
Trans-cinnamaldehyde–Biosurfactant Complex as a Potent Agent against Enterococcus faecalis Biofilms
 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. Chemicals
2.3. Synthesis of TC–ASL Complex
2.4. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM) of TC–ASL
2.5. Time-Kill Kinetics and Membrane Disruption
2.6. Biofilm Formation and Treatment Assay
2.7. Quantification of Viable Biofilm Cell Count
2.8. Biomass Quantification
2.9. Confocal Laser-Scanning Microscopic (CLSM) Analysis
2.10. Statistical Analysis
3. Results
3.1. TC–ASL Forms Spherical to Ellipsoidal Micelles
3.2. TC–ASL Complex Rapidly Arrests Planktonic Bacterial Growth and Disrupts the Bacterial Cell Membrane
3.3. TC–ASL Complex Kills E. faecalis Biofilm Cells
3.4. TC–ASL Complex Reduces the Biomass of E. faecalis Biofilm
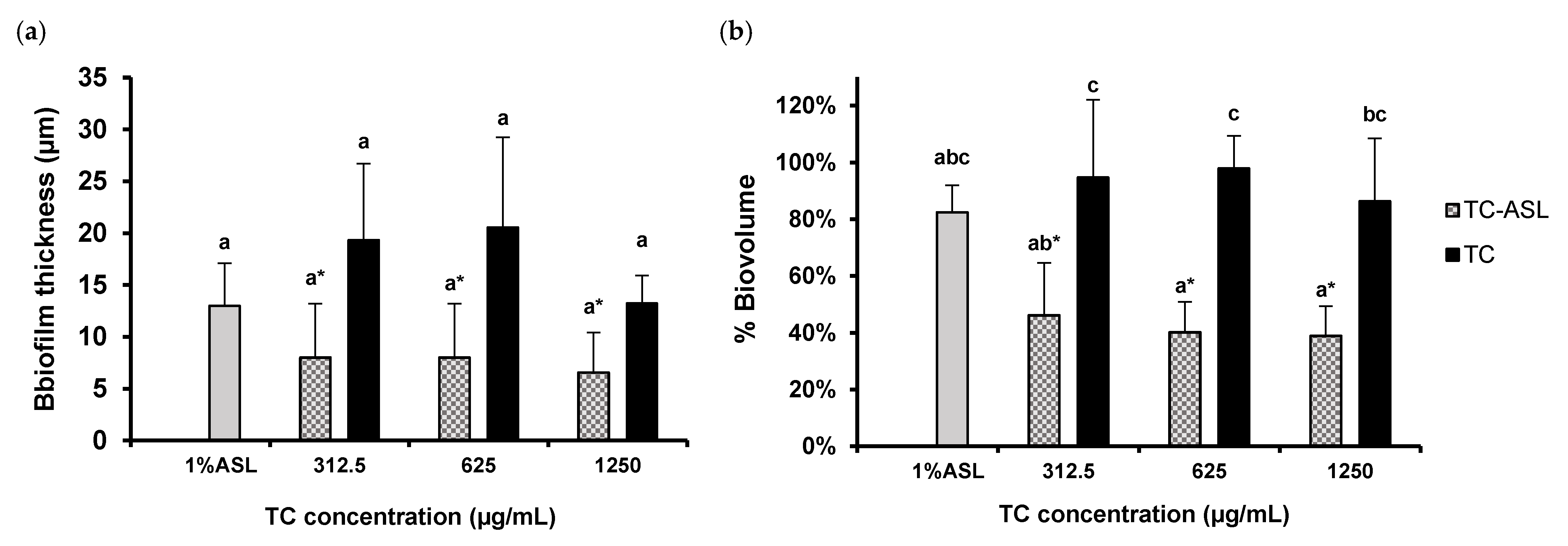
3.5. TC–ASL Reduces Biofilm Thickness and Alters the Biofilm Architecture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ch’ng, J.H.; Chong, K.K.L.; Lam, L.N.; Wong, J.J.; Kline, K.A. Biofilm-associated infection by Enterococci. Nat. Rev. Microbiol. 2019, 17, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.A.; Murray, B.E. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conceição, N.; Rodrigues, W.F.; de Oliveira, K.; da Silva, L.; de Souza, L.; da de Cunha Hueb Barata Oliveira, C.; de Oliveira, A.G. Beta-lactams susceptibility testing of penicillin-resistant, ampicillin-susceptible Enterococcus faecalis isolates: A comparative assessment of Etest and disk diffusion methods against broth dilution. Ann. Clin. Microbiol. Antimicrob. 2020, 19, 43. [Google Scholar] [CrossRef]
- García-Solache, M.; and Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, P.; Kline, K.A. Dr. Jekyll and Mr. Hide: How Enterococcus faecalis subverts the host immune response to cause infection. J. Mol. Biol. 2019, 431, 2932–2945. [Google Scholar] [CrossRef]
- Mohamed, J.A.; Huang, D.B. Biofilm formation by Enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [Green Version]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm matrixome: Extracellular components in structured microbial communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Cook, L.C.; Dunny, G.M. Effects of biofilm growth on plasmid copy number and expression of antibiotic resistance genes in Enterococcus faecalis. Antimicrob. Agents Chemother. 2013, 57, 1850–1856. [Google Scholar] [CrossRef] [Green Version]
- Holmberg, A.; Rasmussen, M. Mature biofilms of Enterococcus faecalis and Enterococcus faecium are highly resistant to antibiotics. Diagn. Microbiol. Infect. Dis. 2016, 84, 19–21. [Google Scholar] [CrossRef]
- Thieme, L.; Klinger-Strobel, M.; Hartung, A.; Stein, C.; Makarewicz, O.; Pletz, M.W. In vitro synergism and anti-biofilm activity of ampicillin, gentamicin, ceftaroline and ceftriaxone against Enterococcus faecalis. J. Antimicrob. Chemother. 2018, 73, 1553–1561. [Google Scholar] [CrossRef]
- Moellering, R.C.; Weinberg, A.N. Studies on antibiotic synergism against enterococci. II. Effect of various antibiotics on the uptake of 14C-labelled streptomycin by enterococci. J. Clin. Investig. 1971, 50, 2580–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.V.; Weinstock, G.M.; Murray, B.E. An Enterococcus faecalis ABC homologue (Lsa) is required for the resistance of this species to clindamycin and quinupristin-dalfopristin. Antimicrob. Agents Chemother. 2002, 46, 1845–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneviratne, C.J.; Suriyanarayanan, T.; Swarup, S.; Chia, K.H.B.; Nagarajan, N.; Zhang, C. Transcriptomics analysis reveals putative genes involved in biofilm formation and biofilm-associated drug resistance of Enterococcus faecalis. J. Endod. 2017, 43, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Panda, A.K.; de Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control biofilm-forming pathogens. Front Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Ali, I.A.A.; Neelakantan, P. Antibiofilm activity of phytochemicals against Enterococcus faecalis: A literature review. Phytother. Res. 2022, 36, 2824–2838. [Google Scholar] [CrossRef]
- Khare, T.; Anand, U.; Dey, A.; Assaraf, Y.G.; Chen, Z.S.; Liu, Z.; Kumar, V. Exploring phytochemicals for combating antibiotic resistance in microbial pathogens. Front. Pharmacol. 2021, 12, 720726. [Google Scholar] [CrossRef]
- Doyle, A.A.; Stephens, J.C. A review of cinnamaldehyde and its derivatives as antibacterial agents. Fitoterapia 2019, 139, 104405. [Google Scholar] [CrossRef]
- Jia, P.; Xue, Y.J.; Duan, X.J.; Shao, S.H. Effect of cinnamaldehyde on biofilm formation and sarA expression by methicillin-resistant Staphylococcus aureus. Lett. Appl. Microbiol. 2011, 53, 409–416. [Google Scholar] [CrossRef]
- Topa, S.H.; Subramoni, S.; Palombo, E.A.; Kingshott, P.; Rice, S.A.; Blackall, L.L. Cinnamaldehyde disrupts biofilm formation and swarming motility of Pseudomonas aeruginosa. Microbiology 2018, 164, 1087–1097. [Google Scholar] [CrossRef]
- Balasubramanian, A.R.; Vasudevan, S.; Shanmugam, K.; Lévesque, C.M.; Solomon, A.P.; Neelakantan, P. Combinatorial effects of trans-cinnamaldehyde with fluoride and chlorhexidine on Streptococcus mutans. J. Appl. Microbiol. 2021, 130, 382–393. [Google Scholar] [CrossRef]
- Ali, I.A.A.; Cheung, B.P.K.; Matinlinna, J.; Lévesque, C.M.; Neelakantan, P. Trans-cinnamaldehyde potently kills Enterococcus faecalis biofilm cells and prevents biofilm recovery. Microb. Pathog. 2020, 149, 104482. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.A.A.; Matinlinna, J.P.; Lévesque, C.M.; Neelakantan, P. Trans-Cinnamaldehyde attenuates Enterococcus faecalis virulence and inhibits biofilm formation. Antibiotics 2021, 10, 702. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Tian, L.L.; Lei, Q.; Zhang, Y.L. Physical stability and antimicrobial activity of encapsulated cinnamaldehyde by self-emulsifying nanoemulsion. J. Food Process Eng. 2015, 39, 462–471. [Google Scholar]
- Gafur, A.; Sukamdani, G.Y.; Kristi, N.; Maruf, A.; Xu, J.; Chen, X.; Wang, G.; Ye, Z. From bulk to nano-delivery of essential phytochemicals: Recent progress and strategies for antibacterial resistance. J. Mater. Chem. B 2020, 8, 9825–9835. [Google Scholar] [CrossRef]
- Naughton, P.J.; Marchant, R.; Naughton, V.; Banat, I.M. Microbial biosurfactants: Current trends and applications in agricultural and biomedical industries. J. Appl. Microbiol. 2019, 127, 12–28. [Google Scholar] [CrossRef] [Green Version]
- De Giani, A.; Zampolli, J.; Di Gennaro, P. Recent trends on biosurfactants with antimicrobial activity produced by bacteria associated with human health: Different perspectives on their properties, challenges, and potential applications. Front. Microbiol. 2021, 12, 655150. [Google Scholar] [CrossRef]
- Van Bogaert, I.N.; Buyst, D.; Martins, J.C.; Roelants, S.L.; Soetaert, W.K. Synthesis of bolaform biosurfactants by an engineered Starmerella bombicola yeast. Biotechnol. Bioeng. 2016, 113, 2644–2651. [Google Scholar] [CrossRef]
- Vasudevan, S.; Prabhune, A.A. Photophysical studies on curcumin-sophorolipid nanostructures: Applications in quorum quenching and imaging. R. Soc. Open Sci. 2018, 5, 170865. [Google Scholar] [CrossRef] [Green Version]
- Lydon, H.L.; Baccile, N.; Callaghan, B.; Marchant, R.; Mitchell, C.A.; Banat, I.M. Adjuvant antibiotic activity of acidic sophorolipids with potential for facilitating wound healing. Antimicrob. Agents Chemother. 2017, 61, e02547-16. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, A.E.; Joshi, S.J.; Al-Wahaibi, Y.M.; Al-Bemani, A.S.; Al-Bahry, S.N.; Al-Maqbali, D.; Banat, I.M. Sophorolipids production by Candida bombicola ATCC 22214 and its potential application in microbial enhanced oil recovery. Front. Microbiol. 2015, 26, 1324. [Google Scholar] [CrossRef] [Green Version]
- Darne, P.; Mehta, M.; Agawane, S.; Prabhune, A. Bioavailability studies of curcumin-sophorolipid nano-conjugates in the aqueous phase: Role in the synthesis of uniform gold nanoparticles. RSC Adv. 2016, 6, 68504–68514. [Google Scholar] [CrossRef]
- Rajasekar, V.; Darne, P.; Prabhune, A.; Kao, R.Y.T.; Solomon, A.P.; Ramage, G.; Samaranayake, L.; Neelakantan, P. A curcumin-sophorolipid nanocomplex inhibits Candida albicans filamentation and biofilm development. Colloids Surf. B Biointerfaces 2021, 200, 111617. [Google Scholar] [CrossRef] [PubMed]
- Lembke, C.; Podbielski, A.; Hidalgo-Grass, C.; Jonas, L.; Hanski, E.; Kreikemeyer, B. Characterization of biofilm formation by clinically relevant serotypes of group A streptococci. Appl. Environ. Microbiol. 2006, 72, 2864–2875. [Google Scholar] [CrossRef] [Green Version]
- Stiefel, P.; Schmidt-Emrich, S.; Maniura-Weber, K.; Ren, Q. Critical aspects of using bacterial cell viability assays with the fluorophores SYTO9 and propidium iodide. BMC Microbiol. 2015, 15, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, D.R.; Brown, B.L.; Clewell, D.B. Analysis of plasmid deoxyribonucleic acid in a cariogenic strain of Streptococcus faecalis: An approach to identifying genetic determinants on cryptic plasmids. J. Bacteriol. 1997, 30, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, A.; Shi, Z.; Neoh, K.G.; Kishen, A. Nanoparticulates for antibiofilm treatment and effect of aging on its antibacterial activity. J. Endod. 2010, 36, 1030–1035. [Google Scholar] [CrossRef]
- Kishen, A.; Upadya, M.; Tegos, G.P.; Hamblin, M.R. Efflux pump inhibitor potentiates antimicrobial photodynamic inactivation of Enterococcus faecalis biofilm. Photochem. Photobiol. 2010, 86, 1343–1349. [Google Scholar] [CrossRef] [Green Version]
- Barnes, A.M.; Ballering, K.S.; Leibman, R.S.; Wells, C.L.; Dunny, G.M. Enterococcus faecalis produces abundant extracellular structures containing DNA in the absence of cell lysis during early biofilm formation. mBio 2012, 24, e00193-12. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, N.G.; Croda, J.; Simionatto, S. Antibacterial mechanisms of Cinnamon and its constituents: A review. Microb. Pathog. 2018, 120, 198–203. [Google Scholar] [CrossRef]
- Manet, S.; Cuvier, A.-S.; Valotteau, C.; Fadda, G.C.; Perez, J.; Karakas, E.; Abel, S.; Baccile, N. Structure of bolaamphiphile sophorolipid micelles characterized with SAXS, SANS, and MD simulations. J. Phys. Chem. B 2015, 119, 13113–13133. [Google Scholar] [CrossRef] [Green Version]
- Joyanes, P.; Pascual, A.; Martínez-Martínez, L.; Hevia, A.; Perea, E.J. In vitro adherence of Enterococcus faecalis and Enterococcus faecium to plastic biomaterials. Clin. Microbiol. Infect. 1999, 5, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Joyanes, P.; Pascual, A.; Martínez-Martínez, L.; Hevia, A.; Perea, E.J. In vitro adherence of Enterococcus faecalis and Enterococcus faecium to urinary catheters. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 19, 124–127. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Kishen, A. Effect of tissue fluids on hydrophobicity and adherence of Enterococcus faecalis to dentin. J. Endod. 2007, 33, 1421–1425. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Sánchez, A.J.; Hernández-Sánchez, H.; Jaramillo-Flores, M.E. Biological activity of glycolipids produced by microorganisms: New trends and possible therapeutic alternatives. Microbiol. Res. 2013, 168, 22–32. [Google Scholar] [CrossRef]
- Di Pasqua, R.; Hoskins, N.; Betts, G.; Mauriello, G. Changes in membrane fatty acids composition of microbial cells induced by addiction of thymol, carvacrol, limonene, cinnamaldehyde, and eugenol in the growing media. J. Agric. Food Chem. 2006, 54, 2745–2749. [Google Scholar] [CrossRef]
- Díaz De Rienzo, M.A.; Stevenson, P.; Marchant, R.; Banat, I.M. Antibacterial properties of biosurfactants against selected Gram-positive and -negative bacteria. FEMS Microbiol. Lett. 2016, 363, fnv224. [Google Scholar] [CrossRef] [Green Version]
- Joshi-Navare, K.; Prabhune, A. A biosurfactant-sophorolipid acts in synergy with antibiotics to enhance their efficiency. Biomed. Res. Int. 2013, 2013, 512495. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, M.; Kalimuthu, S.; Zhang, C.; Ali, I.A.A.; Neelakantan, P. Trans-cinnamaldehyde–Biosurfactant Complex as a Potent Agent against Enterococcus faecalis Biofilms. Pharmaceutics 2022, 14, 2355. https://doi.org/10.3390/pharmaceutics14112355
Hu M, Kalimuthu S, Zhang C, Ali IAA, Neelakantan P. Trans-cinnamaldehyde–Biosurfactant Complex as a Potent Agent against Enterococcus faecalis Biofilms. Pharmaceutics. 2022; 14(11):2355. https://doi.org/10.3390/pharmaceutics14112355
Chicago/Turabian StyleHu, Mingxin, Shanthini Kalimuthu, Chengfei Zhang, Islam A. A. Ali, and Prasanna Neelakantan. 2022. "Trans-cinnamaldehyde–Biosurfactant Complex as a Potent Agent against Enterococcus faecalis Biofilms" Pharmaceutics 14, no. 11: 2355. https://doi.org/10.3390/pharmaceutics14112355