
Persistent Properties of a Subpopulation of Cancer Cells Overexpressing the Hedgehog Receptor Patched



 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Biological Material
2.2. Flow Cytometry
2.3. Immunofluorescence
2.4. Western-Blot
2.5. Cytotoxicity Assay
2.6. Doxorubicin Accumulation in Cells
2.7. Clonogenic Assay
2.8. Transwell Invasion Assay
2.9. Wound-Healing Assay
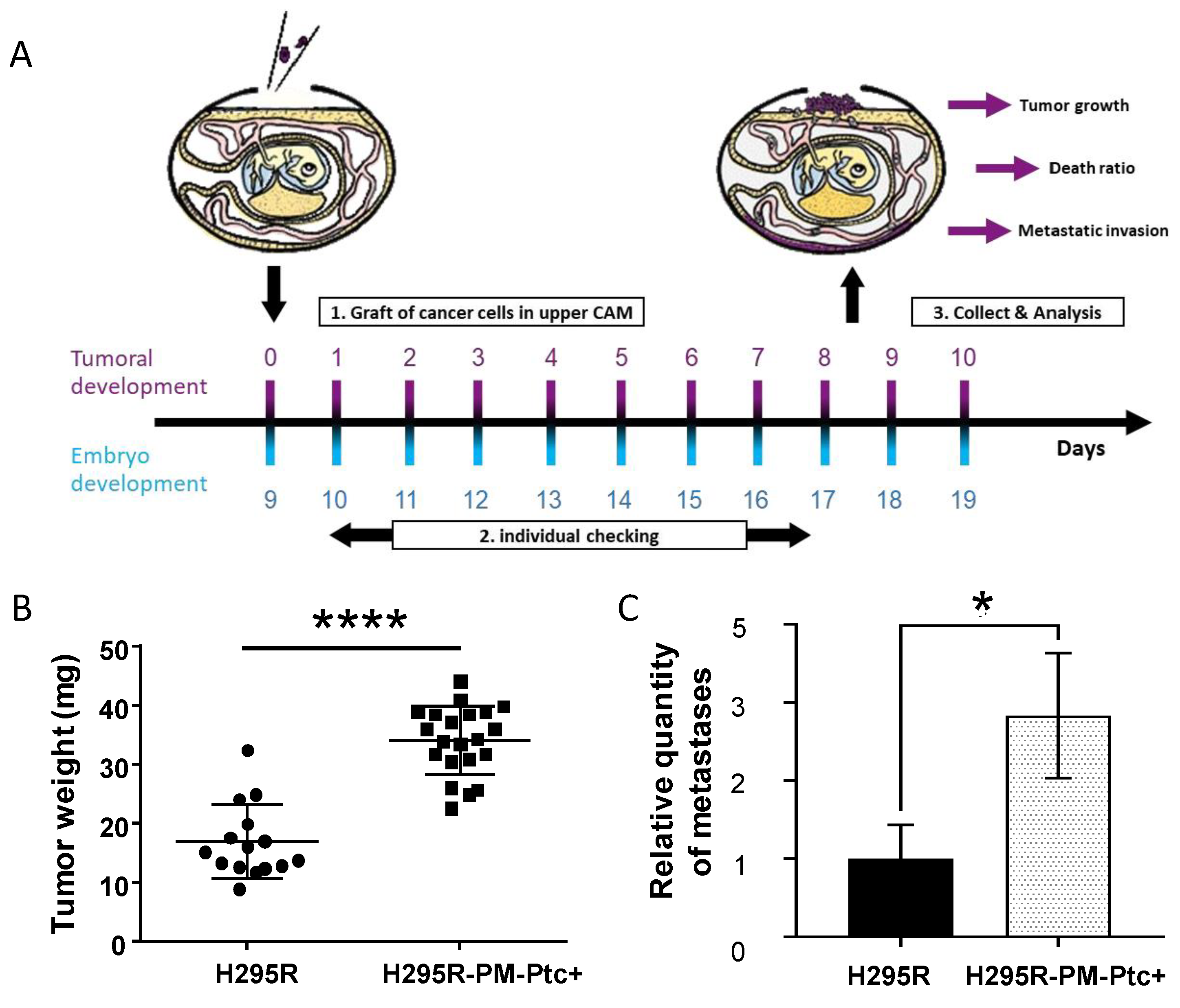
2.10. In ovo Experiment
2.11. Experiments in Mice
2.12. Immunofluorescence on Mouse Tumor Slides
2.13. Statistical Analysis
2.14. RNA-Seq
3. Results:
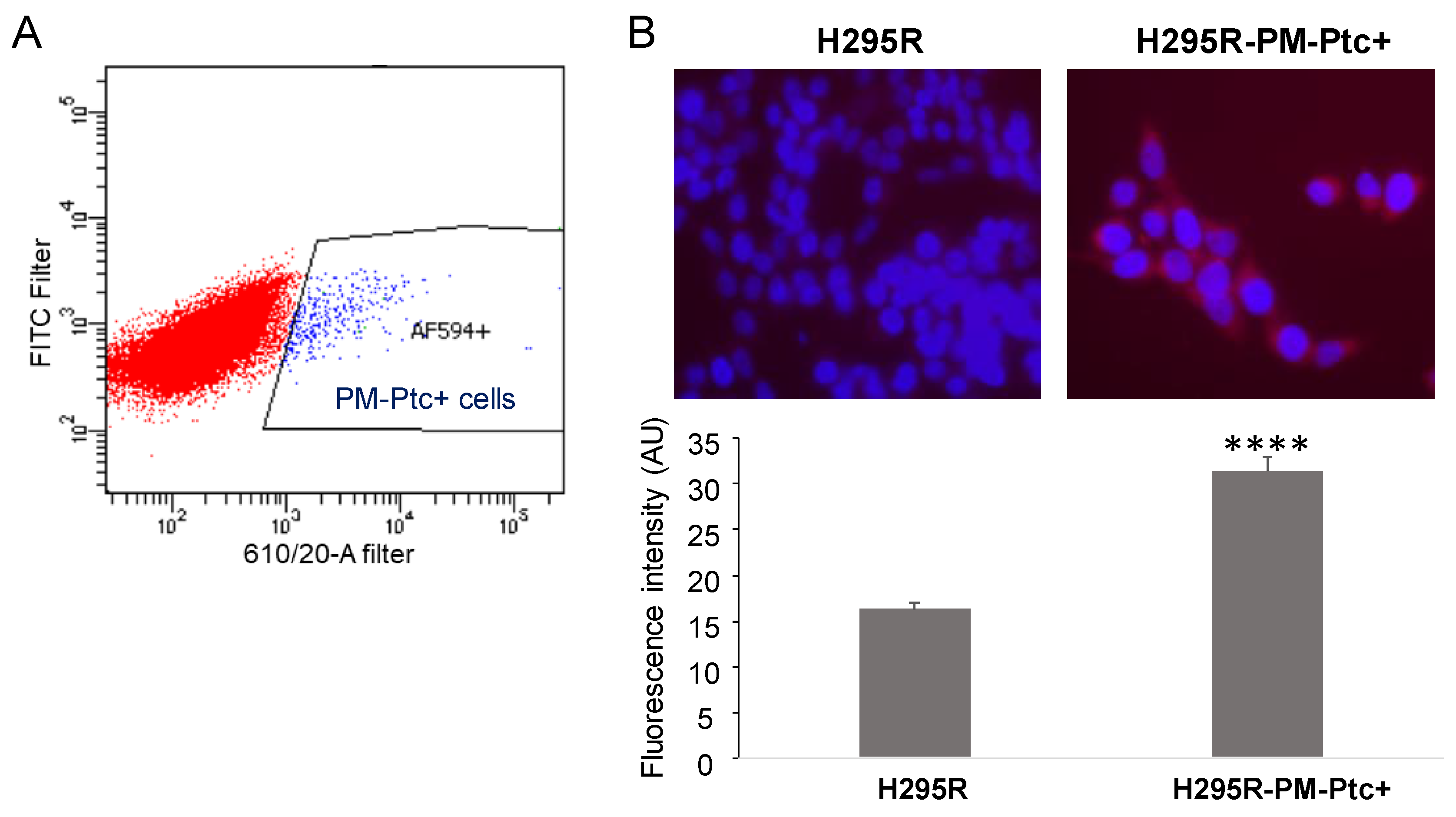
3.1. A Small Subpopulation of the ACC Cell Line H295R Presents an Increased Amount of Ptch1 at the Plasma Membrane
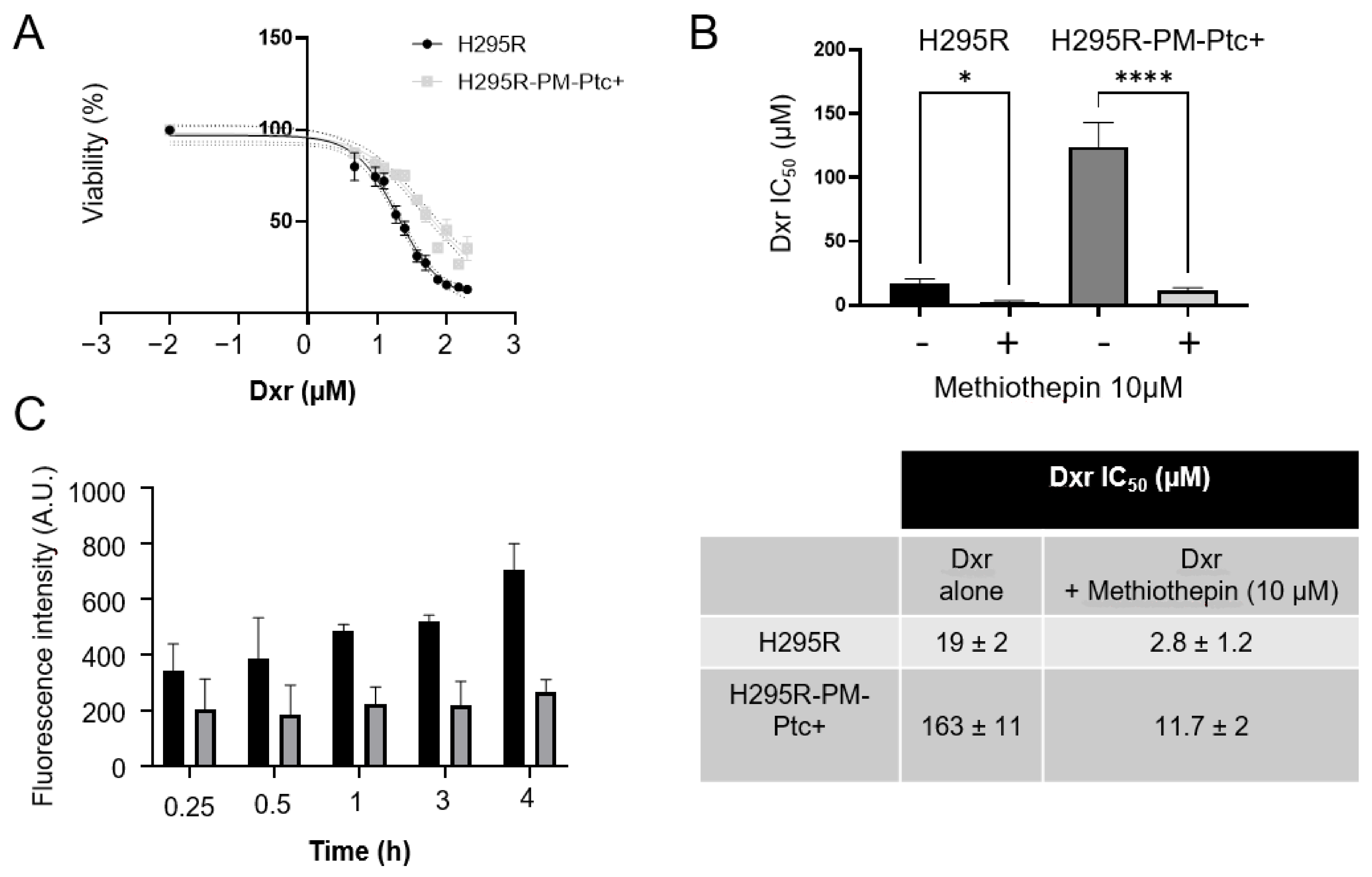
3.2. H295R-PM-Ptc+ Cells Are More Resistant to Chemotherapy Than Parental Cells
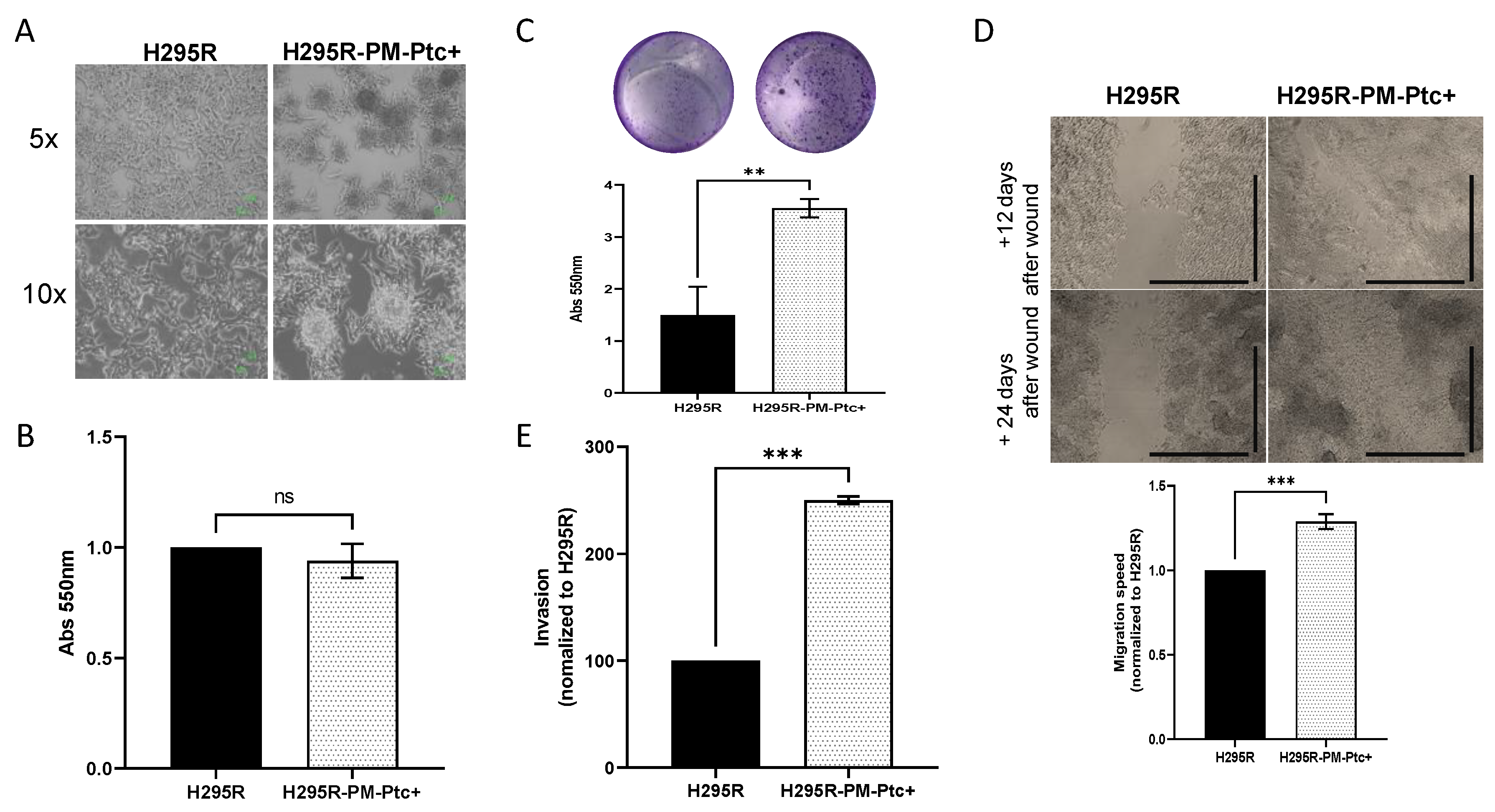
3.3. H295R-PM-Ptc+ Cells Show Superior In Vitro Clonogenic, Migratory and Invasive Capabilities Than Parental H295R Cells
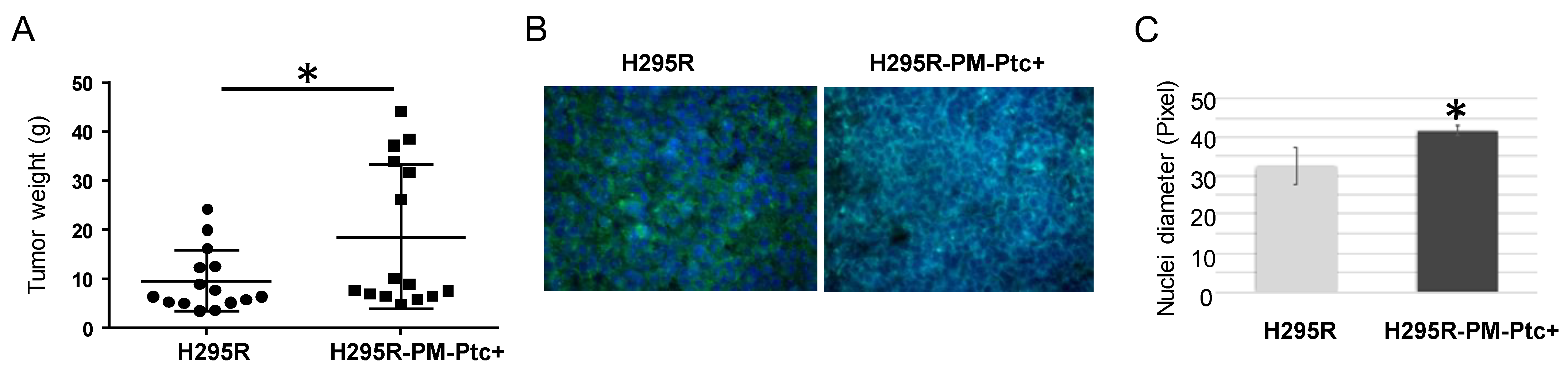
3.4. H295R-PM-Ptc+ Cells Are More Tumorigenic and Metastatic In Vivo Than Parental Cells
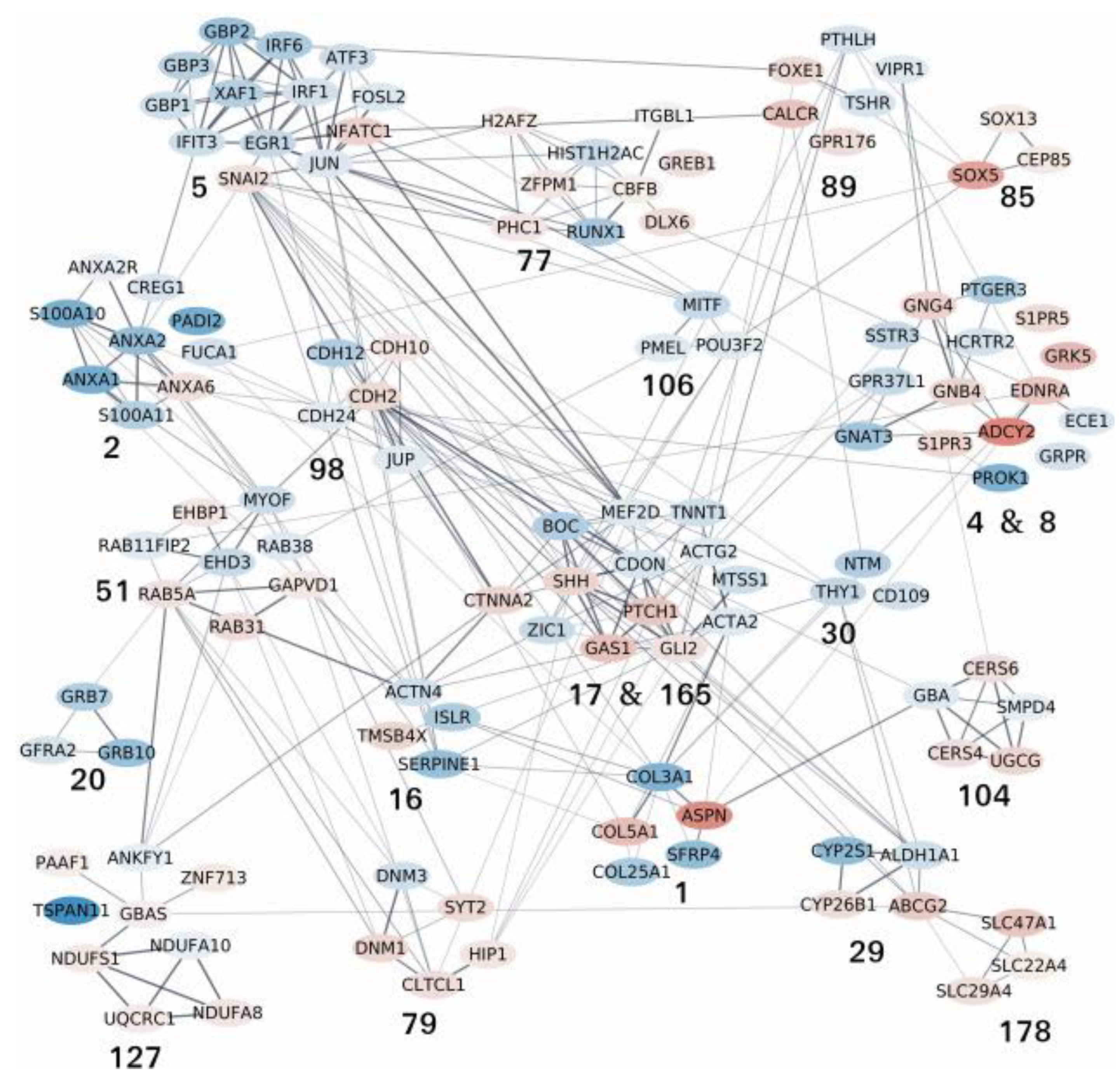
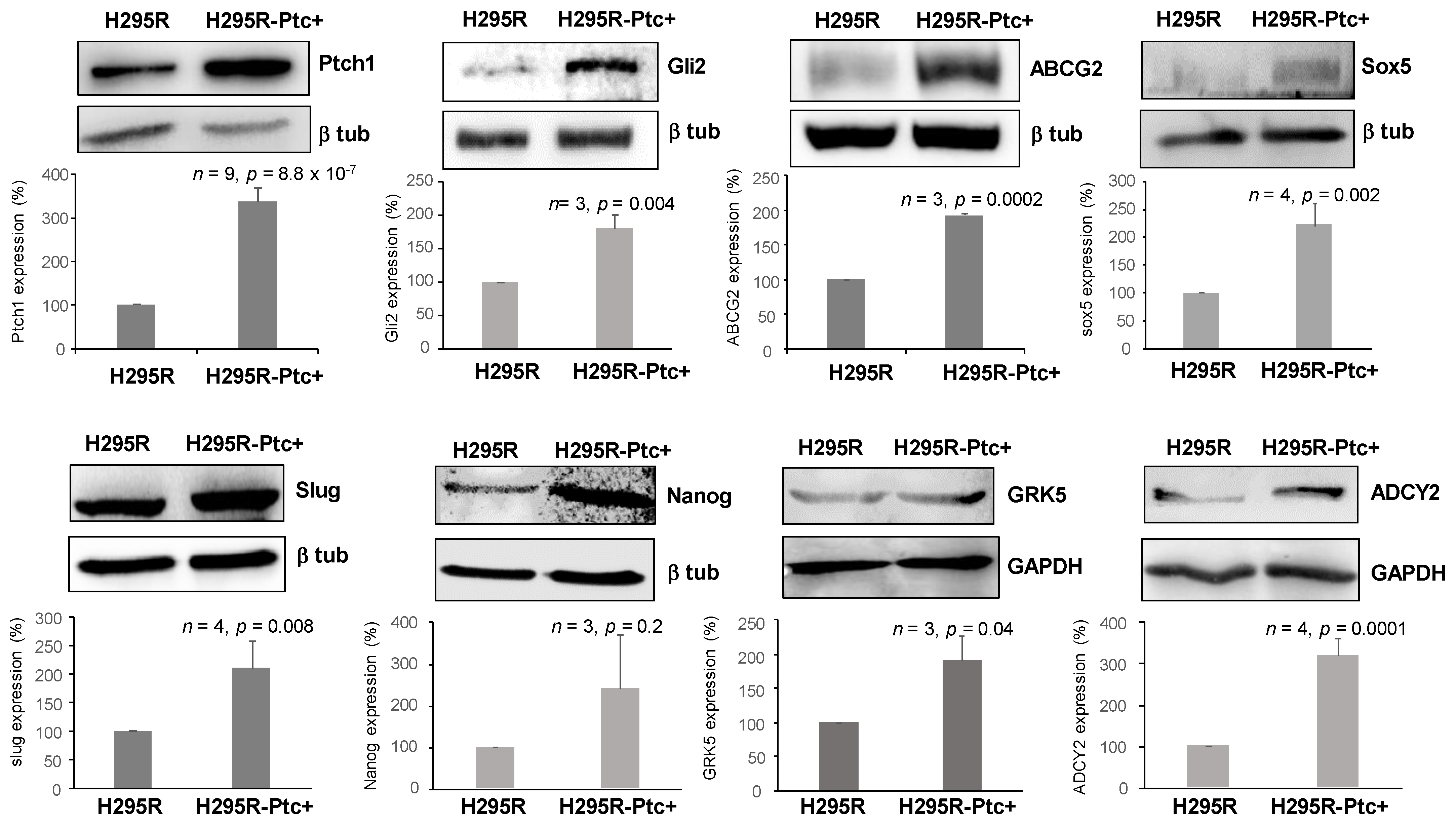
3.5. H295R-PM-Ptc+ Cells Differentially Express Genes Associated with EMC, Invasion, Metastasis and Cancer Stem Cell Properties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Andervont, H.B. Biological Aspects of Cancer. Julian Huxley. Harcourt, Brace, New York, 1958. 156 pp. $3.75. Science 1958, 127, 1440–1441. [Google Scholar] [CrossRef]
- Johnson, B.E.; Mazor, T.; Hong, C.; Barnes, M.; Aihara, K.; McLean, C.Y.; Fouse, S.D.; Yamamoto, S.; Ueda, H.; Tatsuno, K.; et al. Mutational Analysis Reveals the Origin and Therapy-Driven Evolution of Recurrent Glioma. Science 2014, 343, 189–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto-Vila, M.; Takahashi, R.-U.; Usuba, W.; Kohama, I.; Ochiya, T. Drug Resistance Driven by Cancer Stem Cells and Their Niche. Int. J. Mol. Sci. 2017, 18, 2574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikubo, M.; Inoue, Y.; Liu, G.; Tsao, M.-S. Mechanism of Drug Tolerant Persister Cancer Cells: The Landscape and Clinical Implication for Therapy. J. Thorac. Oncol. 2021, 16, 1798–1809. [Google Scholar] [CrossRef] [PubMed]
- Balzerano, A.; Paccosi, E.; Proietti-De-Santis, L. Evolutionary Mechanisms of Cancer Suggest Rational Therapeutic Approaches. Cytogenet. Genome Res. 2021, 161, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Ozben, T. Mechanisms and strategies to overcome multiple drug resistance in cancer. FEBS Lett. 2006, 580, 2903–2909. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-Q.; Yang, Y.; Cai, C.-Y.; Teng, Q.-X.; Cui, Q.; Lin, J.; Assaraf, Y.G.; Chen, Z.-S. Multidrug resistance proteins (MRPs): Structure, function and the overcoming of cancer multidrug resistance. Drug Resist. Updat. 2021, 54, 100743. [Google Scholar] [CrossRef]
- Cree, I.A.; Charlton, P. Molecular chess? Hallmarks of anti-cancer drug resistance. BMC Cancer 2017, 17, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R., Jr.; Chen, Z.-S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updat. 2015, 18, 1–17. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Scales, S.J.; de Sauvage, F.J. Mechanisms of Hedgehog pathway activation in cancer and implications for therapy. Trends Pharmacol. Sci. 2009, 30, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, C.; Szczepny, A.; Watkins, D.N.; Cain, J.E. Hedgehog Signaling in the Maintenance of Cancer Stem Cells. Cancers 2015, 7, 1554–1585. [Google Scholar] [CrossRef] [PubMed]
- Hasanovic, A.; Mus-Veteau, I. Targeting the Multidrug Transporter Ptch1 Potentiates Chemotherapy Efficiency. Cells 2018, 7, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saze, Z.; Terashima, M.; Kogure, M.; Ohsuka, F.; Suzuki, H.; Gotoh, M. Activation of the Sonic Hedgehog Pathway and Its Prognostic Impact in Patients with Gastric Cancer. Dig. Surg. 2012, 29, 115–123. [Google Scholar] [CrossRef]
- Xu, X.; Ding, H.; Rao, G.; Arora, S.; Saclarides, C.P.; Esparaz, J.; Gattuso, P.; Solorzano, C.C.; A Prinz, R. Activation of the Sonic Hedgehog pathway in thyroid neoplasms and its potential role in tumor cell proliferation. Endocr. -Relat. Cancer 2012, 19, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Bidet, M.; Tomico, A.; Martin, P.; Guizouarn, H.; Mollat, P.; Mus-Veteau, I. The Hedgehog Receptor Patched Functions in Multidrug Transport and Chemotherapy Resistance. Mol. Cancer Res. 2012, 10, 1496–1508. [Google Scholar] [CrossRef] [Green Version]
- Hasanovic, A.; Ruggiero, C.; Jung, S.; Rapa, I.; Signetti, L.; Ben Hadj, M.; Terzolo, M.; Beuschlein, F.; Volante, M.; Hantel, C.; et al. Targeting the multidrug transporter Patched potentiates chemotherapy efficiency on adrenocortical carcinoma in vitro and in vivo. Int. J. Cancer 2018, 143, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Signetti, L.; Elizarov, N.; Simsir, M.; Paquet, A.; Douguet, D.; Labbal, F.; Debayle, D.; Di Giorgio, A.; Biou, V.; Girard, C.; et al. Inhibition of Patched Drug Efflux Increases Vemurafenib Effectiveness against Resistant BrafV600E Melanoma. Cancers 2020, 12, 1500. [Google Scholar] [CrossRef]
- Lalli, E.; Luconi, M. The next step: Mechanisms driving adrenocortical carcinoma metastasis. Endocr. Relat. Cancer 2018, 25, R31–R48. [Google Scholar] [CrossRef] [Green Version]
- Uchihara, M.; Tanioka, M.; Kojima, Y.; Nishikawa, T.; Sudo, K.; Shimoi, T.; Noguchi, E.; Maeshima, A.M.; Yonemori, K. Clinical management and outcomes associated with etoposide, doxorubicin, and cisplatin plus mitotane treatment in metastatic adrenocortical carcinoma: A single institute experience. Int. J. Clin. Oncol. 2021, 26, 2275–2281. [Google Scholar] [CrossRef] [PubMed]
- Edamaghi, M.; Wojtkowiak, J.W.; Gillies, R.J. pH sensing and regulation in cancer. Front. Physiol. 2013, 4, 370. [Google Scholar] [CrossRef] [Green Version]
- Fiorini, L.; Mus-Veteau, I. Method to Screen Multidrug Transport Inhibitors Using Yeast Overexpressing a Human MDR Transporter. In Heterologous Expression of Membrane Proteins; Humana Press: New York, NY, USA, 2016; Volume 1432, pp. 303–318. [Google Scholar] [CrossRef]
- Durand, N.; Simsir, M.; Signetti, L.; Labbal, F.; Ballotti, R.; Mus-Veteau, I. Methiothepin Increases Chemotherapy Efficacy against Resistant Melanoma Cells. Molecules 2021, 26, 1867. [Google Scholar] [CrossRef] [PubMed]
- Hasanovic, A.; Simsir, M.; Choveau, F.S.; Lalli, E.; Mus-Veteau, I. Astemizole Sensitizes Adrenocortical Carcinoma Cells to Doxorubicin by Inhibiting Patched Drug Efflux Activity. Biomedicines 2020, 8, 251. [Google Scholar] [CrossRef] [PubMed]
- Fiorini, L.; Tribalat, M.-A.; Sauvard, L.; Cazareth, J.; Lalli, E.; Broutin, I.; Thomas, O.P.; Mus-Veteau, I. Natural paniceins from mediterranean sponge inhibit the multidrug resistance activity of Patched and increase chemotherapy efficiency on melanoma cells. Oncotarget 2015, 6, 22282–22297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funakoshi, K.; Bagheri, M.; Zhou, M.; Suzuki, R.; Abe, H.; Akashi, H. Highly sensitive and specific Alu-based quantification of human cells among rodent cells. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Mus-Veteau, I. Comparison of Gene Expression between Parental Adrenocortical Carcinoma Cells and a Subpopulation Overexpressing Ptch1. Available online: https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE189424 (accessed on 19 January 2022).
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2018, 18, 623–632. [Google Scholar] [CrossRef]
- Pasquier, C.; Guerlais, V.; Pallez, D.; Rapetti-Mauss, R.; Soriani, O. Identification of active modules in interaction networks using node2vec network embedding. bioRxiv 2021, 2021.09.22.461345. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Grover, A.; Leskovec, J. node2vec. KDD 2016, 2016, 855–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquier, C. Active Module Deletion with AMINE Algorithm. Available online: http://amine.i3s.unice.fr/ (accessed on 10 January 2022).
- Izzi, L.; Lévesque, M.; Morin, S.; Laniel, D.; Wilkes, B.C.; Mille, F.; Krauss, R.S.; McMahon, A.P.; Allen, B.L.; Charron, F. Boc and Gas1 Each Form Distinct Shh Receptor Complexes with Ptch1 and Are Required for Shh-Mediated Cell Proliferation. Dev. Cell 2011, 20, 788–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.-M.; Zhang, J.-G.; Zhang, X.; Li, Q. Targeting cancer stem cells for reversing therapy resistance: Mechanism, signaling, and prospective agents. Signal Transduct. Target. Ther. 2021, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Farhood, B.; Mortezaee, K. Cancer stem cells (CSCs) in cancer progression and therapy. J. Cell. Physiol. 2019, 234, 8381–8395. [Google Scholar] [CrossRef] [PubMed]
- Im, S.; Choi, H.J.; Yoo, C.; Jung, J.-H.; Jeon, Y.-W.; Suh, Y.J.; Kang, C.S. Hedgehog Related Protein Expression in Breast Cancer: Gli-2 Is Associated with Poor Overall Survival. Korean J. Pathol. 2013, 47, 116–123. [Google Scholar] [CrossRef]
- Papadopoulos, V.; Tsapakidis, K.; Galdo, N.R.-D.; Papandreou, C.N.; Del Galdo, F.; Anthoney, A.; Sakellaridis, N.; Dimas, K.; Kamposioras, K. The Prognostic Significance of the Hedgehog Signaling Pathway in Colorectal Cancer. Clin. Color. Cancer 2016, 15, 116–127. [Google Scholar] [CrossRef]
- Zhu, W.; You, Z.; Li, T.; Yu, C.; Tao, G.; Hu, M.; Chen, X. Correlation of Hedgehog Signal Activation with Chemoradio-therapy Sensitivity and Survival in Esophageal Squamous Cell Carcinomas. Jpn. J. Clin. Oncol. 2010, 41, 386–393. [Google Scholar] [CrossRef] [Green Version]
- Gonnissen, A.; Isebaert, S.; Perneel, C.; McKee, C.M.; Van Utterbeeck, F.; Lerut, E.; Verrill, C.; Bryant, R.J.; Joniau, S.; Muschel, R.J.; et al. Patched 1 Expression Correlates with Biochemical Relapse in High-Risk Prostate Cancer Patients. Am. J. Pathol. 2018, 188, 795–804. [Google Scholar] [CrossRef] [Green Version]
- Delloye-Bourgeois, C.; Gibert, B.; Rama, N.; Delcros, J.-G.; Gadot, N.; Scoazec, J.-Y.; Krauss, R.; Bernet, A.; Mehlen, P. Sonic Hedgehog Promotes Tumor Cell Survival by Inhibiting CDON Pro-Apoptotic Activity. PLOS Biol. 2013, 11, e1001623. [Google Scholar] [CrossRef] [Green Version]
- Mille, F.; Tamayo-Orrego, L.; Lévesque, M.; Remke, M.; Korshunov, A.; Cardin, J.; Bouchard, N.; Izzi, L.; Kool, M.; Northcott, P.A.; et al. The Shh Receptor Boc Promotes Progression of Early Medulloblastoma to Advanced Tumors. Dev. Cell 2014, 31, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Song, J.Y.; Holtz, A.M.; Pinskey, J.M.; Allen, B.L. Distinct structural requirements for CDON and BOC in the promotion of Hedgehog signaling. Dev. Biol. 2015, 402, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaszak, I.; Witkowska-Piłaszewicz, O.; Niewiadomska, Z.; Dworecka-Kaszak, B.; Toka, F.N.; Jurka, P. Role of Cadherins in Cancer—A Review. Int. J. Mol. Sci. 2020, 21, 7624. [Google Scholar] [CrossRef] [PubMed]
- Da, C.; Wu, K.; Yue, C.; Bai, P.; Wang, R.; Wang, G.; Zhao, M.; Lv, Y.; Hou, P. N-cadherin promotes thyroid tumorigenesis through modulating major signaling pathways. Oncotarget 2016, 8, 8131–8142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Lin, R.; Liao, X.; Li, D.; Qin, Y. Identification and verification of the molecular mechanisms and prognostic values of the cadherin gene family in gastric cancer. Sci. Rep. 2021, 11, 23674 . [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-J.; Dehghanian, S.Z.; Chan, T.-C.; He, H.-L.; Li, W.-S.; Abdollahi, S.; Chen, N.-Y.; Li, C.-F.; Shiue, Y.-L. High G protein subunit beta 4 protein level is correlated to poor prognosis of urothelial carcinoma. Med Mol. Morphol. 2021, 54, 356–367. [Google Scholar] [CrossRef]
- Zhou, W.; Gross, K.M.; Kuperwasser, C. Molecular regulation of Snai2 in development and disease. J. Cell Sci. 2019, 132, jcs235127. [Google Scholar] [CrossRef]
- You, S.; Knudsen, B.S.; Erho, N.; Alshalalfa, M.; Takhar, M.; Al-Deen Ashab, H.; Davicioni, E.; Karnes, R.J.; Klein, E.A.; Den, R.B.; et al. Integrated Classification of Prostate Cancer Reveals a Novel Luminal Subtype wth Poor Outcome. Cancer Res. 2016, 76, 4948–4958. [Google Scholar] [CrossRef] [Green Version]
- Haslehurst, A.M.; Koti, M.; Dharsee, M.; Nuin, P.; Evans, K.; Geraci, J.; Childs, T.; Chen, J.; Li, J.; Weberpals, J.; et al. EMT transcription factors snail and slug directly contribute to cisplatin resistance in ovarian cancer. BMC Cancer 2012, 12, 91. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Duan, Z.; Yu, J.; Dang, H.-X. NFATc1 regulates cell proliferation, migration, and invasion of ovarian cancer SKOV3 cells in vitro and in vivo. Oncol. Rep. 2016, 36, 918–928. [Google Scholar] [CrossRef] [Green Version]
- Britzen-Laurent, N.; Lipnik, K.; Ocker, M.; Naschberger, E.; Schellerer, V.S.; Croner, R.S.; Vieth, M.; Waldner, M.; Steinberg, P.; Hohenadl, C.; et al. GBP-1 acts as a tumor suppressor in colorectal cancer cells. Carcinogenesis 2012, 34, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Min, H.; Hu, B.; Xue, X.; Liu, Y. Guanylate-binding protein-2 inhibits colorectal cancer cell growth and increases the sensitivity to paclitaxel of paclitaxel-resistant colorectal cancer cells by interfering Wnt signaling. J. Cell. Biochem. 2019, 121, 1250–1259. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Dong, P.; Chen, H.; Xu, L.; Liu, Y.; Ma, Y.; Zheng, Y.; Yang, J.; Zhou, Y.; Chen, L.; et al. Decreased expression of ATF3, orchestrated by β-catenin/TCF3, miR-17-5p and HOXA11-AS, promoted gastric cancer progression via increased β-catenin and CEMIP. Exp. Mol. Med. 2021, 53, 1706–1722. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Chen, C.; Zheng, F.; Ni, X.; Lin, J.; Wu, W.; Lai, X. ATF3 inhibits the growth and stem cells-like features of SW620 colorectal cancer cells in vitro. J. Mens. Health 2021, 17, 287–294. [Google Scholar] [CrossRef]
- Shi, Q.; Yan, X.; Wang, J.; Zhang, X. Collagen Family Genes Associated with Risk of Recurrence after Radiation Therapy for Vestibular Schwannoma and Pan-Cancer Analysis. Dis. Markers 2021, 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, L.; Yang, H.; Li, C.; Fang, C. Identification of potential genes related to breast cancer brain metastasis in breast cancer patients. Biosci. Rep. 2021, 41, BSR20211615. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, H.; Wang, L.; Zhang, H.; Lu, J.; Liang, Z.; Liu, T. Asporin promotes pancreatic cancer cell invasion and migration by regulating the epithelial-to-mesenchymal transition (EMT) through both autocrine and paracrine mechanisms. Cancer Lett. 2017, 398, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Itoh, G.; Takagane, K.; Fukushi, Y.; Kuriyama, S.; Umakoshi, M.; Goto, A.; Yanagihara, K.; Yashiro, M.; Tanaka, M. Cancer-associated fibroblasts educate normal fibroblasts to facilitate cancer cell spreading and T-cell suppression. Mol. Oncol. 2021, 16, 166–187. [Google Scholar] [CrossRef]
- Sasaki, Y.; Takagane, K.; Konno, T.; Itoh, G.; Kuriyama, S.; Yanagihara, K.; Yashiro, M.; Yamada, S.; Murakami, S.; Tanaka, M. Expression of asporin reprograms cancer cells to acquire resistance to oxidative stress. Cancer Sci. 2021, 112, 1251–1261. [Google Scholar] [CrossRef]
- Chi, L.-H.; Chang, W.-M.; Chang, Y.-C.; Chan, Y.-C.; Tai, C.-C.; Leung, K.-W.; Chen, C.-L.; Wu, A.T.; Lai, T.-C.; Li, Y.-C.; et al. Global Proteomics-based Identification and Validation of Thymosin Beta-4 X-Linked as a Prognostic Marker for Head and Neck Squamous Cell Carcinoma. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Duerr, E.-M.; Mizukami, Y.; Ng, A.; Xavier, R.J.; Kikuchi, H.; Deshpande, V.; Warshaw, A.L.; Glickman, J.; Kulke, M.H.; Chung, D.C. Defining molecular classifications and targets in gastroenteropancreatic neuroendocrine tumors through DNA microarray analysis. Endocr. Relat. Cancer 2008, 15, 243–256. [Google Scholar] [CrossRef]
- Hirata, N.; Yamada, S.; Shoda, T.; Kurihara, M.; Sekino, Y.; Kanda, Y. Sphingosine-1-phosphate promotes expansion of cancer stem cells via S1PR3 by a ligand-independent Notch activation. Nat. Commun. 2014, 5, 4806. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yamashita, M.; Suda, T. A Novel Function of Sphingolipid Signaling via S1PR3 in Hematopoietic and Leukemic Stem Cells. Blood Cancer Discov. 2020, 2, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Watters, R.J.; Wang, H.-G.; Sung, S.-S.; Loughran, T.P.; Liu, X. Targeting sphingosine-1-phosphate receptors in cancer. Anti-Cancer Agents Med. Chem. 2011, 11, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Fu, Y.; Xu, H.; Teng, P.; Xie, Q.; Zhang, Y.; Yan, C.; Xu, Y.; Li, C.; Zhou, J.; et al. SOX5 predicts poor prognosis in lung adenocarcinoma and promotes tumor metastasis through epithelial-mesenchymal transition. Oncotarget 2017, 9, 10891–10904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, H.; Kanda, M.; Miwa, T.; Umeda, S.; Sawaki, K.; Tanaka, C.; Kobayashi, D.; Hayashi, M.; Yamada, S.; Nakayama, G.; et al. G-protein subunit gamma-4 expression has potential for detection, prediction and therapeutic targeting in liver metastasis of gastric cancer. Br. J. Cancer 2021, 125, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Laurberg, J.R.; Jensen, J.B.; Schepeler, T.; Borre, M.; Ørntoft, T.F.; Dyrskjøt, L. High expression of GEM and EDNRA is associated with metastasis and poor outcome in patients with advanced bladder cancer. BMC Cancer 2014, 14, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kashiwagi, E.; Shiota, M.; Yokomizo, A.; Itsumi, M.; Inokuchi, J.; Uchiumi, T.; Naito, S. Prostaglandin receptor EP3 mediates growth inhibitory effect of aspirin through androgen receptor and contributes to castration resistance in prostate cancer cells. Endocr. Relat. Cancer 2013, 20, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Fa, J. Dynamin 3 overexpression suppresses the proliferation, migration and invasion of cervical cancer cells. Oncol. Lett. 2021, 22, 1–8. [Google Scholar] [CrossRef]
- Hu, M.; Gu, J.; Su, W.; Zhang, Z.; Zhu, B.; Wang, Q.; Xing, C. DNM1: A Prognostic Biomarker Associated with Immune Infiltration in Colon Cancer—A Study Based on TCGA Database. BioMed Res. Int. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Rao, D.; Hyun, T.S.; Kumar, P.D.; Mizukami, I.F.; Rubin, M.A.; Lucas, P.; Sanda, M.G.; Ross, T.S. Huntingtin-interacting protein 1 is overexpressed in prostate and colon cancer and is critical for cellular survival. J. Clin. Investig. 2002, 110, 351–360. [Google Scholar] [CrossRef]
- Sheridan, M.; Ogretmen, B. The Role of Ceramide Metabolism and Signaling in the Regulation of Mitophagy and Cancer Therapy. Cancers 2021, 13, 2475. [Google Scholar] [CrossRef] [PubMed]
- Schömel, N.; Gruber, L.; Alexopoulos, S.J.; Trautmann, S.; Olzomer, E.M.; Byrne, F.L.; Hoehn, K.L.; Gurke, R.; Thomas, D.; Ferreirós, N.; et al. UGCG overexpression leads to increased glycolysis and increased oxidative phosphorylation of breast cancer cells. Sci. Rep. 2020, 10, 8182. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Cao, K.; Kato, S.; Mizutani, N.; Tanaka, K.; Arima, C.; Tai, M.C.; Nakatani, N.; Yanagisawa, K.; Takeuchi, T.; et al. CERS6 required for cell migration and metastasis in lung cancer. J. Cell. Mol. Med. 2020, 24, 11949–11959. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.R.; Kunkalla, K.; Qu, C.; Schlette, E.; Neelapu, S.S.; Samaniego, F.; Vega, F. ABCG2 is a direct transcriptional target of hedgehog signaling and involved in stroma-induced drug tolerance in diffuse large B-cell lymphoma. Oncogene 2011, 30, 4874–4886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Zhu, M.; Cheng, W.; Xing, R.; Li, W.; Zhao, M.; Xu, L.; Li, E.; Luo, G.; Lu, Y. Downregulation of 425G>A variant of calcium-binding protein S100A14 associated with poor differentiation and prognosis in gastric cancer. J. Cancer Res. Clin. Oncol. 2014, 141, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, A.; Bydoun, M.; Holloway, R.; Waisman, D. Annexin A2 Heterotetramer: Structure and Function. Int. J. Mol. Sci. 2013, 14, 6259–6305. [Google Scholar] [CrossRef] [Green Version]
- Hitchcock, J.K.; Katz, A.A.; Schäfer, G. Dynamic reciprocity: The role of annexin A2 in tissue integrity. J. Cell Commun. Signal. 2014, 8, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.R.; Fernandez, D.J.; Thornton, S.M.; Skeate, J.; Lühen, K.P.; Da Silva, D.M.; Langen, R.; Kast, W.M. Heterotetrameric annexin A2/S100A10 (A2t) is essential for oncogenic human papillomavirus trafficking and capsid disassembly, and protects virions from lysosomal degradation. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, B.; Zhang, Y.; Cao, J.; Chen, T. Silencing of the annexin II gene down-regulates the levels of S100A10, c-Myc, and plasmin and inhibits breast cancer cell proliferation and invasion. Saudi Med. J. 2010, 31, 374–381. [Google Scholar]
- Shen, D.; Nooraie, F.; Elshimali, Y.; Lonsberry, V.; He, J.; Bose, S.; Chia, D.; Seligson, D.; Chang, H.R.; Goodglick, L. Decreased expression of annexin A1 is correlated with breast cancer development and progression as determined by a tissue microarray analysis. Hum. Pathol. 2006, 37, 1583–1591. [Google Scholar] [CrossRef]
- Chua, C.E.L.; Tang, B.L. The role of the smallGTPase Rab31 in cancer. J. Cell. Mol. Med. 2014, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, X.-F.; Wu, H.-C.; Zou, S.-B.; Wang, J.-Y.; Ni, P.-H.; Chen, X.-H.; Fan, Q.-S. Rab5a overexpression promoting ovarian cancer cell proliferation may be associated with APPL1-related epidermal growth factor signaling pathway. Cancer Sci. 2010, 101, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.-S.; Yin, P.-H.; Tseng, L.-M.; Yang, C.-H.; Hsu, C.-Y.; Lee, M.-Y.; Horng, C.-F.; Chi, C.-W. Rab5A is associated with axillary lymph node metastasis in breast cancer patients. Cancer Sci. 2011, 102, 2172–2178. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhiheng, H.; Zhanhuai, W.; Qian, X.; Yue, L.; Xiaoxu, G.; Jingsun, W.; Shu, Z.; Kefeng, D. Increased RAB31 Expression in Cancer-Associated Fibroblasts Promotes Colon Cancer Progression Through HGF-MET Signaling. Front. Oncol. 2020, 10, 1747. [Google Scholar] [CrossRef]
- Hong, D.; Fritz, A.J.; Finstad, K.H.; Fitzgerald, M.P.; Weinheimer, A.; Viens, A.L.; Ramsey, J.; Stein, J.L.; Lian, J.B.; Stein, G.S. Suppression of Breast Cancer Stem Cells and Tumor Growth by the RUNX1 Transcription Factor. Mol. Cancer Res. 2018, 16, 1952–1964. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Liang, S.; Sun, F. LncRNA DLX6-AS1 Promotes Malignant Phenotype and Lymph Node Metastasis in Prostate Cancer by Inducing LARGE Methylation. Front. Oncol. 2020, 10, 1172. [Google Scholar] [CrossRef]
- Wu, Q.; Ma, J.; Meng, W.; Hui, P. DLX6-AS1 promotes cell proliferation, migration and EMT of gastric cancer through FUS-regulated MAP4K1. Cancer Biol. Ther. 2019, 21, 17–25. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Zhu, R.; Zhang, J.; Zhang, Y. DLX6-AS1 activated by H3K4me1 enhanced secondary cisplatin resistance of lung squamous cell carcinoma through modulating miR-181a-5p/miR-382-5p/CELF1 axis. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Hodgkinson, K.; Forrest, L.A.; Vuong, N.; Garson, K.; Djordjevic, B.; Vanderhyden, B.C. GREB1 is an estrogen receptor-regulated tumour promoter that is frequently expressed in ovarian cancer. Oncogene 2018, 37, 5873–5886. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Tian, J.; Zhu, S.; Sun, L.; Yu, J.; Tian, H.; Dong, Q.; Luo, Q.; Jiang, N.; Niu, Y.; et al. Sox5 contributes to prostate cancer metastasis and is a master regulator of TGF-β-induced epithelial mesenchymal transition through controlling Twist1 expression. Br. J. Cancer 2017, 118, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, L.; Qin, Y.; Liu, S.; Qiao, Y.; Wan, X.; Zeng, H.; Tang, X.; Liu, M.; Hou, Y. Effects of differential distributed-JUP on the malignancy of gastric cancer. J. Adv. Res. 2020, 28, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Ban, Y.; Wang, K.; Sun, Y.; Zhao, Z. SOX5 promotes breast cancer proliferation and invasion by transactivation of EZH2. Oncol. Lett. 2019, 17, 2754–2762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F.; Li, X.; Feng, W.; Qiao, C.; Chen, J.; Jiang, M.; Qiu, Z.; Qian, M.; Tian, D.; Nie, Y.; et al. SOX13 promotes colorectal cancer metastasis by transactivating SNAI2 and c-MET. Oncogene 2020, 39, 3522–3540. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Fang, F.; Fang, T.; You, Y.; Feng, M.; Wang, X.; Yin, Z.; Zhao, W. SOX13 regulates cancer stem-like properties and tumorigenicity in hepatocellular carcinoma cells. Am. J. Cancer Res. 2021, 11, 760–772. [Google Scholar] [PubMed]
- Yamaguchi, M.; Watanabe, Y.; Ohtani, T.; Uezumi, A.; Mikami, N.; Nakamura, M.; Sato, T.; Ikawa, M.; Hoshino, M.; Tsuchida, K.; et al. Calcitonin Receptor Signaling Inhibits Muscle Stem Cells from Escaping the Quiescent State and the Niche. Cell Rep. 2015, 13, 302–314. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, P.; Xu, S.; Li, Y.; Dekker, J.D.; Li, B.; Fan, Y.; Zhang, Z.; Hong, Y.; Yang, G.; et al. FOXP1 controls mesenchymal stem cell commitment and senescence during skeletal aging. J. Clin. Investig. 2017, 127, 1241–1253. [Google Scholar] [CrossRef]
- Simmons, J.; Pierce, C.J.; Al-Ejeh, F.; Boyle, G.M. MITF and BRN2 contribute to metastatic growth after dissemination of melanoma. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Carreira, S.; Goodall, J.; Denat, L.; Rodriguez, M.; Nuciforo, P.; Hoek, K.S.; Testori, A.; LaRue, L.; Goding, C.R. Mitf regulation of Dia1 controls melanoma proliferation and invasiveness. Genes Dev. 2006, 20, 3426–3439. [Google Scholar] [CrossRef] [Green Version]
- Travnickova, J.; Wojciechowska, S.; Khamseh, A.; Gautier, P.; Brown, D.V.; Lefevre, T.; Brombin, A.; Ewing, A.; Capper, A.; Spitzer, M.; et al. Zebrafish MITF-Low Melanoma Subtype Models Reveal Transcriptional Subclusters and MITF-Independent Residual Disease. Cancer Res. 2019, 79, 5769–5784. [Google Scholar] [CrossRef] [Green Version]
- Hoek, K.S.; Goding, C.R. Cancer stem cells versus phenotype-switching in melanoma. Pigment. Cell Melanoma Res. 2010, 23, 746–759. [Google Scholar] [CrossRef]
- Johannes, L. The Cellular and Chemical Biology of Endocytic Trafficking and Intracellular Delivery—The GL–Lect Hypothesis. Molecules 2021, 26, 3299. [Google Scholar] [CrossRef] [PubMed]
- Christenson, L. Steroidogenic acute regulatory protein (StAR) and the intramitochondrial translocation of cholester-ol. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1529, 175–187. [Google Scholar]
- Wiedenmann, B.; Franke, W.W.; Kuhn, C.; Moll, R.; E Gould, V. Synaptophysin: A marker protein for neuroendocrine cells and neoplasms. Proc. Natl. Acad. Sci. USA 1986, 83, 3500–3504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faillot, S.; Assie, G. ENDOCRINE TUMOURS: The genomics of adrenocortical tumors. Eur. J. Endocrinol. 2016, 174, R249–R265. [Google Scholar] [CrossRef] [Green Version]
- Brondani, V.; Lacombe, A.; Mariani, B.; Montenegro, L.; Soares, I.; Bezerra-Neto, J.; Tanno, F.; Srougi, V.; Chambo, J.; Mendonca, B.; et al. Low Protein Expression of both ATRX and ZNRF3 as Novel Negative Prognostic Markers of Adult Adrenocortical Carcinoma. Int. J. Mol. Sci. 2021, 22, 1238. [Google Scholar] [CrossRef]
- Bai, X.; Fisher, D.E.; Flaherty, K.T. Cell-state dynamics and therapeutic resistance in melanoma from the perspective of MITF and IFNγ pathways. Nat. Rev. Clin. Oncol. 2019, 16, 549–562. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GDE | H295R-PMPtc+/H295R Log2 | p-Value | Role in Cancer |
|---|---|---|---|
| Hedgehog (Hh) Signaling Pathway | |||
| PTCH1 | +0.629 | 1.08 × 10−6 | Hh target gene. drug efflux. chemotherapy resistance |
| GAS1 | +0.724 | 4.52 × 10−8 | Hh pathway activation |
| SHH | +0.421 | 2.82 × 10−3 | |
| BOC | −0.794 | 3.25 × 10−7 | BOC endocytosis via NUMB is required for Ptch1 internalization and Shh signaling |
| HDAC9 | −0.925 | 6.59 × 10−17 | Negative regulator of Hh signaling |
| GPR37L1 | −0.548 | 1.27 × 10−4 | Interacts with Ptch1 for Hh signaling modulation |
| NUMBL | −0.286 | 1.47 × 10−2 | Negative regulator of canonical Shh signaling. Required for BOC and Ptch1 internalization |
| Tumor Suppressor | |||
| DNM3 | −0.427 | 1.21 × 10−4 | Decreased expression is associated with worse prognosis |
| RUNX1 | −0.898 | 9.00 × 10−11 | Represses breast CSC phenotype through direct inhibition of Zeb1/2. Low RunX1 expression is associated with poor patient survival |
| CSC Maintenance | |||
| UGCG | +0.418 | 2.70 × 10−7 | CSC maintenance and chemotherapy resistance. increase antiapoptotic gene expression |
| SOX5 | +1.149 | 7.70 × 10−17 | Overexpressed in CSC in lung cancer, promotes migration, invasion and metastasis. Predicts poor prognosis |
| NANOGP1 | +0.424 | 6.42 × 10−4 | Self-renewal transcription factor, maintenance of CSC, EMT and metastasis. |
| GATA6 | +0.444 | 1.18 × 10−4 | Maintenance of stem cell phenotype, regulated by Nanog |
| KLF7 | +0.456 | 9.06 × 10−4 | Upregulated in quiescent cells Restores hematopoietic stem cells niche |
| COL5A1 | +0.754 | 4.58 × 10−9 | Quiescence and self-renewal of stem cells |
| CALCR | +0.696 | 1.90 10−6 | Maintains cells in a quiescent state |
| MITF | −0.564 | 1.58 × 10−10 | Low MITF is associated to slow cycling and senescence phenotype, dedifferentiation and treatment resistance |
| GRB10 | −1.104 | 5.20 × 10−16 | Deletion promotes hematopoietic stem cell self-renewal and regeneration |
| CSC and Chemotherapy Resistance | |||
| ABCG2 | +0.592 | 1.39 × 10−4 | Drug efflux. Resistance to chemotherapy |
| ABCC4 | +0.352 | 1.94 × 10−2 | Chemotherapy resistance. CSC marker |
| SLC47A1 | +0.667 | 6.46 × 10−8 | SLC transporters in chemotherapy resistance |
| SLC2A4 | +0.429 | 9.32 × 10−4 | Energy production of cancer cells, migration, metastasis |
| SLC22A3 | +0.339 | 4.10 × 10−3 | Cell invasion and filopodia formation, metastasis |
| ID2 | +0.294 | 1.08 × 10−3 | Inhibition of differentiation and maintenance of self-renewal and multipotency in stem cells |
| EMT | |||
| TMSB4X | +0.434 | 6.13 × 10−5 | Suppresses E-cadherin expression, promotes cancer cell growth and migration |
| SNAI2 | +0.295 | 1.36 × 10−3 | Triggers EMT, blocks cell cycle and confers resistance to cell death |
| RAB5A | +0.256 | 8.33 × 10−4 | Migration and metastasis |
| ASPN | +1.476 | 2.51 × 10−22 | Promotes cell migration and invasion |
| CDH2 | +0.390 | 2.63 × 10−3 | Mesenchymal marker |
| CD109 | −0.425 | 6.50 × 10−3 | Co-receptor of TGFB1 Inversely correlates with EMT, migration and invasion |
| TGFBI | −1.75 | 2.83 × 10−29 | TGFB1 deficiency predisposes mice to tumor development |
| Metastasis, Cancer Cell Survival | |||
| ADCY2 | +1.56 | 5.63 × 10−24 | Involved in metastasis. High expression related to worse survival. |
| DNM1 | +0.452 | 1.05 × 10−4 | Invasion and metastasis |
| GRK5 | +0.831 | 3.84 × 10−9 | Tumor progression |
| Endocytosis | |||
| S100A10 | −1.473 | 3.86 × 10−21 | Undifferentiated phenotype and poor prognostic-low level in metastatic melanoma |
| ANXA2 | −1.144 | 8.08 × 10−16 | ANXA2 and S100A10 heterotetramer is involved in regulation of endocytosis. |
| ST6GAL1 | +0.668 | 1.54 × 10−10 | Inhibition of endocytosis GL-Lect dependent. High tumor grade, metastasis and reduced patient prognosis |
| CERS6 | +0.314 | 5.55 × 10−3 | |
| Adrenocortical Carcinoma Markers | |||
| STAR | −0.289 | 4.21 × 10−2 | Lower expression this steroidogenic enzymes is associated with a less differentiated phenotype. |
| ZNRF3 | −0.315 | 3.21 × 10−3 | Low expression: negative prognostic marker of ACC |
| SYP | −0.453 | 3.11 × 10−6 | Differentiation marker in tumor diagnosis |
| Module Number and Composition | Representative Enrichment | Role of the DEGs in Cancer |
|---|---|---|
| 1 ASPN, COL5A1, COL25A1, COL3A1, SFRP4 | Extracellular matrix organization | CSC maintenance, EMT |
| 2 ANXA6,ANXA1, ANXA2, ANXA2R, GREG1, FUCA1, PADi2, S100A10, S100A11 | Vesicle transport, cell adhesion | Cancer development and progression, poorer prognosis |
| 4 and 8 ADCY2, S1PR3, S1PR5, EDNRA, GNB4, GNG4, GRK5,GNAT3, GPR37L1, PTGER3, SSTR3, ECE1, GRPR, HCRTR2, PROK1 | Signaling by GPCR | Regulation of CSC, cancer cell survival, metastasis, poor prognosis |
| 5 NFATC1, SNAI2, ATF3, EGR1, FOSL2, GBP1, GBP2, GBP3, IFIT3, IRF1, IRF6, JUN, XAF1 | Regulation of cell population proliferation | Maintenance of mesenchymal phenotype, invasive migration, metastasis, poor prognosis, chemotherapy resistance |
| 16 TMSB4X, ACTN4, ISLR, SERPINE1 | Platelet degranulation | EMT induction and metastasis |
| 17 and 165 PTCH1, SHH, GLI2, CTNNA2, GAS1, BOC, CDON, MEF2D, TNNT1, ZIC1, ACTA2, ACTG2, MTSS1 | Hedgehog signaling, cell differentiation, cell fate specification, development | Hedgehog pathway activation, CSC maintenance, tumor cell survival |
| 20 GFRA2, GRB10, GRB7 | RET signaling | Stem cell self-renewal |
| 29 ABCG2, CYP26B1ALDH1A1, CYP2S1 | Retinoic acid metabolic process | Chemotherapy resistance, CSC, poor prognosis |
| 51 EHBP1, GAPVD1, RAB5A, RAB31, EHD3, MYOF, RAB11, FIP2, RAB38 | Endosome membrane, endocytosis | Membrane trafficking, endocytosis, cancer cell motility and invasiveness, cancer development and progression, metastasis |
| 77 DLX6, GREB1, HIST1, H2AC, CBFB,H2AFZ, ITGBL1, PHC1, RUNX1, ZFPM1 | Negative regulation of differentiation | Poor patient survival, malignant phenotype and metastasis, tumor progression and development, cell proliferation, migration and EMT |
| 79 CLTCL1, DNM1, HIP1, SYT2,DNM3 | Endocytosis, membrane trafficking | Proliferation, migration and invasion, cancer progression and metastasis, poor prognosis |
| 85 CEP85 SOX13 SOX5 | Transcription factors, cell-cycle progression | CSC maintenance, EMT, invasion, metastasis |
| 89 CALCR, FOXE1, GPR176,PTHLH, TSHR, VIPR1 | G protein-coupled receptor activity | Stem cell maintenance |
| 98 CDH10, CDH2, CDH12, CDH24, JUP | Cell–cell adhesion | EMT, cell migration and invasion, poor prognosis |
| 104 CERS4, CERS6, UGCG,GBA, SMPD4 | Sphingolipid metabolism | Multidrug resistance, proliferation of cancer cells, cell migration and metastasis |
| 106 MITF, PMEL, POU3F2 | Melanoma phenotype switching | Enrichment of stem cells, de-differentiated state and invasiveness |
| 127 GBAS, NDUFA8 NDUFS1 PAAF1 UQCRC1 ZNF713,ANKFY1, NDUFA10 TSPAN11 | Mitochondrial respiratory chain complex | Malignant behavior of cancer cells, cell cycle progression |
| 178 SLC22A4 SLC29A4 SLC47A1 | SLC-mediated transmembrane transport | Cell proliferation, epithelial-to-mesenchymal transition |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feliz Morel, Á.J.; Hasanovic, A.; Morin, A.; Prunier, C.; Magnone, V.; Lebrigand, K.; Aouad, A.; Cogoluegnes, S.; Favier, J.; Pasquier, C.; et al. Persistent Properties of a Subpopulation of Cancer Cells Overexpressing the Hedgehog Receptor Patched. Pharmaceutics 2022, 14, 988. https://doi.org/10.3390/pharmaceutics14050988
Feliz Morel ÁJ, Hasanovic A, Morin A, Prunier C, Magnone V, Lebrigand K, Aouad A, Cogoluegnes S, Favier J, Pasquier C, et al. Persistent Properties of a Subpopulation of Cancer Cells Overexpressing the Hedgehog Receptor Patched. Pharmaceutics. 2022; 14(5):988. https://doi.org/10.3390/pharmaceutics14050988
Chicago/Turabian StyleFeliz Morel, Álvaro Javier, Anida Hasanovic, Aurélie Morin, Chloé Prunier, Virginie Magnone, Kevin Lebrigand, Amaury Aouad, Sarah Cogoluegnes, Judith Favier, Claude Pasquier, and et al. 2022. "Persistent Properties of a Subpopulation of Cancer Cells Overexpressing the Hedgehog Receptor Patched" Pharmaceutics 14, no. 5: 988. https://doi.org/10.3390/pharmaceutics14050988