
Current Strategies to Enhance Delivery of Drugs across the Blood–Brain Barrier



 ,
,  and
and
Abstract
:1. Introduction
2. General Background on the Blood–Brain Barrier
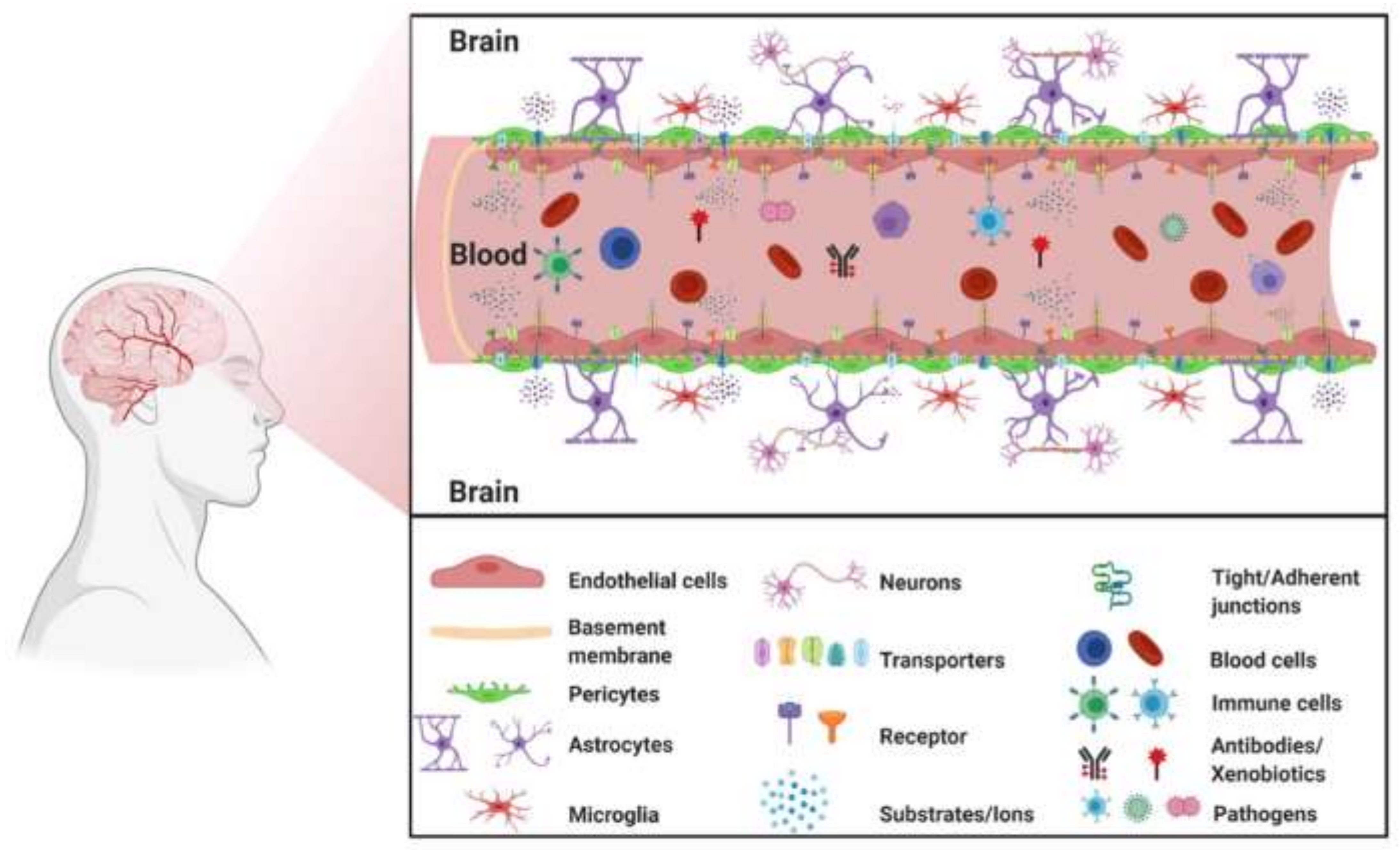
2.1. The Blood–Brain Barrier in a Healthy Brain
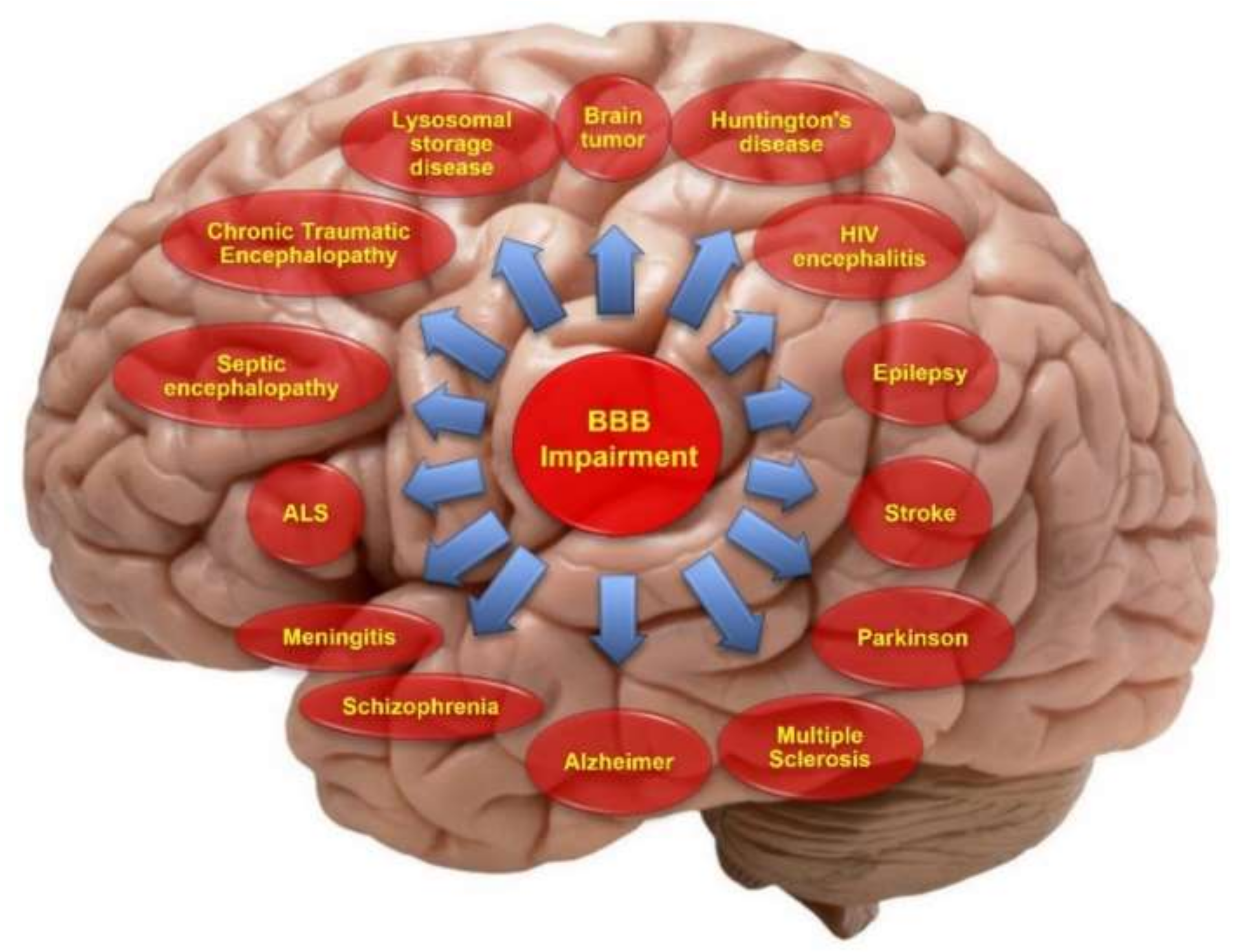
2.2. Blood–Brain Barrier Disruption in Certain Pathological Conditions
2.3. Blood–Brain Tumor Barrier
2.4. Approaches to Inhibit ABC Transporters
3. Advances in Drug Delivery across the Blood–Brain Barrier
3.1. Invasive Approaches
3.1.1. Blood–Brain Barrier Transient Disruption
3.1.2. Intracerebroventricular and Intrathecal Infusion
3.2. Non-Invasive Approaches
3.2.1. Chemical Modification of Drugs
3.2.2. Virus-Mediated Distribution across the Blood–Brain Barrier
3.2.3. Exosome-Mediated Transport across the Blood–Brain Barrier
3.2.4. Intranasal Route of Delivery
3.2.5. Modulating Blood–Brain Barrier Permeability
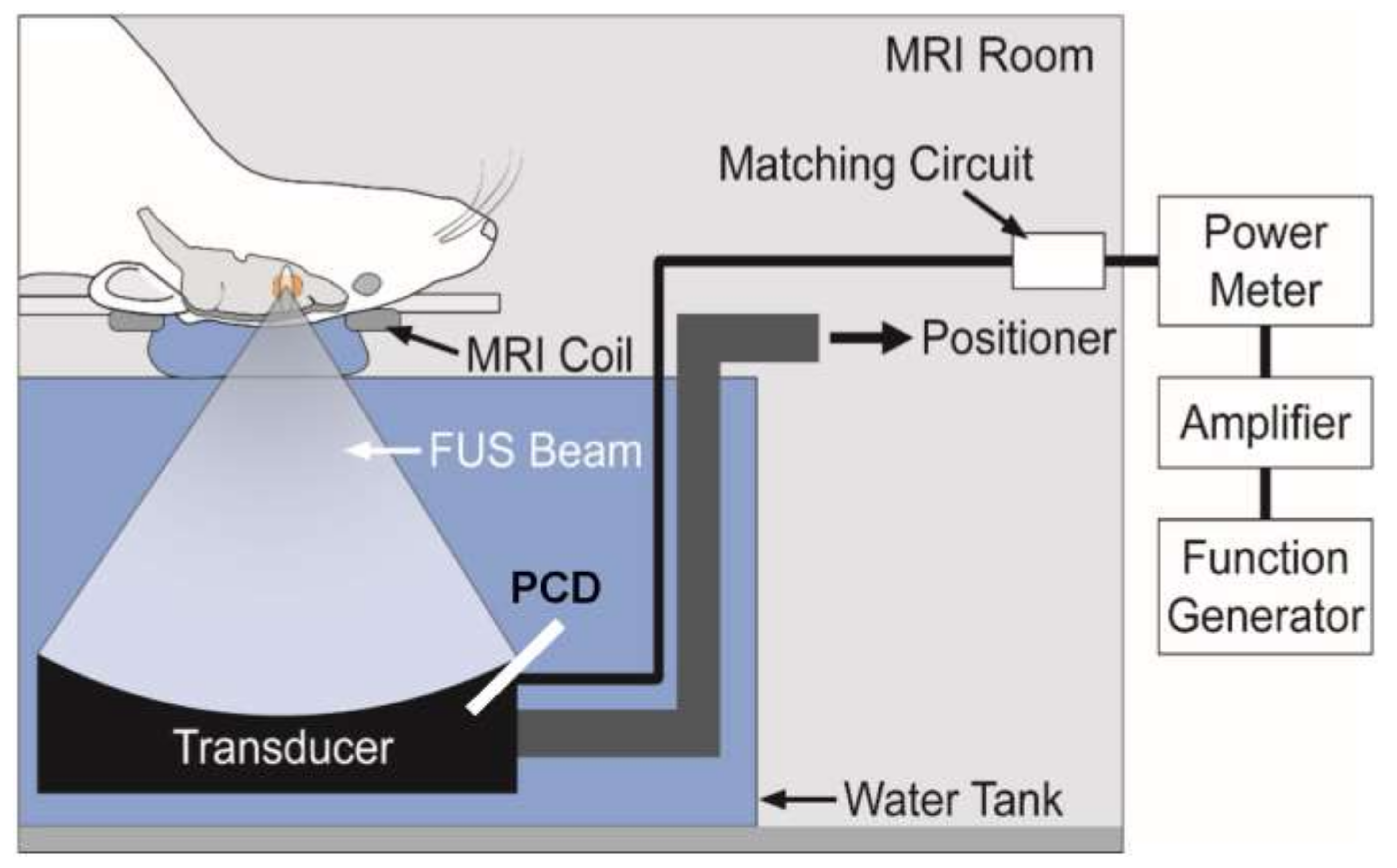
3.2.6. Focused Ultrasound for Brain Diseases
3.2.7. Nanoparticles for Brain Imaging/Diagnostics
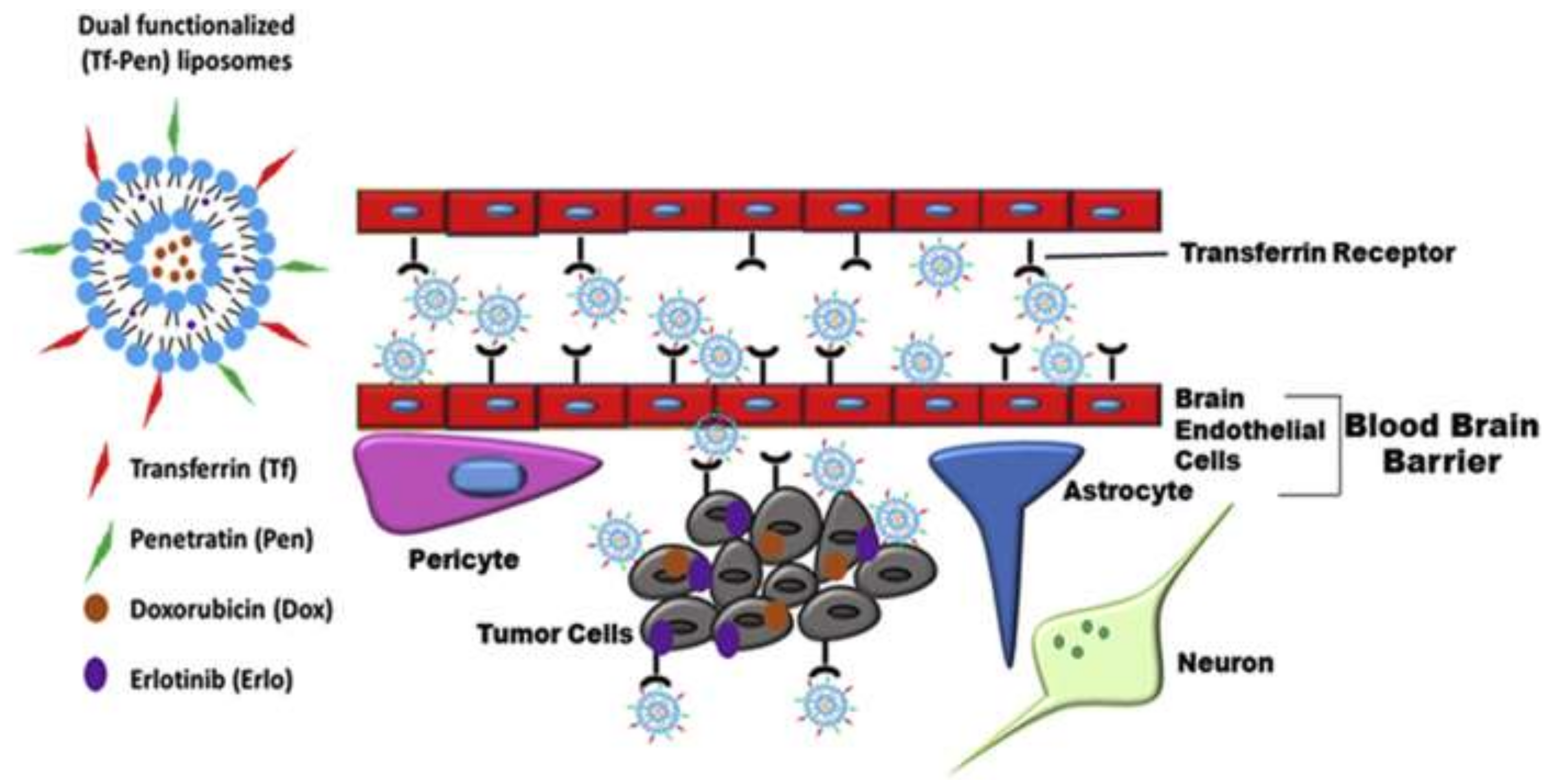
3.2.8. Liposome-Based Strategies for Effective Drug Delivery across the Blood–Brain Barrier
3.2.9. Solid-Lipid Nanoparticles
3.3. Mechanisms by Which Nanosystems Pass through the BBB
4. In Vitro and In Vivo Models of BBB for Assessing Drug Delivery
5. Clinical Trials and Products Available on the Market
6. Perspectives and Future Research Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asaduzzaman, S.; Chakma, R.; Rehana, H.; Raihan, M. Regulatory Gene Network Pathway among Brain Cancer and Associated Disease: A Computational Analysis. Biointerface Res. Appl. Chem. 2021, 11, 12973–12984. [Google Scholar] [CrossRef]
- Lingineni, K.; Belekar, V.; Tangadpalliwar, S.R.; Garg, P. The role of multidrug resistance protein (MRP-1) as an active efflux transporter on blood-brain barrier (BBB) permeability. Mol. Divers. 2017, 21, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Goyal, D.; Shuaib, S.; Mann, S.; Goyal, B. Rationally Designed Peptides and Peptidomimetics as Inhibitors of Amyloid-beta (A beta) Aggregation: Potential Therapeutics of Alzheimer’s Disease. Acs Comb. Sci. 2017, 19, 55–80. [Google Scholar] [CrossRef]
- Pardridge, W.M. Transport of small molecules through the blood-brain-barrier—biology and methodology. Adv. Drug Deliv. Rev. 1995, 15, 5–36. [Google Scholar] [CrossRef]
- McCaffrey, G.; Davis, T.P. Physiology and pathophysiology of the blood-brain barrier: P-glycoprotein and occludin trafficking as therapeutic targets to optimize central nervous system drug delivery. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 2012, 60, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A. From blood-brain barrier to blood-brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2016, 15, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Ishola, A.A.; Oyinloye, B.E.; Ajiboye, B.O.; Kappo, A.P. Molecular Docking Studies of Flavonoids from Andrographis paniculata as Potential Acetylcholinesterase, Butyrylcholinesterase and Monoamine Oxidase Inhibitors Towards the Treatment of Neurodegenerative Diseases. Biointerface Res. Appl. Chem. 2021, 11, 9871–9879. [Google Scholar] [CrossRef]
- Neuwelt, E.; Abbott, N.; Abrey, L.; Banks, W.A.; Blakley, B.; Davis, T.; Engelhardt, B.; Grammas, P.; Nedergaard, M.; Nutt, J.; et al. Strategies to advance translational research into brain barriers. Lancet Neurol. 2008, 7, 84–96. [Google Scholar] [CrossRef]
- Circulation and the Central Nervous System Anatomy and Physiology I. Available online: https://courses.lumenlearning.com/ap1/chapter/circulation-and-the-central-nervous-system/ (accessed on 3 December 2021).
- De Luca, C.; Colangelo, A.M.; Alberghina, L.; Papa, M. Neuro-Immune Hemostasis: Homeostasis and Diseases in the Central Nervous System. Front. Cell. Neurosci. 2018, 12, 459. [Google Scholar] [CrossRef] [Green Version]
- Profaci, C.P.; Munji, R.N.; Pulido, R.S.; Daneman, R. The blood-brain barrier in health and disease: Important unanswered questions. J. Exp. Med. 2020, 217, e20190062. [Google Scholar] [CrossRef]
- Abbott, N.J.; Ronnback, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Rihani, S.B.; Darakjian, L.I.; Deodhar, M.; Dow, P.; Turgeon, J.; Michaud, V. Disease-Induced Modulation of Drug Transporters at the Blood–Brain Barrier Level. Int. J. Mol. Sci. 2021, 22, 3742. [Google Scholar] [CrossRef]
- Pehlivan, S.B. Nanotechnology-based drug delivery systems for targeting, imaging and diagnosis of neurodegenerative diseases. Pharm. Res. 2013, 30, 2499–2511. [Google Scholar] [CrossRef] [PubMed]
- Komarova, Y.A.; Kruse, K.; Mehta, D.; Malik, A.B. Protein Interactions at Endothelial Junctions and Signaling Mechanisms Regulating Endothelial Permeability. Circ. Res. 2017, 120, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecuyer, M.A.; Saint-Laurent, O.; Bourbonniere, L.; Larouche, S.; Larochelle, C.; Michel, L.; Charabati, M.; Abadier, M.; Zandee, S.; Jahromi, N.H.; et al. Dual role of ALCAM in neuroinflammation and blood-brain barrier homeostasis. Proc. Natl. Acad. Sci. USA 2017, 114, E524–E533. [Google Scholar] [CrossRef] [Green Version]
- Banks, W.A. Characteristics of compounds that cross the blood-brain barrier. BMC Neurol. 2009, 9 (Suppl. 1), S3. [Google Scholar] [CrossRef] [Green Version]
- Kadry, H.; Noorani, B.; Cucullo, L. A blood–brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS 2020, 17, 69. [Google Scholar] [CrossRef]
- Koepsell, H. Glucose transporters in brain in health and disease. Pflug. Arch. Eur. J. Physiol. 2020, 472, 1299–1343. [Google Scholar] [CrossRef]
- Ballabh, P.; Braun, A.; Nedergaard, M. The blood-brain barrier: An overview—Structure, regulation, and clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [Google Scholar] [CrossRef]
- Mager, I.; Meyer, A.H.; Li, J.H.; Lenter, M.; Hildebrandt, T.; Leparc, G.; Wood, M.J.A. Targeting blood-brain-barrier transcytosis—perspectives for drug delivery. Neuropharmacology 2017, 120, 4–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Ferdous, K.S.; Ahmed, M. Emerging Promise of Nanoparticle-Based Treatment for Parkinson’s disease. Biointerface Res. Appl. Chem. 2020, 10, 7135–7151. [Google Scholar] [CrossRef]
- Haghighatseir, N.; Ashrafi, H.; Rafiei, P.; Azadi, A. Dexamethasone ameliorates Alzheimer’s pathological condition via inhibiting Nf-kappa B and mTOR signaling pathways. Biointerface Res. Appl. Chem. 2020, 10, 5792–5796. [Google Scholar] [CrossRef]
- Archie, S.R.; Al Shoyaib, A.; Cucullo, L. Blood-Brain Barrier Dysfunction in CNS Disorders and Putative Therapeutic Targets: An Overview. Pharmaceutics 2021, 13, 1779. [Google Scholar] [CrossRef]
- Shiraishi, K.; Wang, Z.J.; Kokuryo, D.; Aoki, I.; Yokoyama, M. A polymeric micelle magnetic resonance imaging (MRI) contrast agent reveals blood-brain barrier (BBB) permeability for macromolecules in cerebral ischemia-reperfusion injury. J. Control. Release 2017, 253, 165–171. [Google Scholar] [CrossRef]
- Di Pardo, A.; Amico, E.; Scalabri, F.; Pepe, G.; Castaldo, S.; Elifani, F.; Capocci, L.; De Sanctis, C.; Comerci, L.; Pompeo, F.; et al. Impairment of blood-brain barrier is an early event in R6/2 mouse model of Huntington Disease. Sci. Rep. 2017, 7, 41316. [Google Scholar] [CrossRef]
- Gao, X.P.; Kouklis, P.; Xu, N.; Minshall, R.D.; Sandoval, R.; Vogel, S.M.; Malik, A.B. Reversibility of increased microvessel permeability in response to VE-cadherin disassembly. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1218–L1225. [Google Scholar] [CrossRef]
- Rosenberg, G.A. Blood-Brain Barrier Permeability in Aging and Alzheimer’s Disease. J. Prev. Alzheimers Dis. 2014, 1, 138–139. [Google Scholar] [CrossRef]
- Rasouli, R.; Zaaeri, F.; Rajabi, A.B.; Darbandi-Azar, A.; Faridi-Majidi, R.; Ardestani, M.S. Tc-99m-Anionic Linear Globular Dendrimer-G2-Phenylalanine Conjugate: Novel Brain Tumor SPECT Imaging. Biointerface Res. Appl. Chem. 2021, 11, 11244–11255. [Google Scholar] [CrossRef]
- van Tellingen, O.; Yetkin-Arik, B.; de Gooijer, M.C.; Wesseling, P.; Wurdinger, T.; de Vries, H.E. Overcoming the blood-brain tumor barrier for effective glioblastoma treatment. Drug Resist. Updates 2015, 19, 1–12. [Google Scholar] [CrossRef]
- Sharma, H.S.; Muresanu, D.F.; Castellani, R.J.; Nozari, A.; Lafuente, J.V.; Tian, Z.R.; Sahib, S.; Bryukhovetskiy, I.; Bryukhovetskiy, A.; Buzoianu, A.D.; et al. Chapter One—Pathophysiology of blood-brain barrier in brain tumor. Novel therapeutic advances using nanomedicine. In International Review of Neurobiology; Bryukhovetskiy, I., Sharma, A., Zhang, Z., Sharma, H.S., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 151, pp. 1–66. [Google Scholar]
- Wanjale, M.V.; Kumar, G.S.V. Peptides as a therapeutic avenue for nanocarrier-aided targeting of glioma. Expert Opin. Drug Deliv. 2017, 14, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Kooij, G.; van Horssen, J.; Bandaru, V.V.; Haughey, N.J.; de Vries, H.E. The Role of ATP-Binding Cassette Transporters in Neuro-Inflammation: Relevance for Bioactive Lipids. Front. Pharmacol. 2012, 3, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewinson, O.; Orelle, C.; Seeger, M.A. Structures of ABC transporters: Handle with care. Febs Lett. 2020, 594, 3799–3814. [Google Scholar] [CrossRef] [PubMed]
- Ford, R.C.; Beis, K. Learning the ABCs one at a time: Structure and mechanism of ABC transporters. Biochem. Soc. Trans. 2019, 47, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Tampe, R. Structural and Mechanistic Principles of ABC Transporters. Annu. Rev. Biochem. 2020, 89, 605–636. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Hotz, J.M.; Thomas, J.R.; Katz, E.N.; Robey, R.W.; Horibata, S.; Gottesman, M.M. ATP-binding cassette transporters at the zebrafish blood-brain barrier and the potential utility of the zebrafish as an in vivo model. Cancer Drug Resist. 2021, 4, 620–633. [Google Scholar] [CrossRef]
- Bugde, P.; Biswas, R.; Merien, F.; Lu, J.; Liu, D.X.; Chen, M.W.; Zhou, S.F.; Li, Y. The therapeutic potential of targeting ABC transporters to combat multi-drug resistance. Expert Opin. Ther. Targets 2017, 21, 511–530. [Google Scholar] [CrossRef]
- Tournier, N.; Goutal, S.; Mairinger, S.; Hernandez-Lozano, I.; Filip, T.; Sauberer, M.; Caille, F.; Breuil, L.; Stanek, J.; Freeman, A.F.; et al. Complete inhibition of ABCB1 and ABCG2 at the blood-brain barrier by co-infusion of erlotinib and tariquidar to improve brain delivery of the model ABCB1/ABCG2 substrate C-11 erlotinib. J. Cereb. Blood Flow Metab. 2021, 41, 1634–1646. [Google Scholar] [CrossRef]
- Dash, R.P.; Babu, R.J.; Srinivas, N.R. Therapeutic Potential and Utility of Elacridar with Respect to P-glycoprotein Inhibition: An Insight from the Published In Vitro, Preclinical and Clinical Studies. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 915–933. [Google Scholar] [CrossRef]
- Bauer, M.; Wulkersdorfer, B.; Karch, R.; Philippe, C.; Jager, W.; Stanek, J.; Wadsak, W.; Hacker, M.; Zeitlinger, M.; Langer, O. Effect of P-glycoprotein inhibition at the blood-brain barrier on brain distribution of (R)- C-11 verapamil in elderly vs. young subjects. Br. J. Clin. Pharmacol. 2017, 83, 1991–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Zepeda, D.; Taghi, M.; Scherrmann, J.M.; Decleves, X.; Menet, M.C. ABC Transporters at the Blood-Brain Interfaces, Their Study Models, and Drug Delivery Implications in Gliomas. Pharmaceutics 2020, 12, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, J.I.; Tseng, Y.J.; Chen, M.H.; Huang, C.Y.F.; Chang, P.M.H. Clinical Perspective of FDA Approved Drugs With P-Glycoprotein Inhibition Activities for Potential Cancer Therapeutics. Front. Oncol. 2020, 10, 2336. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, Y.; Katayama, K.; Kusuhara, H.; Nozaki, Y. Impact of P-Glycoprotein-Mediated Active Efflux on Drug Distribution into Lumbar Cerebrospinal Fluid in Nonhuman Primates. Drug Metab. Dispos. 2020, 48, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.W. Current Strategies for Brain Drug Delivery. Theranostics 2018, 8, 1481–1493. [Google Scholar] [CrossRef]
- Burgess, A.; Hynynen, K. Drug delivery across the blood-brain barrier using focused ultrasound. Expert Opin. Drug Deliv. 2014, 11, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Vagner, T.; Dvorzhak, A.; Wojtowicz, A.M.; Harms, C.; Grantyn, R. Systemic application of AAV vectors targeting GFAP-expressing astrocytes in Z-Q175-KI Huntington’s disease mice. Mol. Cell. Neurosci. 2016, 77, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Check, E. Gene therapy put on hold as third child develops cancer. Nature 2005, 433, 561. [Google Scholar] [CrossRef]
- Sarin, H. Recent progress towards development of effective systemic chemotherapy for the treatment of malignant brain tumors. J. Transl. Med. 2009, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.Z.; Fogarty, B.; LaForge, B.; Aziz, S.; Pham, T.; Lai, L.N.; Bai, S.H. Delivery of Small Interfering RNA to Inhibit Vascular Endothelial Growth Factor in Zebrafish Using Natural Brain Endothelia Cell-Secreted Exosome Nanovesicles for the Treatment of Brain Cancer. AAPS J. 2017, 19, 475–486. [Google Scholar] [CrossRef]
- Thomas, F.C.; Taskar, K.; Rudraraju, V.; Goda, S.; Thorsheim, H.R.; Gaasch, J.A.; Mittapalli, R.K.; Palmieri, D.; Steeg, P.S.; Lockman, P.R.; et al. Uptake of ANG1005, A Novel Paclitaxel Derivative, Through the Blood-Brain Barrier into Brain and Experimental Brain Metastases of Breast Cancer. Pharm. Res. 2009, 26, 2486–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Has, C.; Sunthar, P. A comprehensive review on recent preparation techniques of liposomes. J. Liposome Res. 2020, 30, 336–365. [Google Scholar] [CrossRef] [PubMed]
- Bayon-Cordero, L.; Alkorta, I.; Arana, L. Application of Solid Lipid Nanoparticles to Improve the Efficiency of Anticancer Drugs. Nanomaterials 2019, 9, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, M.M.; Patel, B.M. Crossing the Blood-Brain Barrier: Recent Advances in Drug Delivery to the Brain. CNS Drugs 2017, 31, 109–133. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.D.; Hu, G.Y.; Zhang, G.Y.; Zhao, G.; Shi, M. Ginsenoside Rd attenuates blood-brain barrier damage by suppressing proteasome-mediated signaling after transient forebrain ischemia. Neuroreport 2020, 31, 466–472. [Google Scholar] [CrossRef]
- Poznanski, P.; Lesniak, A.; Korostynski, M.; Sacharczuk, M. Ethanol consumption following mild traumatic brain injury is related to blood-brain barrier permeability. Addict. Biol. 2020, 25, e12683. [Google Scholar] [CrossRef]
- Li, Z.G.; Wang, Z.Y.; Xue, K.B.; Wang, Z.S.; Guo, C.; Qian, Y.; Li, X.Q.; Wei, Y.J. High concentration of blood cobalt is associated with the impairment of blood-brain barrier permeability. Chemosphere 2021, 273, 129579. [Google Scholar] [CrossRef]
- Fan, C.H.; Liu, H.L.; Huang, C.Y.; Ma, Y.J.; Yen, T.C.; Yeh, C.K. Detection Of Intracerebral Hemorrhage And Transient Blood-Supply Shortage In Focused-Ultrasound-Induced Blood-Brain Barrier Disruption By Ultrasound Imaging. Ultrasound Med. Biol. 2012, 38, 1372–1382. [Google Scholar] [CrossRef]
- Li, Y.J.; Wu, M.X.; Zhang, N.S.; Tang, C.Y.; Jiang, P.; Liu, X.; Yan, F.; Zheng, H.R. Mechanisms of enhanced antiglioma efficacy of polysorbate 80-modified paclitaxel-loaded PLGA nanoparticles by focused ultrasound. J. Cell. Mol. Med. 2018, 22, 4171–4182. [Google Scholar] [CrossRef]
- Yoshida, Y.; Sejimo, Y.; Kurachi, M.; Ishizaki, Y.; Nakano, T.; Takahashi, A. X-ray irradiation induces disruption of the blood-brain barrier with localized changes in claudin-5 and activation of microglia in the mouse brain. Neurochem. Int. 2018, 119, 199–206. [Google Scholar] [CrossRef]
- Sharabi, S.; Bresler, Y.; Ravid, O.; Shemesh, C.; Atrakchi, D.; Schnaider-Beeri, M.; Gosselet, F.; Dehouck, L.; Last, D.; Guez, D.; et al. Transient blood-brain barrier disruption is induced by low pulsed electrical fields in vitro: An analysis of permeability and trans-endothelial electric resistivity. Drug Deliv. 2019, 26, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Giri, P.; Mittal, L.; Camarillo, I.G.; Sundararajan, R. Analysis of Pathways in Triple-Negative Breast Cancer Cells Treated with the Combination of Electrochemotherapy and Cisplatin. Biointerface Res. Appl. Chem. 2021, 11, 13453–13464. [Google Scholar] [CrossRef]
- Hjouj, M.; Last, D.; Guez, D.; Daniels, D.; Sharabi, S.; Lavee, J.; Rubinsky, B.; Mardor, Y. MRI Study on Reversible and Irreversible Electroporation Induced Blood Brain Barrier Disruption. PLoS ONE 2012, 7, e42817. [Google Scholar] [CrossRef] [PubMed]
- Sharabi, S.; Last, D.; Guez, D.; Daniels, D.; Hjouj, M.I.; Salomon, S.; Maor, E.; Mardor, Y. Dynamic effects of point source electroporation on the rat brain tissue. Bioelectrochemistry 2014, 99, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Quintero, S.V.; Datta, A.; Amaya, R.; Elwassif, M.; Bikson, M.; Tarbell, J.M. DBS-relevant electric fields increase hydraulic conductivity of in vitro endothelial monolayers. J. Neural Eng. 2010, 7, 16005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonakdar, M.; Graybill, P.M.; Davalos, R.V. A microfluidic model of the blood-brain barrier to study permeabilization by pulsed electric fields. Rsc. Adv. 2017, 7, 42811–42818. [Google Scholar] [CrossRef]
- Calias, P.; Banks, W.A.; Begley, D.; Scarpa, M.; Dickson, P. Intrathecal delivery of protein therapeutics to the brain: A critical reassessment. Pharmacol. Ther. 2014, 144, 114–122. [Google Scholar] [CrossRef]
- Scarpa, M.; Bellettato, C.M.; Lampe, C.; Leader, D.; Begley, D.J. Neuronopathic lysosomal storage disorders: Approaches to treat the central nervous system. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 159–171. [Google Scholar] [CrossRef]
- Kakkis, E.D.; Muenzer, J.; Tiller, G.E.; Waber, L.; Belmont, J.; Passage, M.; Izykowski, B.; Phillips, J.; Doroshow, R.; Walot, I.; et al. Enzyme-replacement therapy in mucopolysaccharidosis I. N. Engl. J. Med. 2001, 344, 182–188. [Google Scholar] [CrossRef]
- Dickson, P.I.; Chen, A.H. Intrathecal enzyme replacement therapy for mucopolysaccharidosis I: Translating success in animal models to patients. Curr. Pharm. Biotechnol. 2011, 12, 946–955. [Google Scholar] [CrossRef]
- King, B.; Setford, M.L.; Hassiotis, S.; Trim, P.J.; Duplock, S.; Tucker, J.N.; Hattersley, K.; Snel, M.F.; Hopwood, J.J.; Hemsley, K.M. Low-dose, continual enzyme delivery ameliorates some aspects of established brain disease in a mouse model of a childhood-onset neurodegenerative disorder. Exp. Neurol. 2016, 278, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Winner, L.K.; Marshall, N.R.; Jolly, R.D.; Trim, P.J.; Duplock, S.K.; Snel, M.F.; Hemsley, K.M. Evaluation of Disease Lesions in the Developing Canine MPS IIIA Brain. In JIMD Reports; Morava, E., Baumgartner, M., Patterson, M., Rahman, S., Zschocke, J., Peters, V., Eds.; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 2019; Volume 43, pp. 91–101. [Google Scholar]
- Muenzer, J.; Hendriksz, C.J.; Fan, Z.; Vijayaraghavan, S.; Perry, V.; Santra, S.; Solanki, G.A.; Mascelli, M.A.; Pan, L.Y.; Wang, N.; et al. A phase I/II study of intrathecal idursulfase-IT in children with severe mucopolysaccharidosis II. Genet. Med. 2016, 18, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardridge, W.M. Blood-brain barrier drug targeting: The future of brain drug development. Mol. Interv. 2003, 3, 90-105, 151. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, J. Influence of the surface properties on nanoparticle-mediated transport of drugs to the brain. J. Nanosci. Nanotechnol. 2004, 4, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, N.; Niwa, T.; Yotsumoto, Y.; Sugiyama, Y. Impact of drug transporter studies on drug discovery and development. Pharmacol. Rev. 2003, 55, 425–461. [Google Scholar] [CrossRef]
- Bors, L.A.; Erdo, F. Overcoming the Blood-Brain Barrier. Challenges and Tricks for CNS Drug Delivery. Sci. Pharm. 2019, 87, 6. [Google Scholar] [CrossRef] [Green Version]
- Schally, A.V.; Nagy, A. Chemotherapy targeted to cancers through tumoral hormone receptors. Trends Endocrinol. Metab. 2004, 15, 300–310. [Google Scholar] [CrossRef]
- Elmagbari, N.O.; Egleton, R.D.; Palian, M.M.; Lowery, J.J.; Schmid, W.R.; Davis, P.; Navratilova, E.; Dhanasekaran, M.; Keyari, C.M.; Yamamura, H.I.; et al. Antinociceptive structure-activity studies with enkephalin-based opioid glycopeptides. J. Pharmacol. Exp. Ther. 2004, 311, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Palian, M.M.; Boguslavsky, V.I.; O’Brien, D.F.; Polt, R. Glycopeptide-membrane interactions: Glycosyl enkephalin analogues adopt turn conformations by NMR and CD in amphipathic media. J. Am. Chem. Soc. 2003, 125, 5823–5831. [Google Scholar] [CrossRef]
- Juillerat-Jeanneret, L.; Lohm, S.; Hamou, M.F.; Pinet, F. Regulation of aminopeptidase A in human brain tumor vasculature: Evidence for a role of transforming growth factor-beta. Lab. Investig. 2000, 80, 973–980. [Google Scholar] [CrossRef] [Green Version]
- Egleton, R.D.; Davis, T.P. Development of neuropeptide drugs that cross the blood-brain barrier. NeuroRx J. Am. Soc. Exp. Neuro Ther. 2005, 2, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Q.Y.; Haqqani, A.S.; Leclerc, S.; Liu, Z.; Fauteux, F.; Baumann, E.; Delaney, C.E.; Ly, D.; Star, A.T.; et al. Differential expression of receptors mediating receptor-mediated transcytosis (RMT) in brain microvessels, brain parenchyma and peripheral tissues of the mouse and the human. Fluids Barriers CNS 2020, 17, 47. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.M.; Gachulincova, I.; Ho, D.W.; Bailey, C.; Bartlett, C.A.; Norret, M.; Murphy, J.; Buckley, A.; Rigby, P.J.; House, M.J.; et al. An Unexpected Transient Breakdown of the Blood Brain Barrier Triggers Passage of Large Intravenously Administered Nanoparticles. Sci. Rep. 2016, 6, 22595. [Google Scholar] [CrossRef] [PubMed]
- Nilaver, G.; Muldoon, L.L.; Kroll, R.A.; Pagel, M.A.; Breakefield, X.O.; Davidson, B.L.; Neuwelt, E.A. Delivery Of Herpesvirus And Adenovirus To Nude Rat Intracerebral Tumors After Osmotic Blood-Brain-Barrier Disruption. Proc. Natl. Acad. Sci. USA 1995, 92, 9829–9833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louboutin, J.P.; Chekmasova, A.A.; Marusich, E.; Chowdhury, J.R.; Strayer, D.S. Efficient CNS gene delivery by intravenous injection. Nat. Methods 2010, 7, 905–907. [Google Scholar] [CrossRef]
- Fu, H.Y.; Muenzer, J.; Samulski, R.J.; Breese, G.; Sifford, J.; Zeng, X.H.; McCarty, D.M. Self-complementary adeno-associated virus serotype 2 vector: Global distribution and broad dispersion of AAV-mediated transgene expression in mouse brain. Mol. Ther. 2003, 8, 911–917. [Google Scholar] [CrossRef]
- McCarty, D.M.; DiRosario, J.; Gulaid, K.; Muenzer, J.; Fu, H. Mannitol-facilitated CNS entry of rAAV2 vector significantly delayed the neurological disease progression in MPS IIIB mice. Gene Ther. 2009, 16, 1340–1352. [Google Scholar] [CrossRef]
- Fu, H.Y.; Bartz, J.D.; Stephens, R.L.; McCarty, D.M. Peripheral Nervous System Neuropathology and Progressive Sensory Impairments in a Mouse Model of Mucopolysaccharidosis IIIB. PLoS ONE 2012, 7, e45992. [Google Scholar] [CrossRef]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W.D. Strategies for delivering therapeutics across the blood-brain barrier. Nat. Rev. Drug Discov. 2021, 20, 362–383. [Google Scholar] [CrossRef]
- Felix, M.S.; Borloz, E.; Metwally, K.; Dauba, A.; Larrat, B.; Matagne, V.; Ehinger, Y.; Villard, L.; Novell, A.; Mensah, S.; et al. Ultrasound-Mediated Blood-Brain Barrier Opening Improves Whole Brain Gene Delivery in Mice. Pharmaceutics 2021, 13, 1245. [Google Scholar] [CrossRef]
- Huang, L.; Wan, J.; Wu, Y.Q.; Tian, Y.; Yao, Y.Z.; Yao, S.; Ji, X.Y.; Wang, S.J.; Su, Z.L.; Xu, H.X. Challenges in adeno-associated virus-based treatment of central nervous system diseases through systemic injection. Life Sci. 2021, 270, 119142. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, M.; Hejmady, S.; Saha, R.N.; Damle, S.; Singhvi, G.; Alexander, A.; Kesharwani, P.; Dubey, S.K. Evolving new-age strategies to transport therapeutics across the blood-brain-barrier. Int. J. Pharm. 2021, 599, 120351. [Google Scholar] [CrossRef] [PubMed]
- Pulgar, V.M. Transcytosis to Cross the Blood Brain Barrier, New Advancements and Challenges. Front. Neurosci. 2019, 12, 1019. [Google Scholar] [CrossRef] [PubMed]
- Elliott, R.O.; He, M. Unlocking the Power of Exosomes for Crossing Biological Barriers in Drug Delivery. Pharmaceutics 2021, 13, 122. [Google Scholar] [CrossRef] [PubMed]
- Meldolesi, J. Exosomes and Ectosomes in Intercellular Communication. Curr. Biol. 2018, 28, R435–R444. [Google Scholar] [CrossRef] [Green Version]
- Alexander, M.; Ramstead, A.G.; Bauer, K.M.; Lee, S.-H.; Runtsch, M.C.; Wallace, J.; Huffaker, T.B.; Larsen, D.K.; Tolmachova, T.; Seabra, M.C.; et al. Rab27-Dependent Exosome Production Inhibits Chronic Inflammation and Enables Acute Responses to Inflammatory Stimuli. J. Immunol. 2017, 199, 3559–3570. [Google Scholar] [CrossRef] [Green Version]
- Dalkara, T.; Gursoy-Ozdemir, Y.; Yemisci, M. Brain microvascular pericytes in health and disease. Acta Neuropathol. 2011, 122, 1–9. [Google Scholar] [CrossRef]
- Mardpour, S.; Hamidieh, A.A.; Taleahmad, S.; Sharifzad, F.; Taghikhani, A.; Baharvand, H. Interaction between mesenchymal stromal cell-derived extracellular vesicles and immune cells by distinct protein content. J. Cell. Physiol. 2019, 234, 8249–8258. [Google Scholar] [CrossRef]
- Li, C.J.; Fang, Q.H.; Liu, M.L.; Lin, J.N. Current understanding of the role of Adipose-derived Extracellular Vesicles in Metabolic Homeostasis and Diseases: Communication from the distance between cells/tissues. Theranostics 2020, 10, 7422–7435. [Google Scholar] [CrossRef]
- Penfornis, P.; Vallabhaneni, K.C.; Whitt, J.; Pochampally, R. Extracellular vesicles as carriers of microRNA, proteins and lipids in tumor microenvironment. Int. J. Cancer 2016, 138, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, J.; Stewart, T.; Banks, W.A.; Zhang, J. The Transport Mechanism of Extracellular Vesicles at the Blood-Brain Barrier. Curr. Pharm. Des. 2017, 23, 6206–6214. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.J.; Liu, F.; He, X.; Yang, X.; Shan, F.P.; Feng, J. Exosomes derived from mesenchymal stem cells attenuate inflammation and demyelination of the central nervous system in EAE rats by regulating the polarization of microglia. Int. Immunopharmacol. 2019, 67, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, J.P.K.; Holme, M.N.; Stevens, M.M. Re-Engineering Extracellular Vesicles as Smart Nanoscale Therapeutics. Acs Nano 2017, 11, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Smyth, T.; Petrova, K.; Payton, N.M.; Persaud, I.; Redzic, J.S.; Graner, M.W.; Smith-Jones, P.; Anchordoquy, T.J. Surface functionalization of exosomes using click chemistry. Bioconjugate Chem. 2014, 25, 1777–1784. [Google Scholar] [CrossRef] [Green Version]
- Salunkhe, S.; Dheeraj; Basak, M.; Chitkara, D.; Mittal, A. Surface functionalization of exosomes for target-specific delivery and in vivo imaging & tracking: Strategies and significance. J. Control. Release 2020, 326, 599–614. [Google Scholar] [CrossRef]
- Sato, Y.T.; Umezaki, K.; Sawada, S.; Mukai, S.; Sasaki, Y.; Harada, N.; Shiku, H.; Akiyoshi, K. Engineering hybrid exosomes by membrane fusion with liposomes. Sci. Rep. 2016, 6, 21933. [Google Scholar] [CrossRef] [Green Version]
- Kooijmans, S.A.A.; Fliervoet, L.A.L.; van der Meel, R.; Fens, M.; Heijnen, H.F.G.; Henegouwen, P.; Vader, P.; Schiffelers, R.M. PEGylated and targeted extracellular vesicles display enhanced cell specificity and circulation time. J. Control. Release 2016, 224, 77–85. [Google Scholar] [CrossRef]
- Yu, J.E.; Han, S.Y.; Wolfson, B.; Zhou, Q. The role of endothelial lipase in lipid metabolism, inflammation and cancer. Histol. Histopathol. 2018, 33, 1–10. [Google Scholar] [CrossRef]
- Ye, Z.; Zhang, T.; He, W.; Jin, H.; Liu, C.; Yang, Z.; Ren, J. Methotrexate-Loaded Extracellular Vesicles Functionalized with Therapeutic and Targeted Peptides for the Treatment of Glioblastoma Multiforme. ACS Appl. Mater. Interfaces 2018, 10, 12341–12350. [Google Scholar] [CrossRef]
- Hama, H.; Saito, A.; Takeda, T.; Tanuma, A.; Xie, Y.S.; Sato, K.; Kazama, J.J.; Gejyo, F. Evidence indicating that renal tubular metabolism of leptin is mediated by megalin but not by the leptin receptors. Endocrinology 2004, 145, 3935–3940. [Google Scholar] [CrossRef] [Green Version]
- Dhanda, D.S.; Frey, W.; Leopold, D.; Kompella, U.B. Approaches for drug deposition in the human olfactory epithelium. Drug Deliv. Technol. 2005, 5, 64–72. [Google Scholar]
- Hanson, L.R.; Frey, W.H., 2nd. Intranasal delivery bypasses the blood-brain barrier to target therapeutic agents to the central nervous system and treat neurodegenerative disease. BMC Neurosci. 2008, 9 (Suppl. 3), S5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Center for Biotechnology Information. PubChem Patent Summary for WO-9107947-A1. Available online: https://pubchem.ncbi.nlm.nih.gov/patent/WO-9107947-A1 (accessed on 16 April 2022).
- Frey, W.H.; Liu, J.; Chen, X.Q.; Thorne, R.G.; Fawcett, J.R.; Ala, T.A.; Rahman, Y.E. Delivery of I-125-NGF to the brain via the olfactory route. Drug Deliv. 1997, 4, 87–92. [Google Scholar] [CrossRef]
- Ross, T.M.; Martinez, P.M.; Renner, J.C.; Thorne, R.G.; Hanson, L.R.; Frey, W.H. Intranasal administration of interferon beta bypasses the blood-brain barrier to target the central nervous system and cervical lymph nodes: A non-invasive treatment strategy for multiple sclerosis. J. Neuroimmunol. 2004, 151, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Thorne, R.G.; Pronk, G.J.; Padmanabhan, V.; Frey, W.H. Delivery of insulin-like growth factor-I to the rat brain and spinal cord along olfactory and trigeminal pathways following intranasal administration. Neuroscience 2004, 127, 481–496. [Google Scholar] [CrossRef]
- De Rosa, R.; Garcia, A.A.; Braschi, C.; Capsoni, S.; Maffei, L.; Berardi, N.; Cattaneo, A. Intranasal administration of nerve growth factor (NGF) rescues recognition memory deficits in AD11 anti-NGF transgenic mice. Proc. Natl. Acad. Sci. USA 2005, 102, 3811–3816. [Google Scholar] [CrossRef] [Green Version]
- Capsoni, S.; Giannotta, S.; Cattaneo, A. Nerve growth factor and galantamine ameliorate early signs of neurodegeneration in anti-nerve growth factor mice. Proc. Natl. Acad. Sci. USA 2002, 99, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Fawcett, J.R.; Hanson, L.R.; Frey, W.H., 2nd. The window of opportunity for treatment of focal cerebral ischemic damage with noninvasive intranasal insulin-like growth factor-I in rats. J. Stroke Cerebrovasc. Dis. Off. J. Natl. Stroke Assoc. 2004, 13, 16–23. [Google Scholar] [CrossRef]
- Jin, K.L.; Xie, L.; Childs, J.; Sun, Y.J.; Mao, X.O.; Logvinova, A.; Greenberg, D.A. Cerebral neurogenesis is induced by intranasal administration of growth factors. Ann. Neurol. 2003, 53, 405–409. [Google Scholar] [CrossRef]
- Sek, K.; Molck, C.; Stewart, G.D.; Kats, L.; Darcy, P.K.; Beavis, P.A. Targeting Adenosine Receptor Signaling in Cancer Immunotherapy. Int. J. Mol. Sci. 2018, 19, 3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Effendi, W.I.; Nagano, T.; Kobayashi, K.; Nishimura, Y. Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases. Cells 2020, 9, 785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ongini, E.; Fredholm, B.B. Pharmacology of adenosine A2A receptors. Trends Pharmacol. Sci. 1996, 17, 364–372. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Ijzerman, A.P.; Jacobson, K.A.; Linden, J.; Muller, C.E. International Union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and Classification of Adenosine Receptors-An Update. Pharmacol. Rev. 2011, 63, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Bynoe, M.S.; Viret, C.; Yan, A.; Kim, D.-G. Adenosine receptor signaling: A key to opening the blood-brain door. Fluids Barriers CNS 2015, 12, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledent, C.; Vaugeois, J.M.; Schiffmann, S.N.; Pedrazzini, T.; ElYacoubi, M.; Vanderhaeghen, J.J.; Costentin, J.; Heath, J.K.; Vassart, G.; Parmentier, M. Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A(2a) receptor. Nature 1997, 388, 674–678. [Google Scholar] [CrossRef]
- Zhao, G.; Linke, A.; Xu, X.B.; Ochoa, M.; Belloni, F.; Belardinelli, L.; Hintze, T.H. Comparative profile of vasodilation by CVT-3146, a novel A(2A) receptor agonist, and adenosine in conscious dogs. J. Pharmacol. Exp. Ther. 2003, 307, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Wala, K.; Szlasa, W.; Saczko, J.; Rudno-Rudzinska, J.; Kulbacka, J. Modulation of Blood-Brain Barrier Permeability by Activating Adenosine A2 Receptors in Oncological Treatment. Biomolecules 2021, 11, 633. [Google Scholar] [CrossRef]
- Glaser, F.; Steinberg, D.M.; Vakser, I.A.; Ben-Tal, N. Residue frequencies and pairing preferences at protein-protein interfaces. Proteins 2001, 43, 89–102. [Google Scholar] [CrossRef]
- Bobermin, L.D.; Roppa, R.H.A.; Quincozes-Santos, A. Adenosine receptors as a new target for resveratrol-mediated glioprotection. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 634–647. [Google Scholar] [CrossRef]
- Zhang, J.L.; Liu, H.; Du, X.S.; Guo, Y.; Chen, X.; Wang, S.N.; Fang, J.Q.; Cao, P.; Zhang, B.; Liu, Z.; et al. Increasing of Blood-Brain Tumor Barrier Permeability through Transcellular and Paracellular Pathways by Microbubble-Enhanced Diagnostic Ultrasound in a C6 Glioma Model. Front. Neurosci. 2017, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Sheikov, N.; McDannold, N.; Sharma, S.; Hynynen, K. Effect of focused ultrasound applied with an ultrasound contrast agent on the tight junctional integrity of the brain microvascular endothelium. Ultrasound Med. Biol. 2008, 34, 1093–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Aryal, M.; Vykhodtseva, N.; Zhang, Y.Z.; McDannold, N. Evaluation of permeability, doxorubicin delivery, and drug retention in a rat brain tumor model after ultrasound-induced blood-tumor barrier disruption. J. Control. Release 2017, 250, 77–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, M.; Fischer, K.; Gentile, C.; Gitto, S.; Zhang, Y.Z.; McDannold, N. Effects on P-Glycoprotein Expression after Blood-Brain Barrier Disruption Using Focused Ultrasound and Microbubbles. PLoS ONE 2017, 12, e0166061. [Google Scholar] [CrossRef] [Green Version]
- McMahon, D.; Oakden, W.; Hynynen, K. Investigating the effects of dexamethasone on blood-brain barrier permeability and inflammatory response following focused ultrasound and microbubble exposure. Theranostics 2020, 10, 1604–1618. [Google Scholar] [CrossRef]
- Samiotaki, G.; Karakatsani, M.E.; Buch, A.; Papadopoulos, S.; Wu, S.Y.; Jambawalikar, S.; Konofagou, E.E. Pharmacokinetic analysis and drug delivery efficiency of the focused ultrasound-induced blood-brain barrier opening in non-human primates. Magn. Reson. Imaging 2017, 37, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, A.; Liu, M.; Ojha, T.; Storm, G.; Kiessling, F.; Lammers, T. Ultrasound-mediated drug delivery to the brain: Principles, progress and prospects. Drug Discov. Today. Technol. 2016, 20, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Xhima, K.; Weber-Adrian, D.; Silburt, J. Glutamate Induces Blood-Brain Barrier Permeability through Activation of N-Methyl-D-Aspartate Receptors. J. Neurosci. 2016, 36, 12296–12298. [Google Scholar] [CrossRef] [Green Version]
- Beisteiner, R.; Matt, E.; Fan, C.; Baldysiak, H.; Schonfeld, M.; Novak, T.P.; Amini, A.; Aslan, T.; Reinecke, R.; Lehrner, J.; et al. Transcranial Pulse Stimulation with Ultrasound in Alzheimer’s Disease—A New Navigated Focal Brain Therapy. Adv. Sci. 2020, 7, 1902583. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Pople, C.B.; Lea-Banks, H.; Abrahao, A.; Davidson, B.; Suppiah, S.; Vecchio, L.M.; Samuel, N.; Mahmud, F.; Hynynen, K.; et al. Safety and efficacy of focused ultrasound induced blood-brain barrier opening, an integrative review of animal and human studies. J. Control. Release 2019, 309, 25–36. [Google Scholar] [CrossRef]
- Villringer, K.; Cuesta, B.E.S.; Ostwaldt, A.C.; Grittner, U.; Brunecker, P.; Khalil, A.A.; Schindler, K.; Eisenblatter, O.; Audebert, H.; Fiebach, J.B. DCE-MRI blood-brain barrier assessment in acute ischemic stroke. Neurology 2017, 88, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Q.; Wang, Z.Y.; Qi, C.F.; Han, X.N.; Lan, X.; Wan, J.R.; Wang, W.Z.; Zhao, X.C.; Hou, Z.P.; et al. Multimodality MRI assessment of grey and white matter injury and blood-brain barrier disruption after intracerebral haemorrhage in mice. Sci. Rep. 2017, 7, 40358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiraishi, K.; Kawano, K.; Minowa, T.; Maitani, Y.; Yokoyama, M. Preparation and in vivo imaging of PEG-poly(L-lysine)-based polymeric micelle MRI contrast agents. J. Control. Release 2009, 136, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Posadas, I.; Monteagudo, S.; Ceña, V. Nanoparticles for brain-specific drug and genetic material delivery, imaging and diagnosis. Nanomed. Lond. Engl. 2016, 11, 833–849. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, S.; Archunan, G.; Sivakumar, M.; Selvan, S.T.; Fred, A.L.; Kumar, S.; Gulyas, B.; Padmanabhan, P. Theranostic applications of nanoparticles in neurodegenerative disorders. Int. J. Nanomed. 2018, 13, 5561–5576. [Google Scholar] [CrossRef] [Green Version]
- Teleanu, D.M.; Chircov, C.; Grumezescu, A.M.; Volceanov, A.; Teleanu, R.I. Contrast Agents Delivery: An Up-to-Date Review of Nanodiagnostics in Neuroimaging. Nanomaterials 2019, 9, 542. [Google Scholar] [CrossRef] [Green Version]
- Richard, S.; Saric, A.; Boucher, M.; Slomianny, C.; Geffroy, F.; Mériaux, S.; Lalatonne, Y.; Petit, P.X.; Motte, L. Antioxidative Theranostic Iron Oxide Nanoparticles toward Brain Tumors Imaging and ROS Production. ACS Chem. Biol. 2016, 11, 2812–2819. [Google Scholar] [CrossRef]
- Richard, S.; Boucher, M.; Lalatonne, Y.; Meriaux, S.; Motte, L. Iron oxide nanoparticle surface decorated with cRGD peptides for magnetic resonance imaging of brain tumors. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1515–1520. [Google Scholar] [CrossRef]
- Meola, A.; Rao, J.H.; Chaudhary, N.; Sharma, M.; Chang, S.D. Gold Nanoparticles for Brain tumor imaging: A Systematic Review. Front. Neurol. 2018, 9, 328. [Google Scholar] [CrossRef]
- Mahan, M.M.; Doiron, A.L. Gold Nanoparticles as X-Ray, CT, and Multimodal Imaging Contrast Agents: Formulation, Targeting, and Methodology. J. Nanomater. 2018, 2018, 5837276. [Google Scholar] [CrossRef]
- Cho, J.H.; Kim, A.R.; Kim, S.H.; Lee, S.J.; Chung, H.; Yoon, M.Y. Development of a novel imaging agent using peptide-coated gold nanoparticles toward brain glioma stem cell marker CD133. Acta Biomater. 2017, 47, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Rammohan, N.; Holbrook, R.J.; Rotz, M.W.; MacRenaris, K.W.; Preslar, A.T.; Carney, C.E.; Reichova, V.; Meade, T.J. Gd(III)-Gold Nanoconjugates Provide Remarkable Cell Labeling for High Field Magnetic Resonance Imaging. Bioconjugate Chem. 2016, 28, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Azria, D.; Blanquer, S.; Verdier, J.M.; Belamie, E. Nanoparticles as contrast agents for brain nuclear magnetic resonance imaging in Alzheimer’s disease diagnosis. J. Mater. Chem. B 2017, 5, 7216–7237. [Google Scholar] [CrossRef] [PubMed]
- Gale, E.M.; Caravan, P. Gadolinium-Free Contrast Agents for Magnetic Resonance Imaging of the Central Nervous System. Acs Chem. Neurosci. 2018, 9, 395–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.X.; Zhu, Q.X.; Zeng, Y.; Zeng, Q.; Chen, X.L.; Zhan, Y.H. Manganese Oxide Nanoparticles as MRI Contrast Agents in Tumor Multimodal Imaging and Therapy. Int. J. Nanomed. 2019, 14, 8321–8344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, D.B.; Gamarra, L.F. Getting into the brain: Liposome-based strategies for effective drug delivery across the blood-brain barrier. Int. J. Nanomed. 2016, 11, 5381–5414. [Google Scholar] [CrossRef] [Green Version]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal Formulations in Clinical Use: An Updated Review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef]
- Pattni, B.S.; Chupin, V.V.; Torchilin, V.P. New Developments in Liposomal Drug Delivery. Chem. Rev. 2015, 115, 10938–10966. [Google Scholar] [CrossRef]
- Loyse, A.; Thangaraj, H.; Easterbrook, P.; Ford, N.; Roy, M.; Chiller, T.; Govender, N.; Harrison, T.S.; Bicanic, T. Cryptococcal meningitis: Improving access to essential antifungal medicines in resource-poor countries. Lancet Infect. Dis. 2013, 13, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Benesch, M.; Urban, C. Liposomal cytarabine for leukemic and lymphomatous meningitis: Recent developments. Expert Opin. Pharmacother. 2008, 9, 301–309. [Google Scholar] [CrossRef]
- Rompicherla, S.K.L.; Arumugam, K.; Bojja, S.L.; Kumar, N.; Rao, C.M. Pharmacokinetic and pharmacodynamic evaluation of nasal liposome and nanoparticle based rivastigmine formulations in acute and chronic models of Alzheimer’s disease. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 1737–1755. [Google Scholar] [CrossRef] [PubMed]
- Razay, G.; Wilcock, G.K. Galantamine in Alzheimer’s disease. Expert Rev. Neurother. 2008, 8, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Amanzadeh, E.; Esmaeili, A.; Rahgozar, S.; Nourbakhshnia, M. Application of quercetin in neurological disorders: From nutrition to nanomedicine. Rev. Neurosci. 2019, 30, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Karthika, C.; Appu, A.P.; Akter, R.; Rahman, M.H.; Tagde, P.; Ashraf, G.M.; Abdel-Daim, M.M.; ul Hassan, S.S.; Abid, A.; Bungau, S. Potential innovation against Alzheimer’s disorder: A tricomponent combination of natural antioxidants (vitamin E, quercetin, and basil oil) and the development of its intranasal delivery. Environ. Sci. Pollut. Res. 2022, 29, 10950–10965. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Yankee, E.L. A review on Parkinson’s disease treatment. Neuroimmunol. Neuroinflammation 2021, 8, 222–244. [Google Scholar] [CrossRef]
- Ovallath, S.; Sulthana, B. Levodopa: History and Therapeutic Applications. Ann. Indian Acad. Neurol. 2017, 20, 185–189. [Google Scholar] [CrossRef]
- Lopalco, A.; Cutrignelli, A.; Denora, N.; Lopedota, A.; Franco, M.; Laquintana, V. Transferrin Functionalized Liposomes Loading Dopamine HCl: Development and Permeability Studies across an In Vitro Model of Human Blood-Brain Barrier. Nanomaterials 2018, 8, 178. [Google Scholar] [CrossRef] [Green Version]
- Jhaveri, A.; Deshpande, P.; Pattni, B.; Torchilin, V. Transferrin-targeted, resveratrol-loaded liposomes for the treatment of glioblastoma. J. Control. Release 2018, 277, 89–101. [Google Scholar] [CrossRef]
- Lakkadwala, S.; Rodrigues, B.D.; Sun, C.W.; Singh, J. Dual functionalized liposomes for efficient co-delivery of anti-cancer chemotherapeutics for the treatment of glioblastoma. J. Control. Release 2019, 307, 247–260. [Google Scholar] [CrossRef]
- Article, R.; Sarangi, M.; Padhi, S. Solid Lipid Nanoparticles—A Review. J. Crit. Rev. 2016, 3, 5–12. [Google Scholar]
- Paliwal, R.; Paliwal, S.R.; Kenwat, R.; Kurmi, B.D.; Sahu, M.K. Solid lipid nanoparticles: A review on recent perspectives and patents. Expert Opin. Ther. Pat. 2020, 30, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Tatiparti, K.; Sau, S.; Iyer, A.K. Recent advances in nano delivery systems for blood-brain barrier (BBB) penetration and targeting of brain tumors. Drug Discov. Today 2021, 26, 1944–1952. [Google Scholar] [CrossRef] [PubMed]
- Graverini, G.; Piazzini, V.; Landucci, E.; Pantano, D.; Nardiello, P.; Casamenti, F.; Pellegrini-Giampietro, D.E.; Bilia, A.R.; Bergonzi, M.C. Solid lipid nanoparticles for delivery of andrographolide across the blood-brain barrier: In vitro and in vivo evaluation. Colloids Surf. B Biointerfaces 2018, 161, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Mansor, N.I.; Nordin, N.; Mohamed, F.; Ling, K.H.; Rosli, R.; Hassan, Z. Crossing the Blood-Brain Barrier: A Review on Drug Delivery Strategies for Treatment of the Central Nervous System Diseases. Curr. Drug Deliv. 2019, 16, 698–711. [Google Scholar] [CrossRef]
- Huang, R.; Zhu, Y.; Lin, L.; Song, S.; Cheng, L.; Zhu, R. Solid Lipid Nanoparticles Enhanced the Neuroprotective Role of Curcumin against Epilepsy through Activation of Bcl-2 Family and P38 MAPK Pathways. ACS Chem. Neurosci. 2020, 11, 1985–1995. [Google Scholar] [CrossRef]
- Muheem, A.; Jahangir, M.A.; Jaiswal, C.P.; Jafar, M.; Ahmad, M.Z.; Ahmad, J.; Warsi, M.H. Recent Patents, Regulatory Issues, and Toxicity of Nanoparticles in Neuronal Disorders. Curr. Drug Metab. 2021, 22, 263–279. [Google Scholar] [CrossRef]
- Ohta, S.; Kikuchi, E.; Ishijima, A.; Azuma, T.; Sakuma, I.; Ito, T. Investigating the optimum size of nanoparticles for their delivery into the brain assisted by focused ultrasound-induced blood–brain barrier opening. Sci. Rep. 2020, 10, 18220. [Google Scholar] [CrossRef]
- Vega, R.; Zhang, Y.; Curley, C.; Price, R.; Abounader, R. 370 Magnetic Resonance-Guided Focused Ultrasound Delivery of Polymeric Brain-Penetrating Nanoparticle MicroRNA Conjugates in Glioblastoma. Neurosurgery 2016, 63, 210. [Google Scholar] [CrossRef]
- Nowak, M.; Helgeson, M.E.; Mitragotri, S. Delivery of Nanoparticles and Macromolecules across the Blood–Brain Barrier. Remote Sens. 2020, 3, 1900073. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Walker, J.B.; Minic, Z.; Liu, F.C.; Goshgarian, H.; Mao, G.Z. Transporter protein and drug-conjugated gold nanoparticles capable of bypassing the blood-brain barrier. Sci. Rep. 2016, 6, 25794. [Google Scholar] [CrossRef]
- Fornaguera, C.; Lazaro, M.A.; Brugada-Vila, P.; Porcar, I.; Morera, I.; Guerra-Rebollo, M.; Garrido, C.; Rubio, N.; Blanco, J.; Cascante, A.; et al. Application of an assay Cascade methodology for a deep preclinical characterization of polymeric nanoparticles as a treatment for gliomas. Drug Deliv. 2018, 25, 472–483. [Google Scholar] [CrossRef]
- Liu, D.Z.; Cheng, Y.; Cai, R.Q.; Wang, W.W.; Cui, H.; Liu, M.; Zhang, B.L.; Mei, Q.B.; Zhou, S.Y. The enhancement of siPLK1 penetration across BBB and its anti glioblastoma activity in vivo by magnet and transferrin co-modified nanoparticle. Nanomed. -Nanotechnol. Biol. Med. 2018, 14, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Rennick, J.J.; Johnston, A.P.R.; Parton, R.G. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 2021, 16, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Santos, B.; Gremião, M.P.; Chorilli, M. Nanotechnology-based drug delivery systems for the treatment of Alzheimer’s disease. Int. J. Nanomed. 2015, 10, 4981–5003. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Gao, X. Brain Targeted Drug Delivery Systems: A Focus on Nanotechnology and Nanoparticulates; Elsevier Science: London, UK, 2018. [Google Scholar]
- Kiriiri, G.K.; Njogu, P.M.; Mwangi, A.N. Exploring different approaches to improve the success of drug discovery and development projects: A review. Future J. Pharm. Sci. 2020, 6, 27. [Google Scholar] [CrossRef]
- Van Norman, G.A. Limitations of Animal Studies for Predicting Toxicity in Clinical Trials Is it Time to Rethink Our Current Approach? Jacc Basic Transl. Sci. 2019, 4, 845–854. [Google Scholar] [CrossRef] [PubMed]
- GlobalDataHealthcare. Central Nervous System Resumed Trials Showed Biggest Increase in 2021. Available online: https://www.clinicaltrialsarena.com/comment/central-nervous-system-resumed-trials (accessed on 5 April 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Procedures | Strategies | Advantages | Limitations | References |
|---|---|---|---|---|
| Invasive approaches | Blood–brain barrier transient disruption | Transient opening; can achieve therapeutic concentrations | Non-targeted, entire BBB is disrupted | [47,48] |
| Intracerebroventricular and intrathecal infusion | High gene transfection efficiency | Safety concerns; direct brain injection; crossing the BBB; high dose by intravenous administration | [47,49,50] | |
| Non-invasive approaches | Chemical modification of drugs | Option for personalized medicine | Difficult to achieve therapeutic concentration in vivo; expensive and difficult to develop; non-targeted | [48] |
| Virus-mediated blood–brain barrier delivery | High gene transfection efficiency | Safety concerns; direct brain injection; crossing the BBB; high dose by intravenous administration | [47,49,50] | |
| Exosome-mediated blood–brain barrier delivery | Gene delivery to the brain; potential ability to cross the BBB | Exosome donor cells; loading procedure; in-vivo toxicity and pharmacokinetics | [47] | |
| Intranasal route of delivery | Bypass the BBB through nasal administration | Suitable for low dose | [51] | |
| Modulating blood–brain barrier permeability | Transiently open the BBB | Mismatch between findings in rodents and humans | [52,53] | |
| Focused ultrasound for brain diseases | Therapeutic concentrations; targeted to sub-millimeter regions in the brain | Only relatively short-term studies have been performed | [48] | |
| Nanoparticles for brain imaging/diagnostics | Enhance imaging; cross the BBB through the hyper-permeable BBB under disease conditions | Cross the BBB; understand dynamic changes in the BBB | [22] | |
| Liposome-based strategies | Exhibit strong biocompatibility and biodegradability; minimal toxicity; drug-targeted delivery; controlled drug release | Liposomes are not currently used in clinical practice to deliver brain-specific drugs | [54] | |
| Solid-lipid nanoparticles | Site-specific targeted delivery (via receptor-mediated transcytosis across brain capillary endothelial cells); physical stability; ability to escape the reticulo-endothelial system; extended blood circulation time; sustained release; nontoxic; biodegradable; biocompatible | Smaller drug payload; a complicated physical state of the lipid content; storage and administration stability issues | [55] |
| Primary Method | Sub-Method | Description | References |
|---|---|---|---|
| Paracellular pathway | - | Ultrasound/microbubbles and osmotic pressure are two methods for disrupting tight junctions between neighboring endothelial cells, both of which increase BBB permeability locally, allowing nanoparticle entrance. However, when the BBB’s homeostatic role is lost, this strategy carries certain hazards, as it would enable uncontrolled entrance of various substances into the brain, potentially resulting in cerebral toxicity. | [182] |
| Transcellular diffusion | - | The simplest transcellular route involves passive diffusion through the cell membrane and cytoplasm. To cross the phospholipidic bilayer of the membrane, nanoparticles must be small enough and at least partially lipophilic. | [183] |
| Transcytosis pathway | Adsorptive transcytosis | The surface characteristics of the nanoparticles make it easier for the nanoparticle and its payload to attach to the endothelial cells’ luminal plasma membrane. Because the plasma membrane of endothelial cells is negatively charged, positively charged nanoparticles are more likely to undergo this process than neutral or negatively charged ones. Nanoparticles coated with wheat germ agglutinin, for example, may be taken up by nerve terminals and retrogradely transported to the CNS by axons. | [184] |
| Receptor-mediated transcytosis | Where nanoparticles with various ligands on their surfaces can bind to certain receptors and so enhance endocytosis GLUT1, lactoferrin (Lf), transferrin (Tf), or other peptides such as angiopep-2 or Seq12 have all been employed as targets. | [185,186] | |
| Endocytosis | Clathrin-mediated endocytosis | Endocytotic vesicles up to 200 nm in diameter are formed in clathrin-enriched portions of the cell membrane. Once within the cell, clathrin-coated vesicles fuse together to form an early endosome, which progresses to late endosomes when the intravesicular pH drops and, eventually, lysosomes, triggering cargo destruction. To be an effective delivery agent, the nanoparticle must make it easier for its payload to escape from endosomes before they merge with lysosomes, preventing cargo destruction. | [187] |
| Caveolin-mediated endocytosis | This occurs in lipid rafts and results in plasma membrane invaginations of roughly 80 nm in size. Following this first stage, caveolin vesicles merge with other caveolin vesicles to form caveosomes, which evade lysosomes and have a variety of fates depending on the cell type. | [187] |
| Study Title | Intervention/Treatment | Status (as Reported until 15 April 2022) | ClinicalTrials.gov Identifier |
|---|---|---|---|
| Blood–Brain Barrier Permeability Study in Adults with Meningitis (NM-BBBP) | Other: ICG-PULSION Device: LiMON, Pulsion Medical Systems Other: Gadolinium (Gadovist, Bayer, Germany) | Terminated | NCT02902588 |
| The Relevance of the Blood–Brain Barrier to Cognitive Dysfunction and Alzheimer’s Disease | Diagnostic Test: Contrast agent enhanced MRI using Gadovist | Completed | NCT04093882 |
| Blood–Brain-Barrier Opening Using Focused Ultrasound with IV Contrast Agents in Patients With Early Alzheimer’s Disease (BBB-Alzheimers) | Device: BBB opening | Completed | NCT02986932 |
| Surgical Tissue Flap to Bypass the Blood Brain Barrier in GBM | Procedure: Tissue autograft of pedicled temporoparietal fascial (TPF) or pericranial flap to bypass the blood brain barrier (BBB) | Recruiting | NCT03630289 |
| Laparoscopically Harvested Omental Free Tissue Autograft to Bypass the Blood Brain Barrier (BBB) in Human Recurrent Glioblastoma Multiforme (rGBM) | Procedure: Laparoscopically harvested omental free flap | Recruiting | NCT04222309 |
| Blood–Brain Barrier Opening Using MR-Guided Focused Ultrasound in Patients with Amyotrophic Lateral Sclerosis | Device: Blood–Brain Barrier opening with MRgFUS | Active, not recruiting | NCT03321487 |
| The Level of Blood–Brain Barrier Damage Biomarker in Acute Ischemic Stroke | Diagnostic Test: the level of biomarker in blood | Recruiting | NCT05321225 |
| Non-invasive Blood–Brain Barrier Opening in Alzheimer’s Disease Patients Using Focused Ultrasound | Device: Neuronavigation-guided single-element focused ultrasound transducer Drug: Definity Other: Magnetic Resonance Imaging (MRI) with or without gadolinium contrast agents Other: Positron Emission Tomography (PET) Other: Amyvid | Recruiting | NCT04118764 |
| Blood–Brain Barrier Penetration of Therapeutic Agents in Human (BRIAN) | Drug: ODM-104 Drug: Paracetamol | Completed | NCT04571996 |
| A Study to Evaluate Temporary Blood–Brain Barrier Disruption in Patients with Parkinson’s Disease Dementia | Device: MR Guided Focused Ultrasound Blood Brain Barrier Disruption using FUS Other Name: ExAblate | Active, not recruiting | NCT03608553 |
| ExAblate Blood–Brain Barrier Opening for Treatment of Alzheimer’s Disease | Device: Blood Brain Barrier (BBB) Disruption | Recruiting | NCT03739905 |
| ExAblate Blood–Brain Barrier (BBB) Disruption for the Treatment of Alzheimer’s Disease | Device: Blood Brain Barrier (BBB) Disruption | Recruiting | NCT03671889 |
| Blood Brain Barrier Dysfunction and Postoperative Neurocognitive Disorders (BBBSx) | Diagnostic Test: Brain Imaging Diagnostic Test: Cognitive Testing Diagnostic Test: Blood Biomarkers | Recruiting | NCT04566562 |
| Ultrasound-based Blood–Brain Barrier Opening and Albumin-bound Paclitaxel for Recurrent Glioblastoma (SC9/ABX) | Device: Sonication for opening of blood–brain barrier Drug: Chemotherapy, albumin-bound paclitaxel | Recruiting | NCT04528680 |
| Safety Study of the Repeated Opening of the Blood–Brain Barrier with the SonoCloud® Device to Treat Malignant Brain Tumors in Pediatric Patients (SONOKID) | Device: SonoCloud® (9 transducers) | Not yet recruiting | NCT05293197 |
| Developing Advanced Blood–Brain Barrier Permeability Imaging for Early AD | Device: GRASP DCE-MRI Device: 3T Brain Scan | Recruiting | NCT03389698 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teleanu, R.I.; Preda, M.D.; Niculescu, A.-G.; Vladâcenco, O.; Radu, C.I.; Grumezescu, A.M.; Teleanu, D.M. Current Strategies to Enhance Delivery of Drugs across the Blood–Brain Barrier. Pharmaceutics 2022, 14, 987. https://doi.org/10.3390/pharmaceutics14050987
Teleanu RI, Preda MD, Niculescu A-G, Vladâcenco O, Radu CI, Grumezescu AM, Teleanu DM. Current Strategies to Enhance Delivery of Drugs across the Blood–Brain Barrier. Pharmaceutics. 2022; 14(5):987. https://doi.org/10.3390/pharmaceutics14050987
Chicago/Turabian StyleTeleanu, Raluca Ioana, Manuela Daniela Preda, Adelina-Gabriela Niculescu, Oana Vladâcenco, Crina Ioana Radu, Alexandru Mihai Grumezescu, and Daniel Mihai Teleanu. 2022. "Current Strategies to Enhance Delivery of Drugs across the Blood–Brain Barrier" Pharmaceutics 14, no. 5: 987. https://doi.org/10.3390/pharmaceutics14050987