–2992. [Google Scholar] [CrossRef] [Green Version]
- McNeil, M.M.; DeStefano, F. Vaccine-Associated Hypersensitivity. J. Allergy Clin. Immunol. 2018, 141, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Chambers, M.A.; Vordermeier, H.M.; Hewinson, R.G.; Lowrie, D.B.; Ertl, H.J.C. DNA Vaccines against Bacterial Pathogens. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2003; pp. 2000–2013. [Google Scholar]
- Pizza, M.; Bartoloni, A.; Prugnola, A.; Silvestri, S.; Rappuoli, R. Subunit S1 of Pertussis Toxin: Mapping of the Regions Essential for ADP-Ribosyltransferase Activity. Proc. Natl. Acad. Sci. USA 1988, 85, 7521–7525. [Google Scholar] [CrossRef] [Green Version]
- Fraser, C.M.; Rappuoli, R. Application of Microbial Genomic Science to Advanced Therapeutics. Annu. Rev. Med. 2005, 56, 459–474. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.-Y.; Abu Kwaik, Y. Apoptosis in Macrophages and Alveolar Epithelial Cells during Early Stages of Infection by Legionella pneumophila and Its Role in Cytopathogenicity. Infect. Immun. 1999, 67, 862–870. [Google Scholar] [CrossRef] [Green Version]
- Mahamed, D.; Boulle, M.; Ganga, Y.; Mc Arthur, C.; Skroch, S.; Oom, L.; Catinas, O.; Pillay, K.; Naicker, M.; Rampersad, S.; et al. Intracellular Growth of Mycobacterium tuberculosis after Macrophage Cell Death Leads to Serial Killing of Host Cells. Elife 2017, 6, e22028. [Google Scholar] [CrossRef]
- Richter-Dahlfors, A.; Buchan, A.M.J.; Finlay, B.B. Murine Salmonellosis Studied by Confocal Microscopy: Salmonella typhimurium Resides Intracellularly inside Macrophages and Exerts a Cytotoxic Effect on Phagocytes in Vivo. J. Exp. Med. 1997, 186, 569–580. [Google Scholar] [CrossRef] [Green Version]
- Bargen, K.v.; Gorvel, J.-P.; Salcedo, S.P. Internal Affairs: Investigating the Brucella Intracellular Lifestyle. FEMS Microbiol. Rev. 2012, 36, 533–562. [Google Scholar] [CrossRef] [Green Version]
- Birmingham, C.L.; Canadien, V.; Gouin, E.; Troy, E.B.; Yoshimori, T.; Cossart, P.; Higgins, D.E.; Brumell, J.H. Listeria Monocytogenes Evades Killing by Autophagy during Colonization of Host Cells. Autophagy 2007, 3, 442–451. [Google Scholar] [CrossRef] [Green Version]
- Kristof, M.N.; Allen, P.E.; Yutzy, L.D.; Thibodaux, B.; Paddock, C.D.; Martinez, J.J. Significant Growth by Rickettsia Species within Human Macrophage-like Cells Is a Phenotype Correlated with the Ability to Cause Disease in Mammals. Pathogens 2021, 10, 228. [Google Scholar] [CrossRef]
- Hilbi, H.; Moss, J.E.; Hersh, D.; Chen, Y.; Arondel, J.; Banerjee, S.; Flavell, R.A.; Yuan, J.; Sansonetti, P.J.; Zychlinsky, A. Shigella-Induced Apoptosis Is Dependent on Caspase-1 Which Binds to IpaB. J. Biol. Chem. 1998, 273, 32895–32900. [Google Scholar] [CrossRef] [Green Version]
- Lenz, S.M.; Ray, N.A.; Lema, T.; Collins, J.H.; Thapa, R.; Girma, S.; Balagon, M.; Bobosha, K.; Hagge, D.A.; Williams, D.L. Utility of a Mycobacterium leprae Molecular Viability Assay for Clinical Leprosy: An Analysis of Cases from the Philippines, Ethiopia, and Nepal. Front. Trop. Dis. 2022, 3, 967351. [Google Scholar] [CrossRef]
- Galán, J.E. Salmonella Typhimurium and Inflammation: A Pathogen-Centric Affair. Nat. Rev. Microbiol. 2021, 19, 716–725. [Google Scholar] [CrossRef]
- Kumar, Y.; Cocchiaro, J.; Valdivia, R.H. The Obligate Intracellular Pathogen Chlamydia trachomatis Targets Host Lipid Droplets. Curr. Biol. 2006, 16, 1646–1651. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.; Oh, M.H.; Jun, S.H.; Kim, S.I.; Choi, C.W.; Kwon, H.I.; Na, S.H.; Kim, Y.J.; Nicholas, A.; Selasi, G.N.; et al. Variation among Staphylococcus aureus Membrane Vesicle Proteomes Affects Cytotoxicity of Host Cells. Microb. Pathog. 2016, 93, 185–193. [Google Scholar] [CrossRef]
- Pucciarelli, M.G.; Garcia-del Portillo, F. Salmonella Intracellular Lifestyles and Their Impact on Host-to-host Transmission. Microb. Transm. 2019, 95–116. [Google Scholar]
- Osterloh, A. Vaccination against Bacterial Infections: Challenges, Progress, and New Approaches with a Focus on Intracellular Bacteria. Vaccines 2022, 10, 751. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, S.; Wolfe, D.N. Vaccination against Q Fever for Biodefense and Public Health Indications. Front. Microbiol. 2014, 5, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svennerholm, A.-M. From Cholera to Enterotoxigenic Escherichia coli (ETEC) Vaccine Development. Indian J. Med. Res. 2011, 133, 188–196. [Google Scholar] [PubMed]
- Osterloh, A. Vaccine Design and Vaccination Strategies against Rickettsiae. Vaccines 2021, 9, 896. [Google Scholar] [CrossRef]
- Walker, D.H. The Realities of Biodefense Vaccines against Rickettsia. Vaccine 2009, 27, D52–D55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machlaurin, A.; van der Pol, S.; Setiawan, D.; van der Werf, T.S.; Postma, M.J. Health Economic Evaluation of Current Vaccination Strategies and New Vaccines against Tuberculosis: A Systematic Review. Expert Rev. Vaccines 2019, 18, 897–911. [Google Scholar] [CrossRef] [Green Version]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against Invasive Salmonella Disease: Current Status and Future Directions. Hum. Vaccin. Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [Green Version]
- Beasley, D.W.C.; Brasel, T.L.; Comer, J.E. First Vaccine Approval under the FDA Animal Rule. NPJ Vaccines 2016, 1, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Mosley, J.F.; Smith, L.L.; Brantley, P.; Locke, D.; Como, M. Vaxchora: The First FDA-Approved Cholera Vaccination in the United States. Pharm. Ther. 2017, 42, 638. [Google Scholar]
- Kawamura, R.; Kasahara, S.; Toyama, T.; Nishinarita, F.; Tsubaki, S. On the Prevention of Tsutsugamushi. Results of Preventive Inoculations for People in the Endemic Region, and Laboratory Tests with the Pescadores Strain. Kitasato Arch. Exp. Med. 1939, 16, 93–109. [Google Scholar]
- Wisseman, C.L. Concepts of Louse-Borne Typhus Control in Developing Countries: The Use of the Living Attenuated E Strain Typhus Vaccine in Epidemic and Endemic Situations. In Immunity in Viral and Rickettsial Diseases; Springer: Boston, MA, USA, 1972; pp. 97–130. [Google Scholar]
- Safar, H.A.; Mustafa, A.S.; Amoudy, H.A.; El-Hashim, A. The Effect of Adjuvants and Delivery Systems on Th1, Th2, Th17 and Treg Cytokine Responses in Mice Immunized with Mycobacterium tuberculosis-Specific Proteins. PLoS ONE 2020, 15, e0228381. [Google Scholar] [CrossRef]
- Crocquet-Valdes, P.A.; Díaz-Montero, C.M.; Feng, H.M.; Li, H.; Barrett, A.D.T.; Walker, D.H. Immunization with a Portion of Rickettsial Outer Membrane Protein A Stimulates Protective Immunity against Spotted Fever Rickettsiosis. Vaccine 2001, 20, 979–988. [Google Scholar] [CrossRef]
- Xiong, X.; Jiao, J.; Gregory, A.E.; Wang, P.; Bi, Y.; Wang, X.; Jiang, Y.; Wen, B.; Portnoy, D.A.; Samuel, J.E.; et al. Identification of Coxiella burnetii CD8+ T-Cell Epitopes and Delivery by Attenuated Listeria monocytogenes as a Vaccine Vector in a C57BL/6 Mouse Model. J. Infect. Dis. 2017, 215, 1580–1589. [Google Scholar]
- Xu, C.; Zhang, B.-Z.; Lin, Q.; Deng, J.; Yu, B.; Arya, S.; Yuen, K.Y.; Huang, J.-D. Live Attenuated Salmonella typhimurium Vaccines Delivering SaEsxA and SaEsxB via Type III Secretion System Confer Protection against Staphylococcus aureus Infection. BMC Infect. Dis. 2018, 18, 195. [Google Scholar] [CrossRef]
- Dhillon, S.; Keam, S.J. DTaP-IPV/Hib Vaccine (Pentacel®). Pediatr. Drugs 2008, 10, 405–416. [Google Scholar] [CrossRef]
- Gorse, G.J.; Keitel, W.; Keyserling, H.; Taylor, D.N.; Lock, M.; Alves, K.; Kenner, J.; Deans, L.; Gurwith, M. Immunogenicity and Tolerance of Ascending Doses of a Recombinant Protective Antigen (RPA102) Anthrax Vaccine: A Randomized, Double-Blinded, Controlled, Multicenter Trial. Vaccine 2006, 24, 5950–5959. [Google Scholar] [CrossRef]
- Venkatasubramaniam, A.; Adhikari, R.P.; Kort, T.; Liao, G.C.; Conley, S.; Abaandou, L.; Kailasan, S.; Onodera, Y.; Krishnan, S.; Djagbare, D.M.; et al. TBA225, a Fusion Toxoid Vaccine for Protection and Broad Neutralization of Staphylococcal Superantigens. Sci. Rep. 2019, 9, 3279. [Google Scholar] [CrossRef] [Green Version]
- Keitel, W.A. Recombinant Protective Antigen 102 (RPA102): Profile of a Second-Generation Anthrax Vaccine. Expert Rev. Vaccines 2006, 5, 417–430. [Google Scholar] [CrossRef]
- Karauzum, H.; Adhikari, R.P.; Sarwar, J.; Devi, V.S.; Abaandou, L.; Haudenschild, C.; Mahmoudieh, M.; Boroun, A.R.; Vu, H.; Nguyen, T. Structurally Designed Attenuated Subunit Vaccines for S aureus LukS-PV and LukF-PV Confer Protection in a Mouse Bacteremia Model. PLoS ONE 2013, 8, e65384. [Google Scholar]
- MacNeil, J.R.; Rubin, L.; Folaranmi, T.; Ortega-Sanchez, I.R.; Patel, M.; Martin, S.W. Use of Serogroup B Meningococcal Vaccines in Adolescents and Young Adults: Recommendations of the Advisory Committee on Immunization Practices, 2015. Morb. Mortal. Wkly. Rep. 2015, 64, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Zangwill, K.M.; Wenger, J.D.; Sutter, R.W.; Hadler, S.C. Recommendations for Use of Haemophilus b Conjugate Vaccines and a Combined Diphtheria, Tetanus, Pertussis, and Haemophilus b Vaccine: Recommendations of the Advisory Committee on Immunization Practices (ACIP). Morb. Mortal. Wkly. Rep. Recomm. Rep. 1993, 42, 1–15. [Google Scholar]
- Abraham-Van Parijs, B.; Malinoski, F.J. Post-Marketing Effectiveness of Prevnar [Pneumococcal 7-Valent Conjugate Vaccine (Diphtheria CRM197 Protein)] and Implications for Adult Immunization. Mech. Ageing Dev. 2004, 125, 147–148. [Google Scholar] [CrossRef]
- Food and Drug Administration. PNEUMOVAX® 23 (Pneumococcal Vaccine Polyvalent). 1986. Available online: https://www.fda.gov/vaccines-blood-biologics/vaccines/pneumovax-23-pneumococcal-vaccine-polyvalent (accessed on 15 December 2022).
- Pizza, M.; Bekkat-Berkani, R.; Rappuoli, R. Vaccines against Meningococcal Diseases. Microorganisms 2020, 8, 1521. [Google Scholar] [CrossRef] [PubMed]
- Volz, A.; Sutter, G. Modified Vaccinia Virus Ankara: History, Value in Basic Research, and Current Perspectives for Vaccine Development. Adv. Virus Res. 2017, 97, 187–243. [Google Scholar] [PubMed]
- Corthésy-Theulaz, I.E.; Hopkins, S.; Bachmann, D.; Saldinger, P.F.; Porta, N.; Haas, R.; Zheng-Xin, Y.; Meyer, T.; Bouzourène, H.; Blum, A.L.; et al. Mice Are Protected from Helicobacter pylori Infection by Nasal Immunization with Attenuated Salmonella typhimurium PhoPc Expressing Urease A and B Subunits. Infect. Immun. 1998, 66, 581–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Mariri, A.; Tibor, A.; Lestrate, P.; Mertens, P.; Bolle, X.D.; Letesson, J.-J. Yersinia enterocolitica as a Vehicle for a Naked DNA Vaccine Encoding Brucella abortus Bacterioferritin or P39 Antigen. Infect. Immun. 2002, 70, 1915–1923. [Google Scholar] [CrossRef]
- Miki, K.; Nagata, T.; Tanaka, T.; Kim, Y.-H.; Uchijima, M.; Ohara, N.; Nakamura, S.; Okada, M.; Koide, Y. Induction of Protective Cellular Immunity against Mycobacterium tuberculosis by Recombinant Attenuated Self-Destructing Listeria monocytogenes Strains Harboring Eukaryotic Expression Plasmids for Antigen 85 Complex and MPB/MPT51. Infect. Immun. 2004, 72, 2014–2021. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.L.; Bonato, V.L.D.; Coelho-Castelo, A.A.M.; DeSouza, A.O.; Santos, S.A.; Lima, K.M.; Faccioli, L.H.; Rodrigues, J.M. Immunotherapy with Plasmid DNA Encoding Mycobacterial Hsp65 in Association with Chemotherapy Is a More Rapid and Efficient Form of Treatment for Tuberculosis in Mice. Gene Ther. 2005, 12, 281–287. [Google Scholar] [CrossRef]
- Gu, M.-L.; Leppla, S.H.; Klinman, D.M. Protection against Anthrax Toxin by Vaccination with a DNA Plasmid Encoding Anthrax Protective Antigen. Vaccine 1999, 17, 340–344. [Google Scholar] [CrossRef]
- Senevirathne, A.; Hewawaduge, C.; Lee, J.H. Immunization of Chicken with Flagellin Adjuvanted Salmonella enteritidis Bacterial Ghosts Confers Complete Protection against Chicken Salmonellosis. Poult. Sci. 2021, 100, 101205. [Google Scholar] [CrossRef]
- Jiao, H.; Yang, H.; Zheng, W.; Zhang, Q.; Zhao, D.; Li, G. Enhancement of Immune Responses by Co-administration of Bacterial Ghosts-mediated Neisseria Gonorrhoeae DNA Vaccines. J. Appl. Microbiol. 2021, 130, 1770–1777. [Google Scholar] [CrossRef]
- Sierra, G.V.; Campa, H.C.; Varcacel, N.M.; Garcia, I.L.; Izquierdo, P.L.; Sotolongo, P.F.; Casanueva, G.V.; Rico, C.O.; Rodriguez, C.R.; Terry, M.H. Vaccine against Group B Neisseria meningitidis: Protection Trial and Mass Vaccination Results in Cuba. NIPH Ann. 1991, 14, 195–207. [Google Scholar]
- O’Ryan, M.; Stoddard, J.; Toneatto, D.; Wassil, J.; Dull, P.M. A Multi-Component Meningococcal Serogroup B Vaccine (4CMenB): The Clinical Development Program. Drugs 2014, 74, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; MacLennan, C.A. Outer Membrane Vesicle Vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar]
- Chbib, C.; Shah, S.M.; Gala, R.P.; Uddin, M.N. Potential Applications of Microparticulate-Based Bacterial Outer Membrane Vesicles (OMVs) Vaccine Platform for Sexually Transmitted Diseases (STDs): Gonorrhea, Chlamydia, and Syphilis. Vaccines 2021, 9, 1245. [Google Scholar] [CrossRef]
- Fleischmann, R.D.; Adams, M.D.; White, O.; Clayton, R.A.; Kirkness, E.F.; Kerlavage, A.R.; Bult, C.J.; Tomb, J.F.; Dougherty, B.A.; Merrick, J.M.; et al. Whole-Genome Random Sequencing and Assembly of Haemophilus Influenzae Rd. Science 1995, 269, 496–512. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, D.K.; Sehgal, D. Developing Antibacterial Vaccines in Genomics and Proteomics Era. Scand. J. Immunol. 2008, 67, 544–552. [Google Scholar] [CrossRef]
- Casanova, J.-L.; Abel, L. Human Genetics of Infectious Diseases: A Unified Theory. EMBO J. 2007, 26, 915–922. [Google Scholar] [CrossRef]
- Burgner, D.; Jamieson, S.E.; Blackwell, J.M. Genetic Susceptibility to Infectious Diseases: Big Is Beautiful, but Will Bigger Be Even Better? Lancet Infect. Dis. 2006, 6, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Seib, K.L.; Dougan, G.; Rappuoli, R. The Key Role of Genomics in Modern Vaccine and Drug Design for Emerging Infectious Diseases. PLoS Genet. 2009, 5, e1000612. [Google Scholar] [CrossRef] [Green Version]
- Matić, Z.; Šantak, M. Current View on Novel Vaccine Technologies to Combat Human Infectious Diseases. Appl. Microbiol. Biotechnol. 2021, 106, 25–56. [Google Scholar] [CrossRef]
- Nakamura, S.; Yang, C.-S.; Sakon, N.; Ueda, M.; Tougan, T.; Yamashita, A.; Goto, N.; Takahashi, K.; Yasunaga, T.; Ikuta, K.; et al. Direct Metagenomic Detection of Viral Pathogens in Nasal and Fecal Specimens Using an Unbiased High-Throughput Sequencing Approach. PLoS ONE 2009, 4, e4219. [Google Scholar] [CrossRef] [PubMed]
- Bittar, F.; Richet, H.; Dubus, J.-C.; Reynaud-Gaubert, M.; Stremler, N.; Sarles, J.; Raoult, D.; Rolain, J.-M. Molecular Detection of Multiple Emerging Pathogens in Sputa from Cystic Fibrosis Patients. PLoS ONE 2008, 3, e2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaudo, C.D.; Telford, J.L.; Rappuoli, R.; Seib, K.L. Vaccinology in the Genome Era. J. Clin. Invest. 2009, 119, 2515–2525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucci, M.J. Novel Genetic Techniques and Approaches in the Microbial Genomics Era. Drugs R D 2007, 8, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.D. When Will the Genomics Investment Pay off for Antibacterial Discovery? Biochem. Pharmacol. 2006, 71, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Voorhis, W.C.V.; Hol, W.G.J.; Myler, P.J.; Stewart, L.J. The Role of Medical Structural Genomics in Discovering New Drugs for Infectious Diseases. PLoS Comput. Biol. 2009, 5, e1000530. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Sanduja, P.; Anand, A.; Mahajan, P.; Guzman, C.A.; Yadav, P.; Awasthi, A.; Hanski, E.; Dua, M.; Johri, A.K. Advanced Strategies for Development of Vaccines against Human Bacterial Pathogens. World J. Microbiol. Biotechnol. 2021, 37, 67. [Google Scholar] [CrossRef]
- Gong, W.; Pan, C.; Cheng, P.; Wang, J.; Zhao, G.; Wu, X. Peptide-Based Vaccines for Tuberculosis. Front. Immunol. 2022, 13, 830497. [Google Scholar] [CrossRef]
- Soria-Guerra, R.E.; Nieto-Gomez, R.; Govea-Alonso, D.O.; Rosales-Mendoza, S. An Overview of Bioinformatics Tools for Epitope Prediction: Implications on Vaccine Development. J. Biomed. Inform. 2015, 53, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Özdemir, N.; Kök-Taş, T.; Guzel-Seydim, Z. Effect of Gluconacetobacter Spp. on Kefir Grains and Kefir Quality. Food Sci. Biotechnol. 2015, 24, 99–106. [Google Scholar] [CrossRef]
- Gupta, S.; Chavan, S.; Deobagkar, D.N.; Deobagkar, D.D. Bio/Chemoinformatics in India: An Outlook. Brief. Bioinform. 2015, 16, 710–731. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wang, P.; Kim, Y.; Haste-Andersen, P.; Beaver, J.; Bourne, P.E.; Bui, H.-H.; Buus, S.; Frankild, S.; Greenbaum, J.; et al. Immune Epitope Database Analysis Resource (IEDB-AR). Nucleic Acids Res. 2008, 36, W513–W518. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Gomase, V. In-Silico Insights to Predict the Major Histocompatibility Complex Peptide Binders from Protein. Eur. J. Mol. Clin. Med. 2021, 8, 2021. [Google Scholar]
- Frenkel, L.D. The Global Burden of Vaccine-Preventable Infectious Diseases in Children Less than 5 Years of Age: Implications for COVID-19 Vaccination. How Can We Do Better? In Allergy and Asthma Proceedings; OceanSide Publications, Inc.: Providence, RI, USA, 2021; Volume 42, pp. 378–385. [Google Scholar]
- Seib, K.L.; Zhao, X.; Rappuoli, R. Developing Vaccines in the Era of Genomics: A Decade of Reverse Vaccinology. Clin. Microbiol. Infect. 2012, 18, 109–116. [Google Scholar] [CrossRef] [Green Version]
- Rappuoli, R. Reverse Vaccinology, a Genome-Based Approach to Vaccine Development. Vaccine 2001, 19, 2688–2691. [Google Scholar] [CrossRef]
- Goldschneider, I.; Gotschlich, E.C.; Artenstein, M.S. Human Immunity to the Meningococcus: I. The Role of Humoral Antibodies. J. Exp. Med. 1969, 129, 1307–1326. [Google Scholar] [CrossRef]
- Ross, S.C.; Rosenthal, P.J.; Berberich, H.M.; Densen, P. Killing of Neisseria meningitidis by Human Neutrophils: Implications for Normal and Complement-Deficient Individuals. J. Infect. Dis. 1987, 155, 1266–1275. [Google Scholar] [CrossRef]
- Pizza, M.; Scarlato, V.; Masignani, V.; Giuliani, M.M.; Arico, B.; Comanducci, M.; Jennings, G.T.; Baldi, L.; Bartolini, E.; Capecchi, B.; et al. Identification of Vaccine Candidates against Serogroup B Meningococcus by Whole-Genome Sequencing. Science 2000, 287, 1816–1820. [Google Scholar] [CrossRef]
- Tettelin, H.; Saunders, N.J.; Heidelberg, J.; Jeffries, A.C.; Nelson, K.E.; Eisen, J.A.; Ketchum, K.A.; Hood, D.W.; Peden, J.F.; Dodson, R.J.; et al. Complete Genome Sequence of Neisseria meningitidis Serogroup B Strain MC58. Science 2000, 287, 1809–1815. [Google Scholar] [CrossRef] [Green Version]
- Romero, J.D.; Outschoorn, I.M. Current Status of Meningococcal Group B Vaccine Candidates: Capsular or Noncapsular? Clin. Microbiol. Rev. 1994, 7, 559–575. [Google Scholar] [CrossRef]
- Giuliani, M.M.; Adu-Bobie, J.; Comanducci, M.; Aricò, B.; Savino, S.; Santini, L.; Brunelli, B.; Bambini, S.; Biolchi, A.; Capecchi, B.; et al. A Universal Vaccine for Serogroup B Meningococcus. Proc. Natl. Acad. Sci. USA 2006, 103, 10834–10839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarti, D.N.; Fiske, M.J.; Fletcher, L.D.; Zagursky, R.J. Application of Genomics and Proteomics for Identification of Bacterial Gene Products as Potential Vaccine Candidates. Vaccine 2000, 19, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Ross, B.C.; Czajkowski, L.; Hocking, D.; Margetts, M.; Webb, E.; Rothel, L.; Patterson, M.; Agius, C.; Camuglia, S.; Reynolds, E.; et al. Identification of Vaccine Candidate Antigens from a Genomic Analysis of Porphyromonas gingivalis. Vaccine 2001, 19, 4135–4142. [Google Scholar] [CrossRef] [PubMed]
- Wizemann, T.M.; Heinrichs, J.H.; Adamou, J.E.; Erwin, A.L.; Kunsch, C.; Choi, G.H.; Barash, S.C.; Rosen, C.A.; Masure, H.R.; Tuomanen, E.; et al. Use of a Whole Genome Approach to Identify Vaccine Molecules Affording Protection against Streptococcus pneumoniae Infection. Infect. Immun. 2001, 69, 1593–1598. [Google Scholar] [CrossRef] [Green Version]
- Ariel, N.; Zvi, A.; Grosfeld, H.; Gat, O.; Inbar, Y.; Velan, B.; Cohen, S.; Shafferman, A. Search for Potential Vaccine Candidate Open Reading Frames in the Bacillus anthracis Virulence Plasmid PXO1: In Silico and in Vitro Screening. Infect. Immun. 2002, 70, 6817–6827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montigiani, S.; Falugi, F.; Scarselli, M.; Finco, O.; Petracca, R.; Galli, G.; Mariani, M.; Manetti, R.; Agnusdei, M.; Cevenini, R.; et al. Genomic Approach for Analysis of Surface Proteins in Chlamydia Pneumoniae. Infect. Immun. 2002, 70, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Al-Hasani, K.; Boyce, J.; McCarl, V.P.; Bottomley, S.; Wilkie, I.; Adler, B. Identification of Novel Immunogens in Pasteurella multocida. Microb. Cell Fact. 2007, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Bambini, S.; Rappuoli, R. The Use of Genomics in Microbial Vaccine Development. Drug Discov. Today 2009, 14, 252–260. [Google Scholar] [CrossRef]
- Betts, J.C. Transcriptomics and Proteomics: Tools for the Identification of Novel Drug Targets and Vaccine Candidates for Tuberculosis. IUBMB Life 2002, 53, 239–242. [Google Scholar] [CrossRef]
- Maione, D.; Margarit, I.; Rinaudo, C.D.; Masignani, V.; Mora, M.; Scarselli, M.; Tettelin, H.; Brettoni, C.; Iacobini, E.T.; Rosini, R.; et al. Identification of a Universal Group B Streptococcus Vaccine by Multiple Genome Screen. Science 2005, 309, 148–150. [Google Scholar] [CrossRef] [Green Version]
- Mora, M.; Bensi, G.; Capo, S.; Falugi, F.; Zingaretti, C.; Manetti, A.G.O.; Maggi, T.; Taddei, A.R.; Grandi, G.; Telford, J.L. Group A Streptococcus Produce Pilus-like Structures Containing Protective Antigens and Lancefield T Antigens. Proc. Natl. Acad. Sci. USA 2005, 102, 15641–15646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, M.W. Molecular Mimicry, Autoimmunity, and Infection: The Cross-Reactive Antigens of Group A Streptococci and Their Sequelae. Microbiol. Spectr. 2019, 7, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Damian, R.T. Molecular Mimicry: Antigen Sharing by Parasite and Host and Its Consequences. Am. Nat. 1964, 98, 129–149. [Google Scholar] [CrossRef]
- Zabriskie, J.B. Mimetic Relationships between Group A Streptococci and Mammalian Tissues. Adv. Immunol. 1967, 7, 147–188. [Google Scholar] [PubMed]
- Guilherme, L.; Kalil, J.; Cunningham, M. Molecular Mimicry in the Autoimmune Pathogenesis of Rheumatic Heart Disease. Autoimmunity 2006, 39, 31–39. [Google Scholar] [CrossRef]
- Dale, J.B.; Walker, M.J. Update on Group A Streptococcal Vaccine Development. Curr. Opin. Infect. Dis. 2020, 33, 244–250. [Google Scholar] [CrossRef]
- Thorpe, C.; Edwards, L.; Snelgrove, R.; Finco, O.; Rae, A.; Grandi, G.; Guilio, R.; Hussell, T. Discovery of a Vaccine Antigen That Protects Mice from Chlamydia pneumoniae Infection. Vaccine 2007, 25, 2252–2260. [Google Scholar] [CrossRef]
- Noor, F.; Ahmad, S.; Saleem, M.; Alshaya, H.; Qasim, M.; Rehman, A.; Ehsan, H.; Talib, N.; Saleem, H.; Jardan, Y.A.B.; et al. Designing a Multi-Epitope Vaccine against Chlamydia pneumoniae by Integrating the Core Proteomics, Subtractive Proteomics and Reverse Vaccinology-Based Immunoinformatics Approaches. Comput. Biol. Med. 2022, 145, 105507. [Google Scholar] [CrossRef]
- Chaudhuri, R.; Kulshreshtha, D.; Raghunandanan, M.V.; Ramachandran, S. Integrative Immunoinformatics for Mycobacterial Diseases in R Platform. Syst. Synth. Biol. 2014, 8, 27–39. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Racz, R.; Sayers, S.; Lin, Y.; Todd, T.; Hur, J.; Li, X.; Patel, M.; Zhao, B.; Chung, M.; et al. Updates on the Web-Based VIOLIN Vaccine Database and Analysis System. Nucleic Acids Res. 2014, 42, D1124–D1132. [Google Scholar] [CrossRef] [Green Version]
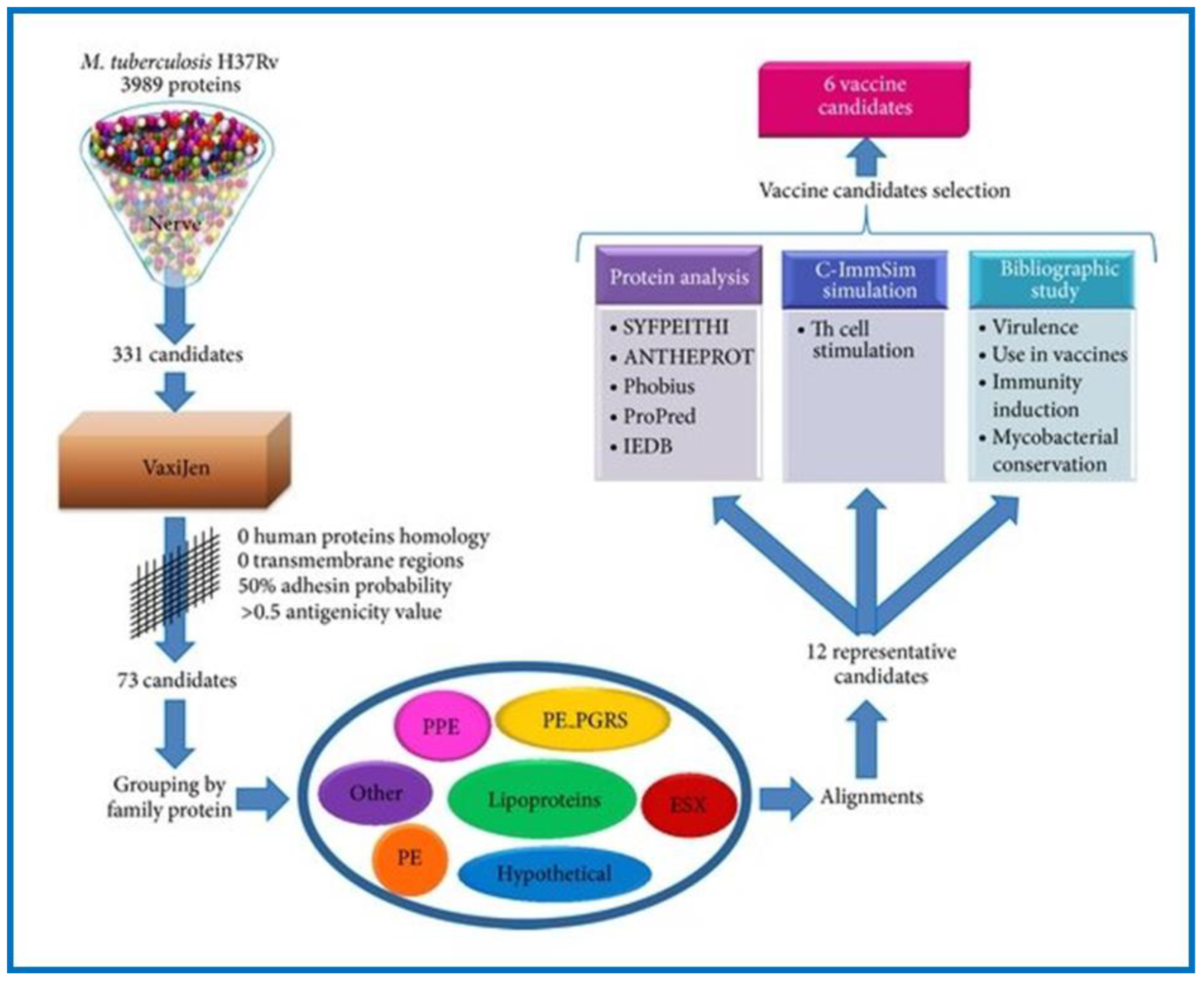
- Monterrubio-López, G.P.; González-Y-Merchand, J.A.; Ribas-Aparicio, R.M. Identification of Novel Potential Vaccine Candidates against Tuberculosis Based on Reverse Vaccinology. Biomed Res. Int. 2015, 2015, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delogu, G.; Pusceddu, C.; Bua, A.; Fadda, G.; Brennan, M.J.; Zanetti, S. Rv1818c-encoded PE_PGRS Protein of Mycobacterium tuberculosis Is Surface Exposed and Influences Bacterial Cell Structure. Mol. Microbiol. 2004, 52, 725–733. [Google Scholar] [CrossRef]
- Sharma, A.; Rajan, G.S.; Kharb, R.; Biswas, S. Genome Wide Analysis of Chlamydia pneumoniae for Candidate Vaccine Development. Curr. Comput. Aided. Drug Des. 2016, 12, 206–215. [Google Scholar] [CrossRef]
- Indrabalan, U.B.; Puttahonnappa, S.K.; Beelagi, M.S.; Patil, S.S.; Shivamallu, C.; Pappana, M.; Amachawadi, R. Reverse Vaccinology Based in Silico Analysis of Epitope Prediction in Cya, Lef and PagA Genes from Bacillus anthracis against Anthrax Infected Species: An Immunoinformatics Approach. Chem. Biol. Lett. 2022, 9, 295. [Google Scholar]
- Caro-Gomez, E.; Gazi, M.; Goez, Y.; Valbuena, G. Discovery of Novel Cross-Protective Rickettsia Prowazekii T-Cell Antigens Using a Combined Reverse Vaccinology and in Vivo Screening Approach. Vaccine 2014, 32, 4968–4976. [Google Scholar] [CrossRef] [Green Version]
- Kale, S.; Gore, D. Ehrlichia ruminantium Vaccine Epitope Search by Reverse Vaccinology Approach. J. Pharm. Res. 2017, 11, 807–810. [Google Scholar]
- Zai, X.; Yin, Y.; Guo, F.; Yang, Q.; Li, R.; Li, Y.; Zhang, J.; Xu, J.; Chen, W. Screening of Potential Vaccine Candidates against Pathogenic Brucella Spp. Using Compositive Reverse Vaccinology. Vet. Res. 2021, 52, 75. [Google Scholar] [CrossRef]
- Aslam, S.; Ashfaq, U.A.; Zia, T.; Aslam, N.; Alrumaihi, F.; Shahid, F.; Noor, F.; Qasim, M. Proteome Based Mapping and Reverse Vaccinology Techniques to Contrive Multi-Epitope Based Subunit Vaccine (MEBSV) against Streptococcus pyogenes. Infect. Genet. Evol. 2022, 100, 105259. [Google Scholar] [CrossRef]
- Jalal, K.; Khan, K.; Ahmad, D.; Hayat, A.; Basharat, Z.; Abbas, M.N.; Alghamdi, S.; Almehmadi, M.; Sahibzada, M.U.K. Pan-Genome Reverse Vaccinology Approach for the Design of Multi-Epitope Vaccine Construct against Escherichia albertii. Int. J. Mol. Sci. 2021, 22, 12814. [Google Scholar] [CrossRef]
- Mora, M.; Donati, C.; Medini, D.; Covacci, A.; Rappuoli, R. Microbial Genomes and Vaccine Design: Refinements to the Classical Reverse Vaccinology Approach. Curr. Opin. Microbiol. 2006, 9, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Vernikos, G.S. Overtake in Reverse Gear. Nat. Rev. Microbiol. 2008, 6, 334–335. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Shan, S.; Zainab, B.; Ayaz, Z.; He, J.; Xie, Z.; Rashid, U.; Zhang, D.; Abbasi, A.M. Novel Vaccine Design Based on Genomics Data Analysis: A Review. Scand. J. Immunol. 2021, 93, e12986. [Google Scholar] [CrossRef]
- Bombaci, M.; Grifantini, R.; Mora, M.; Reguzzi, V.; Petracca, R.; Meoni, E.; Balloni, S.; Zingaretti, C.; Falugi, F.; Manetti, A.G.O.; et al. Protein Array Profiling of Tic Patient Sera Reveals a Broad Range and Enhanced Immune Response against Group A Streptococcus Antigens. PLoS ONE 2009, 4, e6332. [Google Scholar] [CrossRef] [Green Version]
- Anani, H.; Zgheib, R.; Hasni, I.; Raoult, D.; Fournier, P.-E. Interest of Bacterial Pangenome Analyses in Clinical Microbiology. Microb. Pathog. 2020, 149, 104275. [Google Scholar] [CrossRef]
- Binnewies, T.T.; Motro, Y.; Hallin, P.F.; Lund, O.; Dunn, D.; La, T.; Hampson, D.J.; Bellgard, M.; Wassenaar, T.M.; Ussery, D.W. Ten Years of Bacterial Genome Sequencing: Comparative-Genomics-Based Discoveries. Funct. Integr. Genom. 2006, 6, 165–185. [Google Scholar] [CrossRef]
- Fukiya, S.; Mizoguchi, H.; Tobe, T.; Mori, H. Extensive Genomic Diversity in Pathogenic Escherichia coli and Shigella Strains Revealed by Comparative Genomic Hybridization Microarray. J. Bacteriol. 2004, 186, 3911–3921. [Google Scholar] [CrossRef] [Green Version]
- Tettelin, H.; Masignani, V.; Cieslewicz, M.J.; Donati, C.; Medini, D.; Ward, N.L.; Angiuoli, S.V.; Crabtree, J.; Jones, A.L.; Durkin, A.S.; et al. Genome Analysis of Multiple Pathogenic Isolates of Streptococcus agalactiae: Implications for the Microbial “Pan-Genome". Proc. Natl. Acad. Sci. USA 2005, 102, 13950–13955. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.R.; McIntyre, L.; Mutreja, A.; Lacey, J.A.; Lees, J.A.; Towers, R.J.; Duchêne, S.; Smeesters, P.R.; Frost, H.R.; Price, D.J.; et al. Atlas of Group A Streptococcal Vaccine Candidates Compiled Using Large-Scale Comparative Genomics. Nat. Genet. 2019, 51, 1035–1043. [Google Scholar] [CrossRef] [Green Version]
- Kavvas, E.S.; Catoiu, E.; Mih, N.; Yurkovich, J.T.; Seif, Y.; Dillon, N.; Heckmann, D.; Anand, A.; Yang, L.; Nizet, V.; et al. Machine Learning and Structural Analysis of Mycobacterium tuberculosis Pan-Genome Identifies Genetic Signatures of Antibiotic Resistance. Nat. Commun. 2018, 9, 4306. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zhong, J.; Zhang, J.; Li, C.; Yu, X.; Xiao, J.; Jia, X.; Ding, N.; Ma, G.; Wang, G.; et al. Pan-Genomic Study of Mycobacterium tuberculosis Reflecting the Primary/Secondary Genes, Generality/Individuality, and the Interconversion through Copy Number Variations. Front. Microbiol. 2018, 9, 1886. [Google Scholar] [CrossRef]
- Forrellad, M.A.; Klepp, L.I.; Gioffré, A.; Sabio y Garcia, J.; Morbidoni, H.R.; Santangelo, M.d.l.P.; Cataldi, A.A.; Bigi, F. Virulence Factors of the Mycobacterium tuberculosis Complex. Virulence 2013, 4, 3–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubnau, E.; Smith, I. Mycobacterium tuberculosis Gene Expression in Macrophages. Microbes Infect. 2003, 5, 629–637. [Google Scholar] [CrossRef]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA-an Ultra-Fast Pan-Genome Analysis Pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albutti, A. An Integrated Computational Framework to Design a Multi-Epitopes Vaccine against Mycobacterium tuberculosis. Sci. Rep. 2021, 11, 21929. [Google Scholar] [CrossRef] [PubMed]
- Chand, Y.; Alam, M.A.; Singh, S. Pan-Genomic Analysis of the Species Salmonella enterica: Identification of Core Essential and Putative Essential Genes. Gene Rep. 2020, 20, 100669. [Google Scholar] [CrossRef]
- Seribelli, A.A.; Gonzales, J.C.; Almeida, F.d.; Benevides, L.; Medeiros, M.I.C.; Rodrigues, D.D.d.P.; Soares, S.d.C.; Allard, M.W.; Falcão, J.P. Phylogenetic Analysis Revealed That Salmonella typhimurium ST313 Isolated from Humans and Food in Brazil Presented a High Genomic Similarity. Braz. J. Microbiol. 2020, 51, 53–64. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, H.; Chen, R.; Qin, T.; Ren, H.; Liu, B.; Ding, X.; Sha, D.; Zhou, W. Legionnaires’ Disease Caused by Legionella pneumophila Serogroups 5 and 10, China. Emerg. Infect. Dis. 2014, 20, 1242–1243. [Google Scholar] [CrossRef]
- Mercante, J.W.; Winchell, J.M. Current and Emerging Legionella Diagnostics for Laboratory and Outbreak Investigations. Clin. Microbiol. Rev. 2015, 28, 95–133. [Google Scholar] [CrossRef] [Green Version]
- Shah, I.; Jamil, S.; Rehmat, S.; Butt, H.A.; Ali, S.S.; Idrees, M.; Zhan, Y.; Hussain, Z.; Ali, S.; Waseem, M.; et al. Evaluation and Identification of Essential Therapeutic Proteins and Vaccinomics Approach towards Multi-Epitopes Vaccine Designing against Legionella pneumophila for Immune Response Instigation. Comput. Biol. Med. 2022, 143, 105291. [Google Scholar] [CrossRef]
- Felice, A.G.; Alves, L.G.; Freitas, A.S.F.; Rodrigues, T.C.V.; Jaiswal, A.K.; Tiwari, S.; Gomes, L.G.R.; Miranda, F.M.; Ramos, R.T.J.; Azevedo, V.; et al. Pan-Genomic Analyses of 47 Complete Genomes of the Rickettsia Genus and Prediction of New Vaccine Targets and Virulence Factors of the Species. J. Biomol. Struct. Dyn. 2021, 7496–7510. [Google Scholar] [CrossRef]
- Parajuli, P.; Deimel, L.P.; Verma, N.K. Genome Analysis of Shigella flexneri Serotype 3b Strain SFL1520 Reveals Significant Horizontal Gene Acquisitions Including a Multidrug Resistance Cassette. Genome Biol. Evol. 2019, 11, 776–785. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.; Gomes, J.P. Evolution, Phylogeny, and Molecular Epidemiology of Chlamydia. Infect. Genet. Evol. 2014, 23, 49–64. [Google Scholar] [CrossRef]
- Sigalova, O.M.; Chaplin, A.V.; Bochkareva, O.O.; Shelyakin, P.V.; Filaretov, V.A.; Akkuratov, E.E.; Burskaia, V.; Gelfand, M.S. Chlamydia Pan-Genomic Analysis Reveals Balance between Host Adaptation and Selective Pressure to Genome Reduction. BMC Genom. 2019, 20, 710. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Naz, A.; Soares, S.C.; Bakhtiar, M.; Tiwari, S.; Hassan, S.S.; Hanan, F.; Ramos, R.; Pereira, U.; Barh, D.; et al. Pan-Genome Analysis of Human Gastric Pathogen, H. pylori: Comparative Genomics and Pathogenomics Approaches to Identify Regions Associated with Pathogenicity and Prediction of Potential Core Therapeutic Targets. Biomed Res. Int. 2015, 2015, 139580. [Google Scholar] [CrossRef] [Green Version]
- Schubert-Unkmeir, A.; Christodoulides, M. Genome-Based Bacterial Vaccines: Current State and Future Outlook. BioDrugs 2013, 27, 419–430. [Google Scholar] [CrossRef]
- Bai, X.; Findlow, J.; Borrow, R. Recombinant Protein Meningococcal Serogroup B Vaccine Combined with Outer Membrane Vesicles. Expert Opin. Biol. Ther. 2011, 11, 969–985. [Google Scholar] [CrossRef]
- He, Y. Analyses of Brucella Pathogenesis, Host Immunity, and Vaccine Targets Using Systems Biology and Bioinformatics. Front. Cell. Infect. Microbiol. 2012, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Lauer, P.; Rinaudo, C.D.; Soriani, M.; Margarit, I.; Maione, D.; Rosini, R.; Taddei, A.R.; Mora, M.; Rappuoli, R.; Grandi, G.; et al. Genome Analysis Reveals Pili in Group B Streptococcus. Science 2005, 309, 105. [Google Scholar] [CrossRef] [Green Version]
- Nuccitelli, A.; Cozzi, R.; Gourlay, L.J.; Donnarumma, D.; Necchi, F.; Norais, N.; Telford, J.L.; Rappuoli, R.; Bolognesi, M.; Maione, D.; et al. Structure-Based Approach to Rationally Design a Chimeric Protein for an Effective Vaccine against Group B Streptococcus Infections. Proc. Natl. Acad. Sci. USA 2011, 108, 10278–10283. [Google Scholar] [CrossRef] [Green Version]
- Harfouche, C.; Filippini, S.; Gianfaldoni, C.; Ruggiero, P.; Moschioni, M.; Maccari, S.; Pancotto, L.; Arcidiacono, L.; Galletti, B.; Censini, S.; et al. RrgB321, a Fusion Protein of the Three Variants of the Pneumococcal Pilus Backbone RrgB, Is Protective in Vivo and Elicits Opsonic Antibodies. Infect. Immun. 2012, 80, 451–460. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.J.; Norais, N.; Bensi, G.; Liberatori, S.; Capo, S.; Mora, M.; Scarselli, M.; Doro, F.; Ferrari, G.; Garaguso, I.; et al. Characterization and Identification of Vaccine Candidate Proteins through Analysis of the Group A Streptococcus Surface Proteome. Nat. Biotechnol. 2006, 24, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Schrag, A.; Martino, D.; Apter, A.; Ball, J.; Bartolini, E.; Benaroya-Milshtein, N.; Buttiglione, M.; Cardona, F.; Creti, R.; Efstratiou, A.; et al. European Multicentre Tics in Children Studies (EMTICS): Protocol for Two Cohort Studies to Assess Risk Factors for Tic Onset and Exacerbation in Children and Adolescents. Eur. Child Adolesc. Psychiatry 2019, 28, 91–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, F.; Orefici, G. Group A Streptococcal Infections and Tic Disorders in an Italian Pediatric Population. J. Pediatr. 2001, 138, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Romano, A.; Cundari, G.; Ventriglia, F.; Versacci, P.; Orefici, G. Colour Doppler Echocardiography in Children with Group A Streptococcal Infection Related Tic Disorders. Indian J. Med. Res. 2004, 119, 186–190. [Google Scholar] [PubMed]
- Meinke, A.; Henics, T.; Hanner, M.; Minh, D.B.; Nagy, E. Antigenome Technology: A Novel Approach for the Selection of Bacterial Vaccine Candidate Antigens. Vaccine 2005, 23, 2035–2041. [Google Scholar] [CrossRef]
- Nagy, E.; Henics, T.; Gabain, A.v.; Meinke, A. Identification of the ‘Antigenome’—A Novel Tool for Design and Development of Subunit Vaccines Against Bacterial Pathogens. In Genomics Proteomics and Vaccines; Publisher Wiley: Hoboken, NJ, USA, 2004; p. 223. [Google Scholar]
- Chakkyarath, V.; Shanmugam, A.; Natarajan, J. Prioritization of Potential Drug Targets and Antigenic Vaccine Candidates against Klebsiella aerogenes Using the Computational Subtractive Proteome-Driven Approach. J. Proteins Proteom. 2021, 12, 201–211. [Google Scholar] [CrossRef]
- Mora, M.; Telford, J.L. Genome-Based Approaches to Vaccine Development. J. Mol. Med. 2010, 88, 143–147. [Google Scholar] [CrossRef]
- Mayer, R.L.; Impens, F. Immunopeptidomics for Next-Generation Bacterial Vaccine Development. Trends Microbiol. 2021, 29, 1034–1045. [Google Scholar] [CrossRef]
- Ricci, A.D.; Brunner, M.; Ramoa, D.; Carmona, S.J.; Nielsen, M.; Agüero, F. APRANK: Computational Prioritization of Antigenic Proteins and Peptides from Complete Pathogen Proteomes. Front. Immunol. 2021, 12, 702502. [Google Scholar] [CrossRef]
- Palmer, G.H.; Brown, W.C.; Noh, S.M.; Brayton, K.A. Genome-Wide Screening and Identification of Antigens for Rickettsial Vaccine Development. FEMS Immunol. Med. Microbiol. 2012, 64, 115–119. [Google Scholar] [CrossRef]
- Telford, J.L.; Barocchi, M.A.; Margarit, I.; Rappuoli, R.; Grandi, G. Pili in Gram-Positive Pathogens. Nat. Rev. Microbiol. 2006, 4, 509–519. [Google Scholar] [CrossRef]
- Margarit, I.; Rinaudo, C.D.; Galeotti, C.L.; Maione, D.; Ghezzo, C.; Buttazzoni, E.; Rosini, R.; Runci, Y.; Mora, M.; Buccato, S.; et al. Preventing Bacterial Infections with Pilus-Based Vaccines: The Group B Streptococcus Paradigm. J. Infect. Dis. 2009, 199, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Gianfaldoni, C.; Censini, S.; Hilleringmann, M.; Moschioni, M.; Facciotti, C.; Pansegrau, W.; Masignani, V.; Covacci, A.; Rappuoli, R.; Barocchi, M.A.; et al. Streptococcus pneumoniae Pilus Subunits Protect Mice against Lethal Challenge. Infect. Immun. 2007, 75, 1059–1062. [Google Scholar] [CrossRef] [Green Version]
- Grifantini, R.; Frigimelica, E.; Delany, I.; Bartolini, E.; Giovinazzi, S.; Balloni, S.; Agarwal, S.; Galli, G.; Genco, C.; Grandi, G. Characterization of a Novel Neisseria meningitidis Fur and Iron-regulated Operon Required for Protection from Oxidative Stress: Utility of DNA Microarray in the Assignment of the Biological Role of Hypothetical Genes. Mol. Microbiol. 2004, 54, 962–979. [Google Scholar] [CrossRef]
- Kurz, S.; Hübner, C.; Aepinus, C.; Theiss, S.; Guckenberger, M.; Panzner, U.; Weber, J.; Frosch, M.; Dietrich, G. Transcriptome-Based Antigen Identification for Neisseria meningitidis. Vaccine 2003, 21, 768–775. [Google Scholar] [CrossRef]
- Bartolini, E.; Frigimelica, E.; Giovinazzi, S.; Galli, G.; Shaik, Y.; Genco, C.; Welsch, J.A.; Granoff, D.M.; Grandi, G.; Grifantini, R. Role of FNR and FNR-regulated, Sugar Fermentation Genes in Neisseria meningitidis Infection. Mol. Microbiol. 2006, 60, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Holtfreter, S.; Kolata, J.; Stentzel, S.; Bauerfeind, S.; Schmidt, F.; Sundaramoorthy, N.; Bröker, B.M. Omics Approaches for the Study of Adaptive Immunity to Staphylococcus aureus and the Selection of Vaccine Candidates. Proteomes 2016, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Talaat, A.M.; Lyons, R.; Howard, S.T.; Johnston, S.A. The Temporal Expression Profile of Mycobacterium tuberculosis Infection in Mice. Proc. Natl. Acad. Sci. USA 2004, 101, 4602–4607. [Google Scholar] [CrossRef] [Green Version]
- Coppola, M.; Lai, R.P.-J.; Wilkinson, R.J.; Ottenhoff, T.H.M. The in Vivo Transcriptomic Blueprint of Mycobacterium tuberculosis in the Lung. Front. Immunol. 2021, 12, 5212. [Google Scholar] [CrossRef]
- Hinton, J.C.D.; Hautefort, I.; Eriksson, S.; Thompson, A.; Rhen, M. Benefits and Pitfalls of Using Microarrays to Monitor Bacterial Gene Expression during Infection. Curr. Opin. Microbiol. 2004, 7, 277–282. [Google Scholar] [CrossRef]
- Wet, S.C.D.; Denman, S.E.; Sly, L.; McSweeney, C.S. An Improved Method for RNA Extraction from Carcass Samples for Detection of Viable Escherichia coli O157: H7 by Reverse-transcriptase Polymerase Chain Reaction. Lett. Appl. Microbiol. 2008, 47, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.; Marra, M.A. Applications of Next-Generation Sequencing Technologies in Functional Genomics. Genomics 2008, 92, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, K.L. Higher Sensitivity and Multiplexing of Samples Using a Simpler Experimental Protocol. Methods Mol. Biol. 2008, 387, 81–94. [Google Scholar] [PubMed]
- Montemari, A.L.; Marzano, V.; Essa, N.; Mortera, S.L.; Rossitto, M.; Gardini, S.; Selan, L.; Vrenna, G.; Muda, A.O.; Putignani, L.; et al. A Shaving Proteomic Approach to Unveil Surface Proteins Modulation of Multi-Drug Resistant Pseudomonas aeruginosa Strains Isolated From Cystic Fibrosis Patients. Front. Med. 2022, 9, 818669. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Harris, S.R.; Fraser, C.; Quail, M.A.; Burton, J.; Linden, M.V.D.; McGee, L.; Gottberg, A.V.; Song, J.H.; Ko, K.S.; et al. Rapid Pneumococcal Evolution in Response to Clinical Interventions. Science 2011, 331, 430–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.R.; Feil, E.J.; Holden, M.T.G.; Quail, M.A.; Nickerson, E.K.; Chantratita, N.; Gardete, S.; Tavares, A.; Day, N.; Lindsay, J.A.; et al. Evolution of MRSA during Hospital Transmission and Intercontinental Spread. Science 2010, 327, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.M.; Hoffmann, S.; Darfeuille, F.; Reignier, J.; Findeiß, S.; Sittka, A.; Chabas, S.; Reiche, K.; Hackermüller, J.; Reinhardt, R.; et al. The Primary Transcriptome of the Major Human Pathogen Helicobacter pylori. Nature 2010, 464, 250–255. [Google Scholar] [CrossRef]
- Kuhlmann, L.; Cummins, E.; Samudio, I.; Kislinger, T. Cell-Surface Proteomics for the Identification of Novel Therapeutic Targets in Cancer. Expert Rev. Proteom. 2018, 15, 259–275. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Barbieri, G.; Buroni, S.; Scarselli, M.; Pizza, M.; Rappuoli, R.; Riccardi, G. Vaccines to Overcome Antibiotic Resistance: The Challenge of Burkholderia cenocepacia. Trends Microbiol. 2020, 28, 315–326. [Google Scholar] [CrossRef]
- Sousa, A.C.; Neiva, H.P.; Izquierdo, M.; Alves, A.R.; Duarte-Mendes, P.; Ramalho, A.G.; Marques, M.C.; Marinho, D.A. Concurrent Training Intensities: A Practical Approach for Program Design. Strength Cond. J. 2020, 42, 38–44. [Google Scholar] [CrossRef]
- Sharma, A.; Arya, D.K.; Sagar, V.; Bergmann, R.; Chhatwal, G.S.; Johri, A.K. Identification of Potential Universal Vaccine Candidates against Group A Streptococcus by Using High Throughput in Silico and Proteomics Approach. J. Proteome Res. 2013, 12, 336–346. [Google Scholar] [CrossRef]
- Nilsson, J.; Hansson, G.K. Vaccination Strategies and Immune Modulation of Atherosclerosis. Circ. Res. 2020, 126, 1281–1296. [Google Scholar] [CrossRef]
- Zielke, R.A.; Wierzbicki, I.H.; Baarda, B.I.; Gafken, P.R.; Soge, O.O.; Holmes, K.K.; Jerse, A.E.; Unemo, M.; Sikora, A.E. Proteomics-Driven Antigen Discovery for Development of Vaccines against Gonorrhea. Mol. Cell. Proteom. 2016, 15, 2338–2355. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.W.; Moriel, D.G.; Phan, M.-D.; Schulz, B.L.; Kidd, T.J.; Beatson, S.A.; Schembri, M.A. ‘Omic’Approaches to Study Uropathogenic Escherichia coli Virulence. Trends Microbiol. 2017, 25, 729–740. [Google Scholar] [CrossRef]
- Bagnoli, F.; Baudner, B.; Mishra, R.P.N.; Bartolini, E.; Fiaschi, L.; Mariotti, P.; Nardi-Dei, V.; Boucher, P.; Rappuoli, R. Designing the next Generation of Vaccines for Global Public Health. OMICS A J. Integr. Biol. 2011, 15, 545–566. [Google Scholar] [CrossRef]
- Domon, B.; Aebersold, R. Mass Spectrometry and Protein Analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Mann, M.; Hendrickson, R.C.; Pandey, A. Analysis of Proteins and Proteomes by Mass Spectrometry. Annu. Rev. Biochem. 2001, 70, 437–473. [Google Scholar] [CrossRef]
- Karas, M.; Hillenkamp, F. Laser Desorption Ionization of Proteins with Molecular Masses Exceeding 10,000 Daltons. Anal. Chem. 1988, 60, 2299–2301. [Google Scholar] [CrossRef]
- Medzihradszky, K.F.; Campbell, J.M.; Baldwin, M.A.; Falick, A.M.; Juhasz, P.; Vestal, M.L.; Burlingame, A.L. The Characteristics of Peptide Collision-Induced Dissociation Using a High-Performance MALDI-TOF/TOF Tandem Mass Spectrometer. Anal. Chem. 2000, 72, 552–558. [Google Scholar] [CrossRef]
- Chandramouli, K.; Qian, P.Y. Proteomics: Challenges, Techniques and Possibilities to Overcome Biological Sample Complexity. Hum. Genom. Proteom. HGP 2009, 2009, 239204. [Google Scholar] [CrossRef]
- Chen, E.I.; Hewel, J.; Felding-Habermann, B.; Yates, J.R. Large Scale Protein Profiling by Combination of Protein Fractionation and Multidimensional Protein Identification Technology (MudPIT). Mol. Cell. Proteom. 2006, 5, 53–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Wang, H.; Chen, L. Comparative Secretomics Reveals Novel Virulence-Associated Factors of Vibrio parahaemolyticus. Front. Microbiol. 2015, 6, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravipaty, S.; Reilly, J.P. Comprehensive Characterization of Methicillin-Resistant Staphylococcus aureus Subsp. aureus COL Secretome by Two-Dimensional Liquid Chromatography and Mass Spectrometry. Mol. Cell. Proteom. 2010, 9, 1898–1919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Thompson, K.M.; Francis, M.S. Environmental Regulation of Yersinia Pathophysiology. Front. Cell. Infect. Microbiol. 2016, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Torres-Sangiao, E.; Giddey, A.D.; Rodriguez, C.L.; Tang, Z.; Liu, X.; Soares, N.C. Proteomic Approaches to Unravel Mechanisms of Antibiotic Resistance and Immune Evasion of Bacterial Pathogens. Front. Med. 2022, 9, 850374. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Q.; Hu, M.; Yu, K.; Fu, J.; Zhou, F.; Liu, X. Proteomic Analyses of Intracellular Salmonella enterica Serovar Typhimurium Reveal Extensive Bacterial Adaptations to Infected Host Epithelial Cells. Infect. Immun. 2015, 83, 2897–2906. [Google Scholar] [CrossRef] [Green Version]
- Noster, J.; Chao, T.-C.; Sander, N.; Schulte, M.; Reuter, T.; Hansmeier, N.; Hensel, M. Proteomics of Intracellular Salmonella enterica Reveals Roles of Salmonella Pathogenicity Island 2 in Metabolism and Antioxidant Defense. PLoS Pathog. 2019, 15, e1007741. [Google Scholar] [CrossRef] [Green Version]
- Pieper, R.; Fisher, C.R.; Suh, M.-J.; Huang, S.-T.; Parmar, P.; Payne, S.M. Analysis of the Proteome of Intracellular Shigella flexneri Reveals Pathways Important for Intracellular Growth. Infect. Immun. 2013, 81, 4635–4648. [Google Scholar] [CrossRef] [Green Version]
- Birk, M.S.; Ahmed-Begrich, R.; Tran, S.; Elsholz, A.K.W.; Frese, C.K.; Charpentier, E. Time-Resolved Proteome Analysis of Listeria Monocytogenes during Infection Reveals the Role of the AAA+ Chaperone ClpC for Host Cell Adaptation. Msystems 2021, 6, e00215-21. [Google Scholar] [CrossRef]
- Lamberti, Y.; Cafiero, J.H.; Surmann, K.; Valdez, H.; Holubova, J.; Večerek, B.; Sebo, P.; Schmidt, F.; Völker, U.; Rodriguez, M.E. Proteome Analysis of Bordetella pertussis Isolated from Human Macrophages. J. Proteom. 2016, 136, 55–67. [Google Scholar] [CrossRef]
- Lamontagne, J.; Forest, A.; Marazzo, E.; Denis, F.; Butler, H.; Michaud, J.-F.; Boucher, L.; Pedro, I.; Villeneuve, A.; Sitnikov, D.; et al. Intracellular Adaptation of Brucella abortus. J. Proteome Res. 2009, 8, 1594–1609. [Google Scholar] [CrossRef] [Green Version]
- Chandonia, J.-M.; Brenner, S.E. The Impact of Structural Genomics: Expectations and Outcomes. Science 2006, 311, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Dormitzer, P.R.; Ulmer, J.B.; Rappuoli, R. Structure-Based Antigen Design: A Strategy for next Generation Vaccines. Trends Biotechnol. 2008, 26, 659–667. [Google Scholar] [CrossRef]
- Cozzi, R.; Scarselli, M.; Ferlenghi, I. Structural Vaccinology: A Three-Dimensional View for Vaccine Development. Curr. Top. Med. Chem. 2013, 13, 2629–2637. [Google Scholar]
- Masignani, V.; Comanducci, M.; Giuliani, M.M.; Bambini, S.; Adu-Bobie, J.; Aricò, B.; Brunelli, B.; Pieri, A.; Santini, L.; Savino, S.; et al. Vaccination against Neisseria meningitidis Using Three Variants of the Lipoprotein GNA1870. J. Exp. Med. 2003, 197, 789–799. [Google Scholar] [CrossRef]
- Scarselli, M.; Aricò, B.; Brunelli, B.; Savino, S.; Marcello, F.D.; Palumbo, E.; Veggi, D.; Ciucchi, L.; Cartocci, E.; Bottomley, M.J.; et al. Rational Design of a Meningococcal Antigen Inducing Broad Protective Immunity. Sci. Transl. Med. 2011, 3, 91ra62. [Google Scholar] [CrossRef]
- Jiang, H.-Q.; Hoiseth, S.K.; Harris, S.L.; McNeil, L.K.; Zhu, D.; Tan, C.; Scott, A.A.; Alexander, K.; Mason, K.; Miller, L.; et al. Broad Vaccine Coverage Predicted for a Bivalent Recombinant Factor H Binding Protein Based Vaccine to Prevent Serogroup B Meningococcal Disease. Vaccine 2010, 28, 6086–6093. [Google Scholar] [CrossRef]
- Fletcher, L.D.; Bernfield, L.; Barniak, V.; Farley, J.E.; Howell, A.; Knauf, M.; Ooi, P.; Smith, R.P.; Weise, P.; Wetherell, M.; et al. Vaccine Potential of the Neisseria meningitidis 2086 Lipoprotein. Infect. Immun. 2004, 72, 2088–2100. [Google Scholar] [CrossRef] [Green Version]
- Mascioni, A.; Bentley, B.E.; Camarda, R.; Dilts, D.A.; Fink, P.; Gusarova, V.; Hoiseth, S.K.; Jacob, J.; Lin, S.L.; Malakian, K.; et al. Structural Basis for the Immunogenic Properties of the Meningococcal Vaccine Candidate LP2086. J. Biol. Chem. 2009, 284, 8738–8746. [Google Scholar] [CrossRef] [Green Version]
- Cantini, F.; Veggi, D.; Dragonetti, S.; Savino, S.; Scarselli, M.; Romagnoli, G.; Pizza, M.; Banci, L.; Rappuoli, R. Solution Structure of the Factor H-Binding Protein, a Survival Factor and Protective Antigen of Neisseria meningitidis. J. Biol. Chem. 2009, 284, 9022–9026. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.C.; Prosser, B.E.; Caesar, J.J.E.; Kugelberg, E.; Li, S.; Zhang, Q.; Quoraishi, S.; Lovett, J.E.; Deane, J.E.; Sim, R.B.; et al. Neisseria meningitidis Recruits Factor H Using Protein Mimicry of Host Carbohydrates. Nature 2009, 458, 890–893. [Google Scholar] [CrossRef]
- Giuntini, S.; Reason, D.C.; Granoff, D.M. Complement-Mediated Bactericidal Activity of Anti-Factor H Binding Protein Monoclonal Antibodies against the Meningococcus Relies upon Blocking Factor H Binding. Infect. Immun. 2011, 79, 3751–3759. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Khatoon, N.; Mishra, A.; Verma, V.K.; Prajapati, V.K. Structural Vaccinology Approach to Investigate the Virulent and Secretory Proteins of Bacillus anthracis for Devising Anthrax Next-Generation Vaccine. J. Biomol. Struct. Dyn. 2020, 38, 4895–4905. [Google Scholar] [CrossRef]
- Beckham, K.S.H.; Ritter, C.; Chojnowski, G.; Ziemianowicz, D.S.; Mullapudi, E.; Rettel, M.; Savitski, M.M.; Mortensen, S.A.; Kosinski, J.; Wilmanns, M. Structure of the Mycobacterial ESX-5 Type VII Secretion System Pore Complex. Sci. Adv. 2021, 7, eabg9923. [Google Scholar] [CrossRef]
- Hotz, R.L. Scientists Create Synthetic Organism. Wall Str. J. 2010, 21. Available online: https://library.wur.nl/WebQuery/file/cogem/cogem_t4c59c64b_001.pdf (accessed on 12 December 2022).
- Venter, J.C.; Glass, J.I.; Hutchison III, C.A.; Vashee, S. Synthetic Chromosomes, Genomes, Viruses, and Cells. Cell 2022, 185, 2708–2724. [Google Scholar] [CrossRef] [PubMed]
- Nigam, A.; Almabruk, K.H.; Saxena, A.; Yang, J.; Mukherjee, U.; Kaur, H.; Kohli, P.; Kumari, R.; Singh, P.; Zakharov, L.N.; et al. Modification of Rifamycin Polyketide Backbone Leads to Improved Drug Activity against Rifampicin-Resistant Mycobacterium tuberculosis. J. Biol. Chem. 2014, 289, 21142–21152. [Google Scholar] [CrossRef] [Green Version]
- Saxena, A.; Mukherjee, U.; Kumari, R.; Singh, P.; Lal, R. Synthetic Biology in Action: Developing a Drug against MDR-TB. Indian J. Microbiol. 2014, 54, 369–375. [Google Scholar] [CrossRef]



| Tool Category | Bioinformatics Tool | Description/Web Site |
|---|---|---|
| General tools | Basic Local Alignment Search Tool (BLAST) | Sequence similarity database http://www.ncbi.nlm.nih.gov/BLAST/ (accessed on 2 September 2022) |
| Protein Subcellular Localization Predictor Tool for bacteria (PSORTb) | Bacterial protein subcellular localization (SCL) predictor http://www.psort.org/psortb/ (accessed on 2 September 2022) | |
| Signal peptides (SignalP) | Prediction of signal peptides http://www.cbs.dtu.dk/services/SignalP/ (accessed on 2 September 2022) | |
| Expert Protein Analysis System (EXPASY) | Translate nucleotide to protein http://www.expasy.org/ (accessed on 2 September 2022) | |
| Protein Data Bank (PDB) | Structure of protein http://www.pdb.org/pdb/home/home.do (accessed on 2 September 2022) | |
| Protein Variability Server (PVS) | Sequence variability calculator http://imed.med.ucm.es/PVS/ (accessed on 2 September 2022) | |
| Allergenic Prediction (AlgPred) | Predicting allergenic proteins and allergenic regions in a protein http://www.imtech.res.in/raghava/algpred/ (accessed on 2 September 2022) | |
| T cell epitope prediction | Epitopes Major Histocompatibility Complex (EPIMHC) | MHC-binding peptides and T cell epitopes http://bio.dfci.harvard.edu/epimhc/ (accessed on 2 September 2022) |
| Cytotoxic T lymphocytes Predication (CTL PRED) | Direct approach for CTL epitope prediction essential for designing subunit vaccines http://www.imtech.res.in/raghava/ctlpred/ (accessed on 2 September 2022) | |
| EpiVax | Epimatrix algorithm http://www.epivax.com/ (accessed on 2 September 2022) | |
| Kernel-based Inter-allele peptide binding prediction SyStem (KISS) | SVM-based method KISS http://cbio.ensmp.fr/kiss/ (accessed on 2 September 2022) | |
| Protein Predication (ProPred) | Web-based graphical prediction tool for MHC class II binding sites in antigenic protein sequences http://www.imtech.res.in/raghava/propred/ (accessed on 2 September 2022) | |
| IMTECH | Quantitative matrix approach http://www.imtech.res.in/raghava/mhc (accessed on 2 September 2022) | |
| Promiscuous EPitope-based VACcine (PEPVAC) | Prediction of promiscuous epitopes http://immunax.dfci.harvard.edu/PEPVAC/ (accessed on 2 September 2022) | |
| RANKPEP | Predicts peptide binders to MHCI and MHCII http://bio.dfci.harvard.edu/Tools/rankpep.html (accessed on 2 September 2022) | |
| SYFPEITHI | Database for searching and T cell epitope prediction http://www.syfpeithi.de/ (accessed on 2 September 2022) | |
| Immune Epitope Database (IEDB) | Identification of novel B and T cell epitopes in proteins of interest http://www.immuneepitope.org/ (accessed on 2 September 2022) | |
| NetCTL 1.2 Server | Verified Prediction of CTL epitopes in protein sequences http://www.cbs.dtu.dk/services/NetCTL/ (accessed on 2 September 2022) | |
| Major Histocompatibility Complex Predication (MHCPred) | A quantitative T cell epitope prediction server http://www.ddg-pharmfac.net/mhcpred/MHCPred/ (accessed on 2 September 2022) | |
| NetMHC 3.0 | Predicts peptide binding to variety of different HLA alleles http://www.cbs.dtu.dk/services/NetMHC/ (accessed on 2 September 2022) | |
| Support Vector machine-based method form MHC (SVMHC) | Prediction of MHC-binding peptides https://abi.inf.uni-tuebingen.de/Services/SVMHC (accessed on 2 September 2022) | |
| EpiJen | Multistep algorithm for T cell epitope prediction http://www.ddg-pharmfac.net/epijen/EpiJen/ (accessed on 2 September 2022) | |
| Major Histocompatibility Complex Binding and Non-binding peptides (MHCBN) | Information about MHC binders, MHC non-binder, TAP binders, TAP non-binders, and T cell epitopes http://www.imtech.res.in/raghava/mhcbn/ (accessed on 2 September 2022) | |
| B cell epitope tools | B-cell epitopes predication 2.0 (BepiPred 2.0) | Continuous B cell epitope identification http://www.cbs.dtu.dk/services/BepiPred/ (accessed on 2 September 2022) |
| B-cell predication (Bcepred) | Continuous B cell epitope identification http://crdd.osdd.net/raghava/bcepred/ (accessed on 2 September 2022) | |
| B-cell Epitope prediction using Support vector machine Tool (BEST) | Continuous B cell epitope identification http://biomine.cs.vcu.edu/datasets/BEST/ (accessed on 2 September 2022) | |
| Continuous B-cell Epitopes Pro (COBEpro) | Predictions on short peptide fragments http://scratch.proteomics.ics.uci.edu/ (accessed on 2 September 2022) | |
| Conformational B-cell Epitope predication (CBTOPE) | Prediction of conformational B cell epitope http://crdd.osdd.net/raghava/cbtope/submit.php (accessed on 2 September 2022) | |
| DiscoTope 2.0 | Discontinuous B cell epitopes prediction http://www.cbs.dtu.dk/services/DiscoTope/ (accessed on 2 September 2022) | |
| Peptide epitope (Pepitope) | Continuous and discontinuous B cell epitopes prediction http://pepitope.tau.ac.il/ (accessed on 2 September 2022) | |
| Epitopia | Continuous and discontinuous B cell epitope prediction http://epitopia.tau.ac.il/ (accessed on 2 September 2022) | |
| DiscoTope 1.2 Server | Uses 3D structure of protein to forecast discontinuous B cell epitopes http://www.cbs.dtu.dk/services/DiscoTope/ (accessed on 2 September 2022) | |
| Linear B-cell epitopes (LBtope) | Accurate method for linear B cell epitopes (https://webs.iiitd.edu.in/raghava/lbtope/ (accessed on 2 September 2022) | |
| Ellipsoid and Protrusion (ElliPro) | Method used for continuous and discontinuous B cell epitopes prediction http://www.pepitope.tau.ac.il/ (accessed on 2 September 2022) | |
| Artificial neural network-based B-cell epitope prediction (ABCpred) | Predicting linear B cell epitopes (https://webs.iiitd.edu.in/raghava/abcpred/ (accessed on 2 September 2022) | |
| Proteasomal cleavage (Pcleavage) | Proteasomal cleavage site identification (https://webs.iiitd.edu.in/raghava/pcleavage/ (accessed on 2 September 2022) | |
| A database for B-Cells Epitopes (BCIPEP) | B cell epitope validation http://www.imtech.res.in/raghava/bcipep/index.html (accessed on 2 September 2022) |
| Bacteria | Disease | Techniques | Vaccine Status |
|---|---|---|---|
| C. pneumoniae | Pneumonia, meningitis, middle ear infections | Reverse vaccinology Proteomics Pan-genomic | Discovery/pre-clinical |
| B. anthracis | Anthrax | Reverse vaccinology CGH microarray Microarray Proteomics Immunoproteomics Antigen prioritization | Discovery/pre-clinical |
| R. prowazekii | Epidemic typhus, also called louse-borne typhus | Reverse vaccinology Proteomics Pan-genomic | Discovery/pre-clinical |
| M. tuberculosis | Tuberculosis | Reverse vaccinology Structural genomics/vaccinology Synthetic genomics Antigen prioritization Pan-genomic | Discovery/pre-clinical |
| H. pylori | Ulcer, atrophic gastritis, adenocarcinoma, lymphoma | Reverse vaccinology Immunoproteomics Pan-genomic | Discovery/pre-clinical |
| P. gingivalis | Periodontitis | Reverse vaccinology | Discovery/pre-clinical |
| N.meningitidis serogroup B | Bacterial meningitis and septicemia | Reverse vaccinology Microarray Proteomics | Phase II clinical trials |
| Brucella | Brucellosis | Proteomics | Discovery/pre-clinical |
| L. monocytogenes | Invasive foodborne infections. | Proteomics | Phase 1 clinical trials |
| Bacteria | Developed Vaccine/Antigen Identified by Pan-Genomics | References |
|---|---|---|
| N. meningitidis | 4CMenB (Bexsero®) vaccine | [141] |
| C. pneumoniae | LcrE antigen | [102] |
| Brucella | RV web-based vaccine design program (VaxiJen), that identified O-sialoglycoprotein endopeptidase; 32 OMPs (Omp2b, Omp25, Omp31-1, TonB, adhesins, and adhesin-like proteins (FlgE and FlgK) | [142] |
| S. agalactiae | GBS322 (SAG0032, Sip protein), GBS67 (SAG1408), GBS80 | [95] |
| (SAG0645), GBS104 (SAG0649) proteins | [143] | |
| HMW pilus-based vaccine | [144] | |
| S. pneumoniae | Sp36, Sp46, Sp91, Sp101, and Sp128/130 protective antigens (cell wall anchor) | [89] |
| RrgB321 (fusion protein of three RrgB variants) | [145] | |
| S. pyogenes (GAS) | Cpa, MI_128, and MI_130 (Recombinant pilus protein) | [96] |
| Protective antigen Spy0416 | [146] | |
| L. pneumophila | Q5ZVG4, Q5ZRZ1, Q5ZWE6, Q5ZT09, and Q5ZUZ8 | [134] |
| M.tuberculosis | PE/PPE, plcC | [124] |
| ESAT-6 antigen and diacylglycerol acyltransferase | [129] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.A.; Amin, A.; Farid, A.; Ullah, A.; Waris, A.; Shinwari, K.; Hussain, Y.; Alsharif, K.F.; Alzahrani, K.J.; Khan, H. Recent Advances in Genomics-Based Approaches for the Development of Intracellular Bacterial Pathogen Vaccines. Pharmaceutics 2023, 15, 152. https://doi.org/10.3390/pharmaceutics15010152
Khan MA, Amin A, Farid A, Ullah A, Waris A, Shinwari K, Hussain Y, Alsharif KF, Alzahrani KJ, Khan H. Recent Advances in Genomics-Based Approaches for the Development of Intracellular Bacterial Pathogen Vaccines. Pharmaceutics. 2023; 15(1):152. https://doi.org/10.3390/pharmaceutics15010152
Chicago/Turabian StyleKhan, Muhammad Ajmal, Aftab Amin, Awais Farid, Amin Ullah, Abdul Waris, Khyber Shinwari, Yaseen Hussain, Khalaf F. Alsharif, Khalid J. Alzahrani, and Haroon Khan. 2023. "Recent Advances in Genomics-Based Approaches for the Development of Intracellular Bacterial Pathogen Vaccines" Pharmaceutics 15, no. 1: 152. https://doi.org/10.3390/pharmaceutics15010152