


Biomimetic Magnetic Particles for the Removal of Gram-Positive Bacteria and Lipoteichoic Acid
 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Synthesis, Functionalization, and Physicochemical Characterization of SPION-APTES
2.2.2. Bacteria Culture Conditions and Preparation
2.2.3. Bacterial Agglutination and Turbidometric Assays
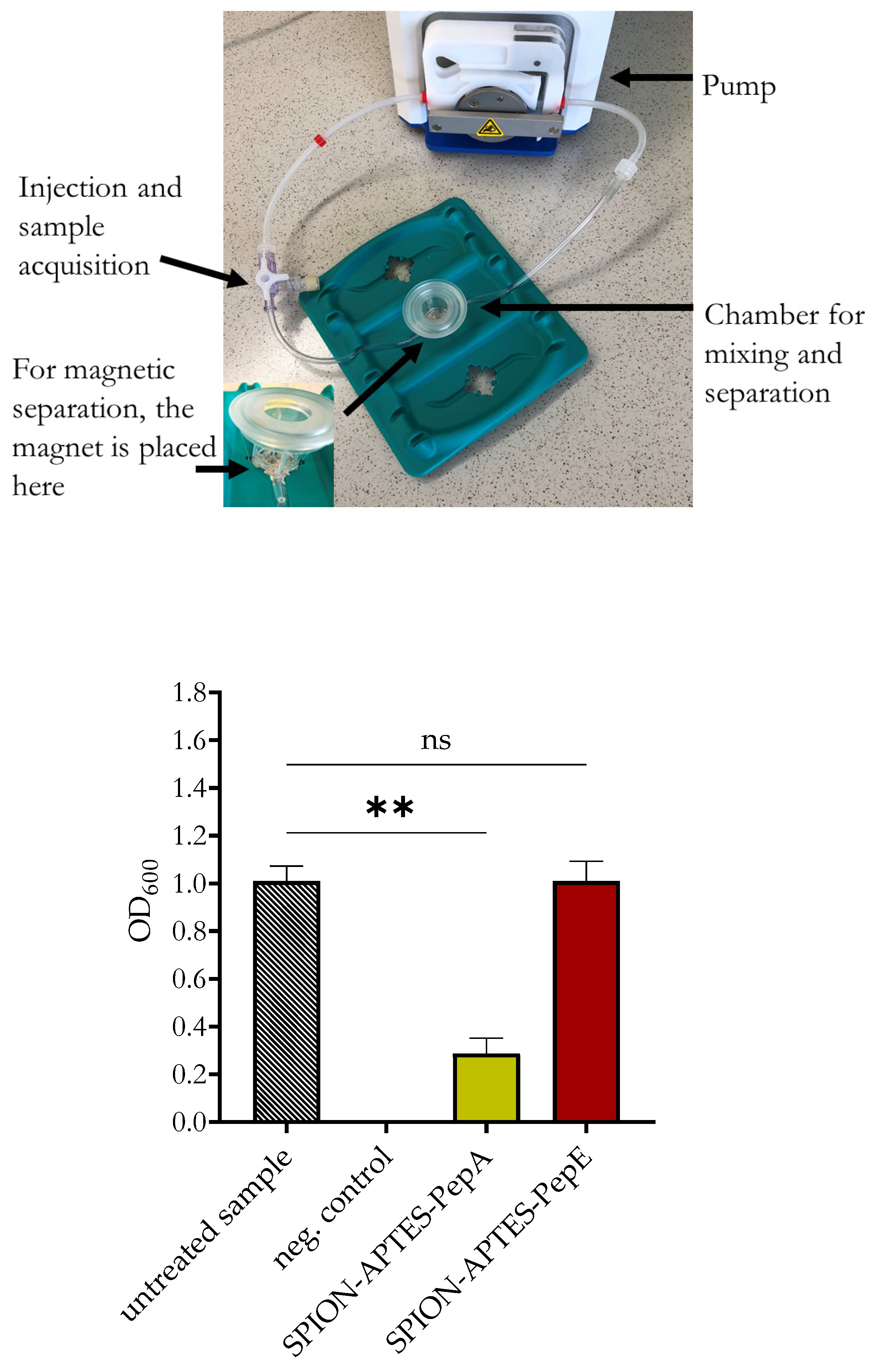
2.2.4. Bacteria Separation from Media by Peptide-Functionalized SPION-APTES
2.2.5. Separation Efficiency of LTA by Peptide-Functionalized SPION-APTES
2.3. Statistics
3. Results
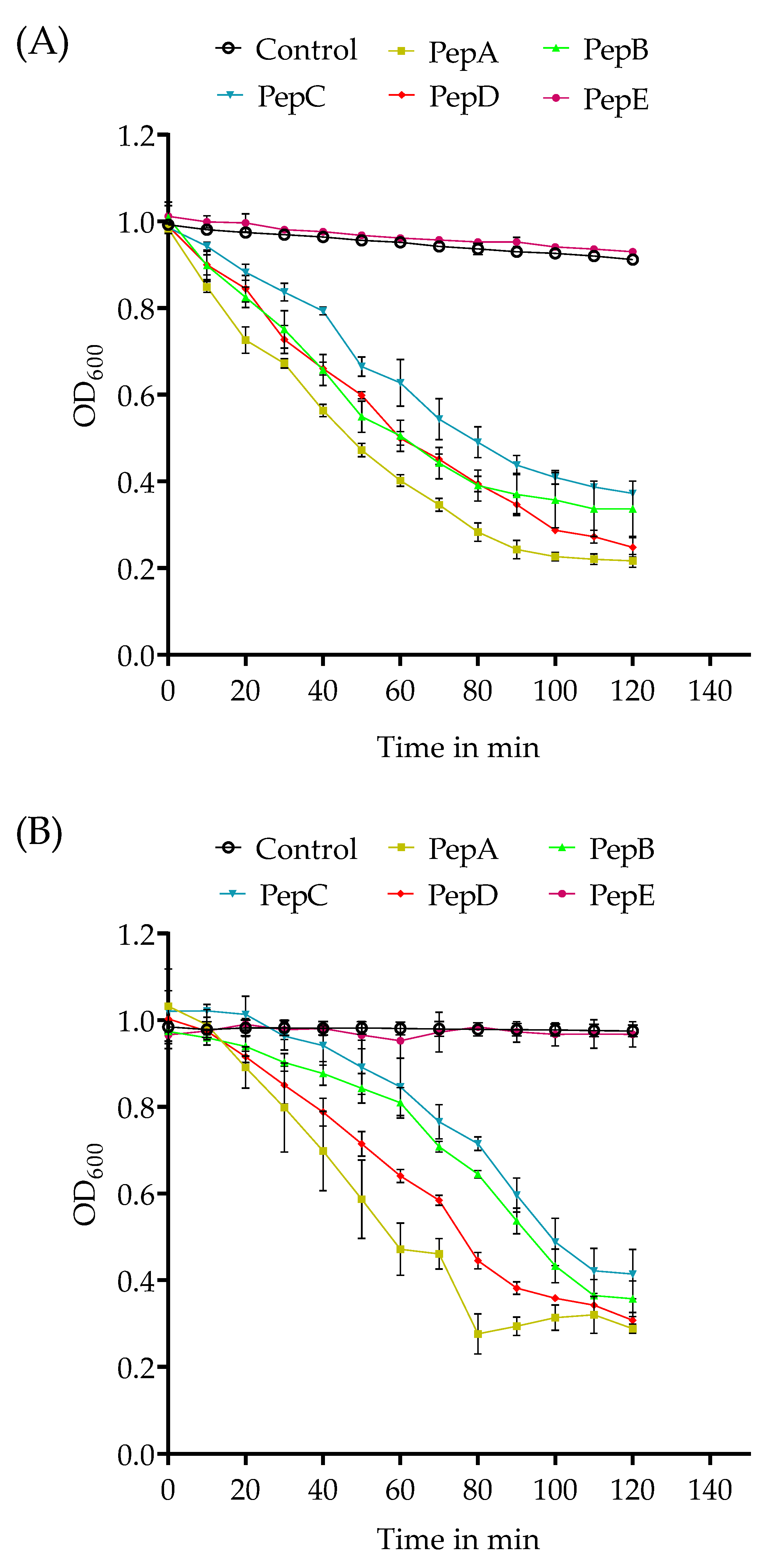
3.1. Agglutination of Gram-Positive Bacteria
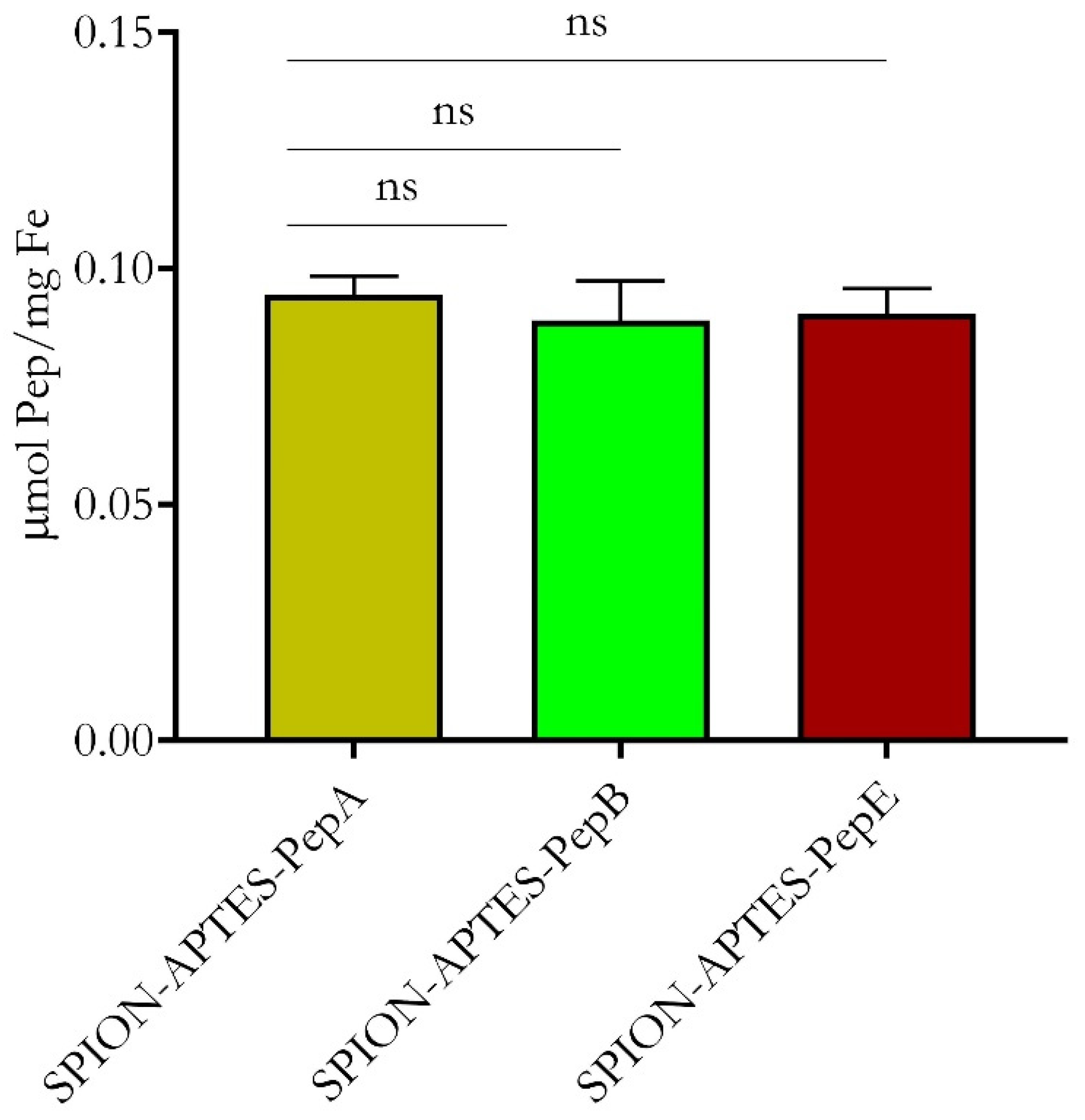
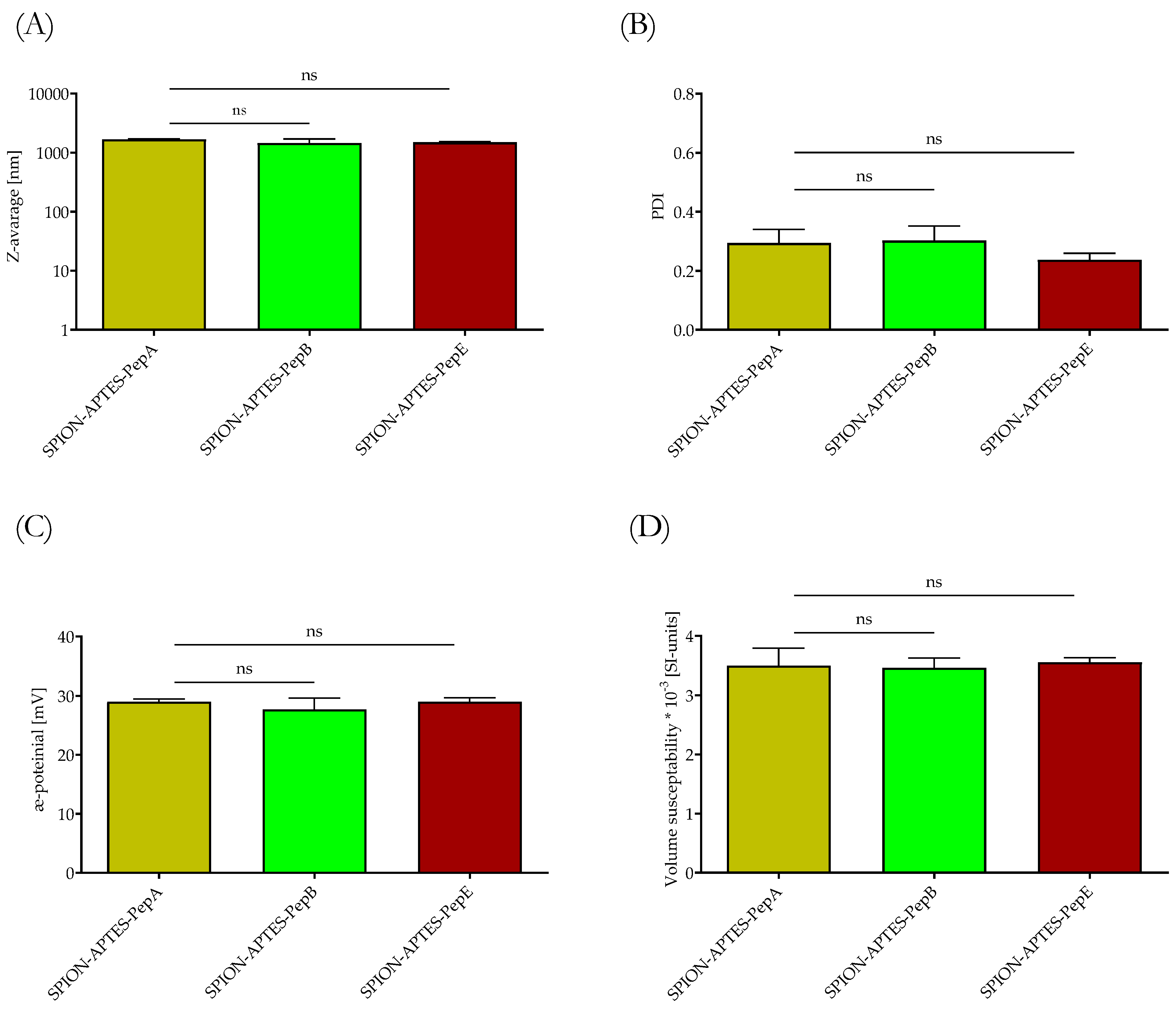
3.2. Physicochemical Characterization of Peptide-Functionalized SPIONs
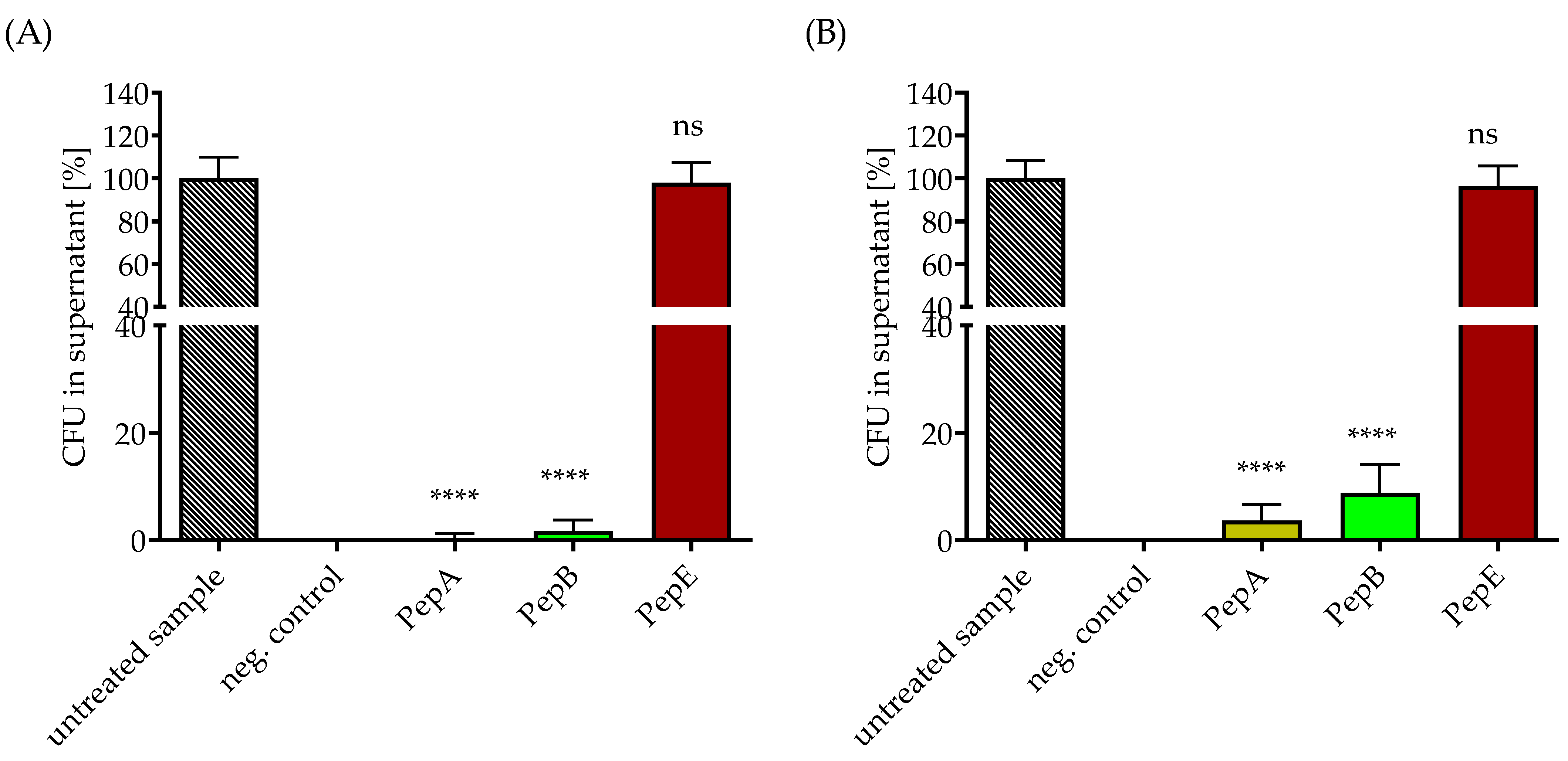
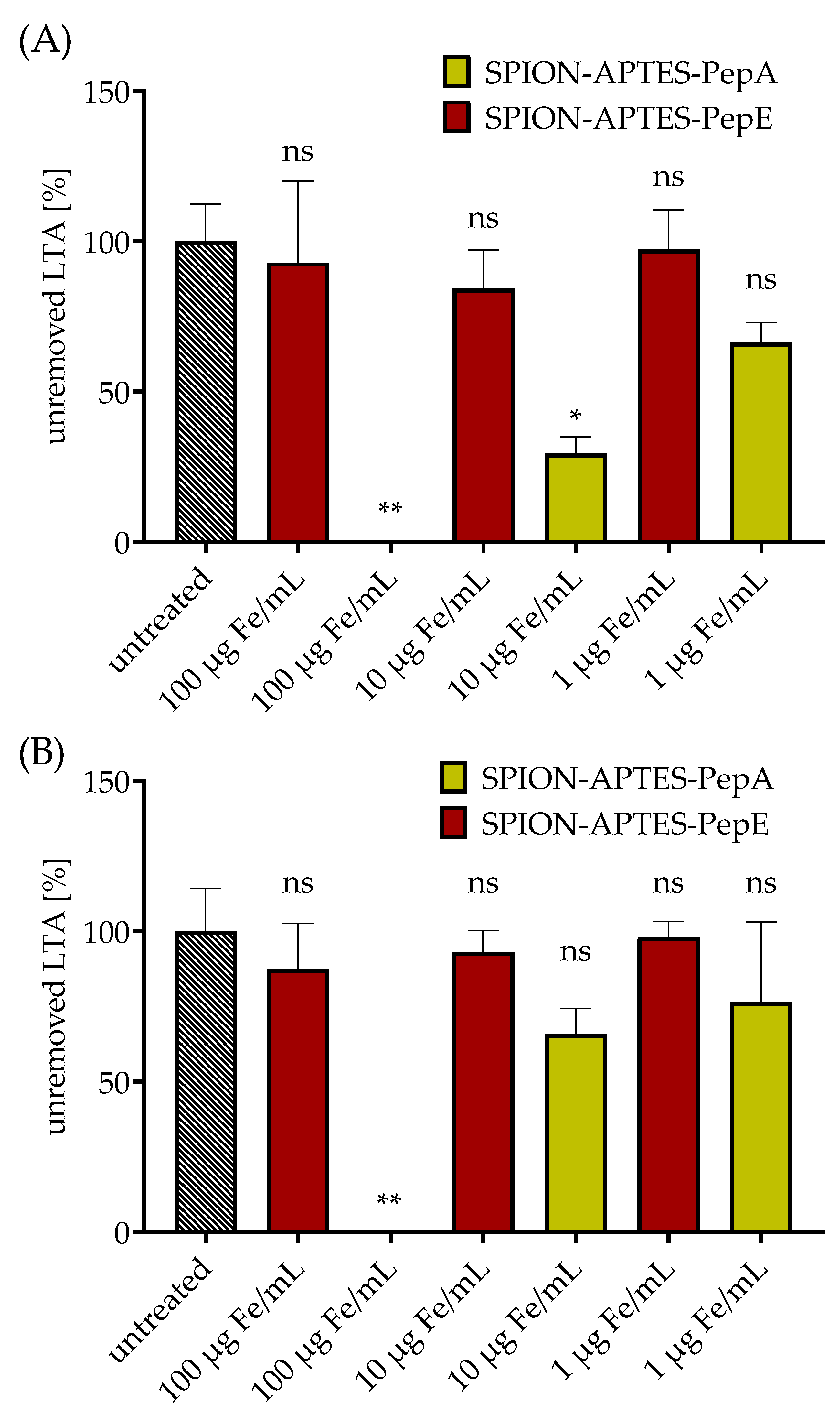
3.3. Removal of Gram-Positive Bacteria from Media under Static and Flow Conditions by Peptide-Functionalized SPION-APTES
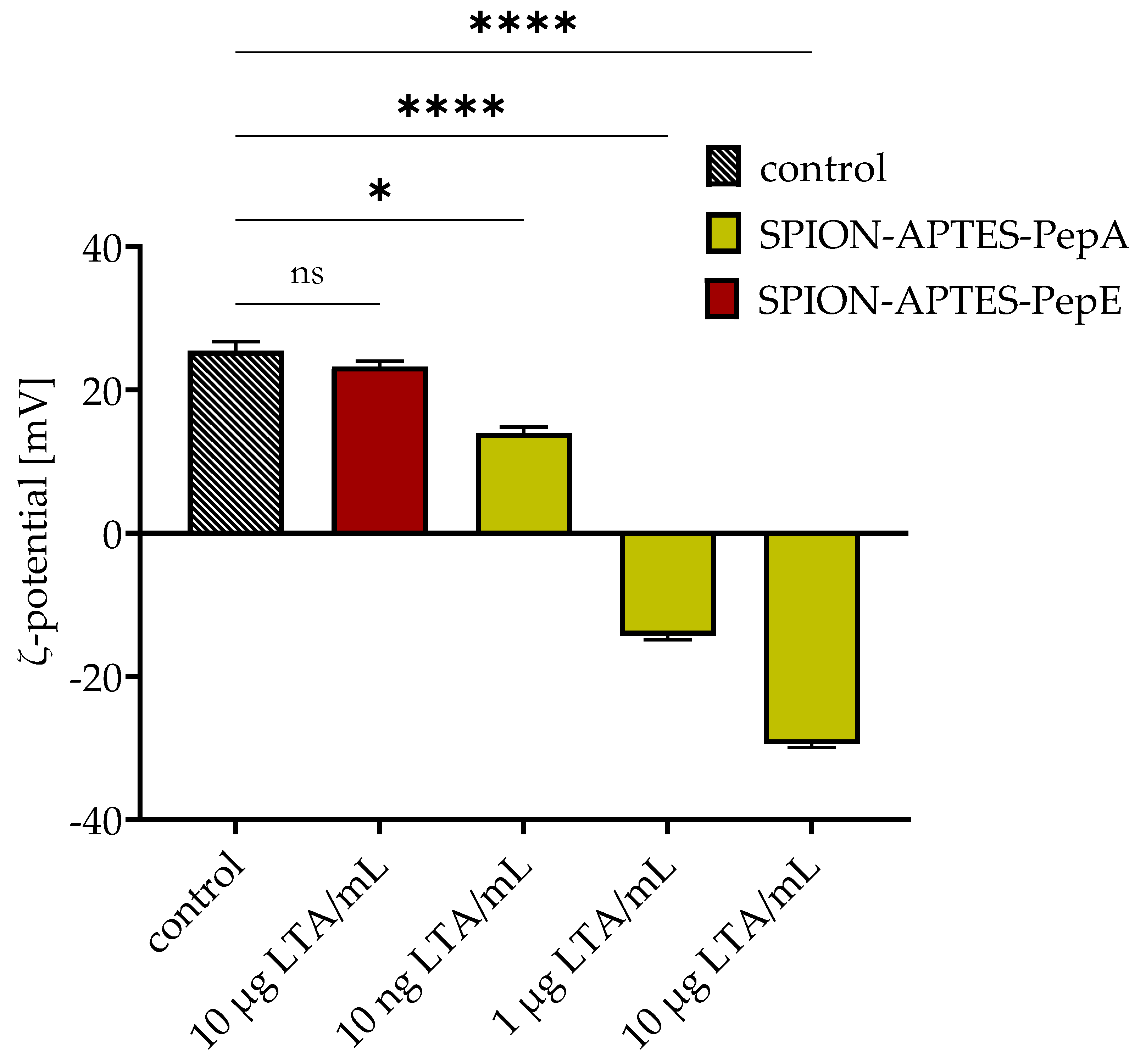
3.4. Removal of LTA by Peptide-Functionalized SPION-APTES
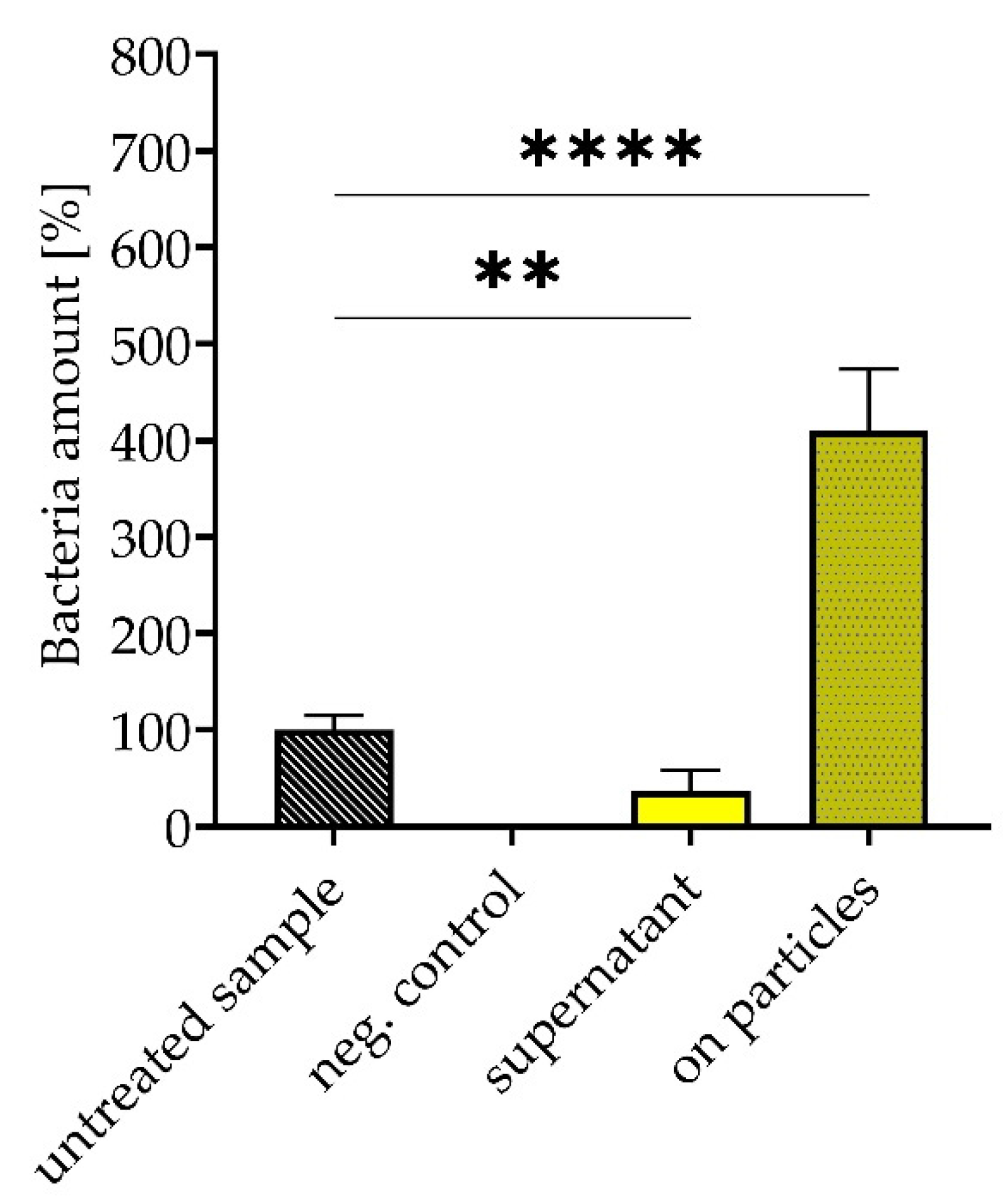
3.5. Removal and Enrichment of Separated Bacteria from Platelet Concentrate
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Ballot, D.E.; Nana, T.; Sriruttan, C.; Cooper, P.A. Bacterial Bloodstream Infections in Neonates in a Developing Country. ISRN Pediatr. 2012, 2012, 508512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, L.B. Antimicrobial resistance in gram-positive bacteria. Am. J. Infect. Control. 2006, 34 (Suppl. 5), S11–S19. [Google Scholar] [CrossRef]
- Veerachamy, S.; Yarlagadda, T.; Manivasagam, G.; Yarlagadda, P.K.D.V. Bacterial adherence and biofilm formation on medical implants: A review. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2014, 228, 1083–1099. [Google Scholar] [CrossRef] [Green Version]
- Blajchman, M.A.; Goldman, M. Bacterial contamination of platelet concentrates: Incidence, significance, and prevention. Semin. Hematol. 2001, 38, 20–26. [Google Scholar] [CrossRef]
- Schöneweck, F.; Schmitz, R.P.; Rißner, F.; Scherag, A.; Löffler, B.; Pletz, M.W.; Weis, S.; Brunkhorst, F.M.; Hagel, S. The epidemiology of bloodstream infections and antimicrobial susceptibility patterns in Thuringia, Germany: A five-year prospective, state-wide surveillance study (AlertsNet). Antimicrob. Resist. Infect. Control. 2021, 10, 132. [Google Scholar] [CrossRef]
- Sizar, O.; Unakal, C.G. Gram Positive Bacteria, in StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2021. [Google Scholar]
- Karthikeyan, G.; Premkumar, K. Neonatal Sepsis: Staphylococcus aureus as the predominant pathogen. Indian J. Pediatr. 2001, 68, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, P.C. Antimicrobial Resistance in Streptococcus pneumoniae: An Overview. Clin. Infect. Dis. 1992, 15, 77–83. [Google Scholar] [CrossRef]
- ižman, M. The use and resistance to antibiotics in the community. Int. J. Antimicrob. Agents 2003, 21, 297–307. [Google Scholar] [CrossRef]
- Yoshimura, A.; Lien, E.; Ingalls, R.; Tuomanen, E.; Dziarski, R.; Golenbock, D. Cutting edge: Recognition of Gram-Positive bacterial cell wall components by the innate immune system occurs via Toll-like receptor 2. J. Immunol. 1999, 163, 1–5. [Google Scholar]
- Vollmer, W.; Massidda, O.; Tomasz, A. The Cell Wall of Streptococcus pneumoniae. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Rajagopal, M.; Walker, S. Envelope structures of Gram-Positive bacteria. In Protein and Sugar Export and Assembly in Gram-positive Bacteria; Springer: Cham, Switzerland, 2015; pp. 1–44. [Google Scholar]
- Lotz, S.; Aga, E.; Wilde, I.; van Zandbergen, G.; Hartung, T.; Solbach, W.; Laskay, T. Highly purified lipoteichoic acid activates neutrophil granulocytes and delays their spontaneous apoptosis via CD14 and TLR2. J. Leukoc. Biol. 2003, 75, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; Fang, L.; Li, S.; Liu, R. NLRC5 negatively regulates LTA-induced inflammation via TLR2/NF-κB and participates in TLR2-mediated allergic airway inflammation. J. Cell. Physiol. 2019, 234, 19990–20001. [Google Scholar] [CrossRef] [PubMed]
- Schwandner, R.; Dziarski, R.; Wesche, H.; Rothe, M.; Kirschning, C.J. Peptidoglycan- and Lipoteichoic Acid-induced Cell Activation Is Mediated by Toll-like Receptor 2. J. Biol. Chem. 1999, 274, 17406–17409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aulock, S.V.; Deininger, S.; Draing, C.; Gueinzius, K.; Dehus, O.; Hermann, C. Gender difference in cytokine secretion on immune stimulation with LPS and LTA. J. Interferon Cytokine Res. 2006, 26, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Triantafilou, M.; Mouratis, M.-A.; Lepper, P.M.; Haston, R.M.; Baldwin, F.; Lowes, S.; Ahmed, M.A.E.; Schumann, C.; Boyd, O.; Triantafilou, K. Serum proteins modulate lipopolysaccharide and lipoteichoic acid-induced activation and contribute to the clinical outcome of sepsis. Virulence 2012, 3, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, B.; Lyer, S.; Janko, C.; Unterweger, H.; Brox, R.; Cunningham, S.; Dutz, S.; Taccardi, N.; Bikker, F.J.; Hurle, K.; et al. Scavenging of bacteria or bacterial products by magnetic particles functionalized with a broad-spectrum pathogen recognition receptor motif offers diagnostic and therapeutic applications. Acta Biomater. 2022, 141, 418–428. [Google Scholar] [CrossRef]
- Chouhan, R.; Horvat, M.; Ahmed, J.; Alhokbany, N.; Alshehri, S.; Gandhi, S. Magnetic Nanoparticles—A Multifunctional Potential Agent for Diagnosis and Therapy. Cancers 2021, 13, 2213. [Google Scholar] [CrossRef]
- Poller, J.M.; Zaloga, J.; Schreiber, E.; Unterweger, H.; Janko, C.; Radon, P.; Eberbeck, D.; Trahms, L.; Alexiou, C.; Friedrich, R.P. Selection of potential iron oxide nanoparticles for breast cancer treatment based on in vitro cytotoxicity and cellular uptake. Int. J. Nanomed. 2017, ume 12, 3207–3220. [Google Scholar] [CrossRef] [Green Version]
- Janko, C.; Ratschker, T.; Nguyen, K.; Zschiesche, L.; Tietze, R.; Lyer, S.; Alexiou, C. Functionalized Superparamagnetic Iron Oxide Nanoparticles (SPIONs) as Platform for the Targeted Multimodal Tumor Therapy. Front. Oncol. 2019, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Zaloga, J.; Janko, C.; Agarwal, R.; Nowak, J.; Müller, R.; Boccaccini, A.R.; Lee, G.; Odenbach, S.; Lyer, S.; Alexiou, C. Different Storage Conditions Influence Biocompatibility and Physicochemical Properties of Iron Oxide Nanoparticles. Int. J. Mol. Sci. 2015, 16, 9368–9384. [Google Scholar] [CrossRef]
- Mühlberger, M.; Unterweger, H.; Band, J.; Lehmann, C.; Heger, L.; Dudziak, D.; Alexiou, C.; Lee, G.; Janko, C. Loading of Primary Human T Lymphocytes with Citrate-Coated Superparamagnetic Iron Oxide Nanoparticles Does Not Impair Their Activation after Polyclonal Stimulation. Cells 2020, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Leito, J.T.; Ligtenberg, A.J.; Nazmi, K.; de Blieck-Hogervorst, J.M.; Veerman, E.C.; Nieuw Amerongen, A.V. A common binding motif for various bacteria of the bacteria-binding peptide SRCRP2 of DMBT1/gp-340/salivary agglutinin. Biol. Chem. 2008, 389, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Bikker, F.J.; Ligtenberg, A.J.; Nazmi, K.; Veerman, E.C.; van’t Hof, W.; Bolscher, J.G.; Poustka, A.; Amerongen, A.V.N.; Mollenhauer, J. Identification of the bacteria-binding peptide domain on salivary agglutinin (gp-340/DMBT1), a member of the scavenger receptor cysteine-rich superfamily. J. Biol. Chem. 2002, 277, 32109–32115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yewle, J.N.; Wei, Y.; Puleo, D.A.; Daunert, S.; Bachas, L.G. Oriented Immobilization of Proteins on Hydroxyapatite Surface Using Bifunctional Bisphosphonates as Linkers. Biomacromolecules 2012, 13, 1742–1749. [Google Scholar] [CrossRef] [PubMed]
- Yewle, J.N.; Puleo, D.A.; Bachas, L.G. Enhanced Affinity Bifunctional Bisphosphonates for Targeted Delivery of Therapeutic Agents to Bone. Bioconjugate Chem. 2011, 22, 2496–2506. [Google Scholar] [CrossRef] [Green Version]
- Bikker, F.J.; Ligtenberg, A.J.M.; End, C.; Renner, M.; Blaich, S.; Lyer, S.; Wittig, R.; Hof, W.V.; Veerman, E.C.I.; Nazmi, K.; et al. Bacteria Binding by DMBT1/SAG/gp-340 Is Confined to the VEVLXXXXW Motif in Its Scavenger Receptor Cysteine-rich Domains. J. Biol. Chem. 2004, 279, 47699–47703. [Google Scholar] [CrossRef] [Green Version]
- End, C.; Bikker, F.; Renner, M.; Bergmann, G.; Lyer, S.; Blaich, S.; Hudler, M.; Helmke, B.; Gassler, N.; Autschbach, F.; et al. DMBT1 functions as pattern-recognition molecule for poly-sulfated and poly-phosphorylated ligands. Eur. J. Immunol. 2009, 39, 833–842. [Google Scholar] [CrossRef]
- Boekhorst, P.T.; Beckers, E.; Vos, M.; Vermeij, H.; Van Rhenen, D. Clinical significance of bacteriologic screening in platelet concentrates. Transfusion 2005, 45, 514–519. [Google Scholar] [CrossRef]
- Draing, C.; Pfitzenmaier, M.; Zummo, S.; Mancuso, G.; Geyer, A.; Hartung, T.; von Aulock, S. Comparison of Lipoteichoic Acid from Different Serotypes of Streptococcus pneumoniae. J. Biol. Chem. 2006, 281, 33849–33859. [Google Scholar] [CrossRef] [Green Version]
- Morath, S.; von Aulock, S.; Hartung, T. Structure/function relationships of lipoteichoic acids. J. Endotoxin Res. 2005, 11, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Fischer, W. Lipoteichoic acid and lipids in the membrane of Staphylococcus aureus. Med. Microbiol. Immunol. 1994, 183, 61–76. [Google Scholar] [CrossRef]
- Wu, Y.-D.; Chen, L.-H.; Wu, X.-J.; Shang, S.-Q.; Lou, J.-T.; Du, L.-Z.; Zhao, Z.-Y. Gram Stain-Specific-Probe-Based Real-Time PCR for Diagnosis and Discrimination of Bacterial Neonatal Sepsis. J. Clin. Microbiol. 2008, 46, 2613–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoudi, M.; Sant, S.; Wang, B.; Laurent, S.; Sen, T. Superparamagnetic iron oxide nanoparticles (SPIONs): Development, surface modification and applications in chemotherapy. Adv. Drug Deliv. Rev. 2011, 63, 24–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Zafar, H.; Zia, M.; ul Haq, I.; Phull, A.R.; Ali, J.S.; Hussain, A. Synthesis, characterization, applications, and challenges of iron oxide nanoparticles. Nanotechnol. Sci. Appl. 2016, 9, 49–67. [Google Scholar] [CrossRef] [Green Version]
- Loos, M.; Clas, F.; Fischer, W. Interaction of purified lipoteichoic acid with the classical complement pathway. Infect. Immun. 1986, 53, 595–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtney, H.S.; A Simpson, W.; Beachey, E.H. Relationship of critical micelle concentrations of bacterial lipoteichoic acids to biological activities. Infect. Immun. 1986, 51, 414–418. [Google Scholar] [CrossRef] [Green Version]
- Bharatiya, B.; Wang, G.; Rogers, S.E.; Pedersen, J.S.; Mann, S.; Briscoe, W.H. Mixed liposomes containing gram-positive bacteria lipids: Lipoteichoic acid (LTA) induced structural changes. Colloids Surf. B Biointerfaces 2021, 199, 111551. [Google Scholar] [CrossRef]
- Olsson, J.; Glantz, P.-O. Effect of pH and counter ions on the zeta-potential of oral streptococci. Arch. Oral Biol. 1977, 22, 461–466. [Google Scholar] [CrossRef]
- Ryan, S.; Kell, A.J.; van Faassen, H.; Tay, L.-L.; Simard, B.; MacKenzie, R.; Gilbert, M.; Tanha, J. Single-Domain Antibody-Nanoparticles: Promising Architectures for Increased Staphylococcus aureus Detection Specificity and Sensitivity. Bioconjugate Chem. 2009, 20, 1966–1974. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedrich, B.; Eichermüller, J.; Bogdan, C.; Cunningham, S.; Hackstein, H.; Strauß, R.; Alexiou, C.; Lyer, S.; Tietze, R. Biomimetic Magnetic Particles for the Removal of Gram-Positive Bacteria and Lipoteichoic Acid. Pharmaceutics 2022, 14, 2356. https://doi.org/10.3390/pharmaceutics14112356
Friedrich B, Eichermüller J, Bogdan C, Cunningham S, Hackstein H, Strauß R, Alexiou C, Lyer S, Tietze R. Biomimetic Magnetic Particles for the Removal of Gram-Positive Bacteria and Lipoteichoic Acid. Pharmaceutics. 2022; 14(11):2356. https://doi.org/10.3390/pharmaceutics14112356
Chicago/Turabian StyleFriedrich, Bernhard, Julia Eichermüller, Christian Bogdan, Sarah Cunningham, Holger Hackstein, Richard Strauß, Christoph Alexiou, Stefan Lyer, and Rainer Tietze. 2022. "Biomimetic Magnetic Particles for the Removal of Gram-Positive Bacteria and Lipoteichoic Acid" Pharmaceutics 14, no. 11: 2356. https://doi.org/10.3390/pharmaceutics14112356