
Human Brain Organoids-on-Chip: Advances, Challenges, and Perspectives for Preclinical Applications
 , ,
, ,
Abstract
:1. Introduction
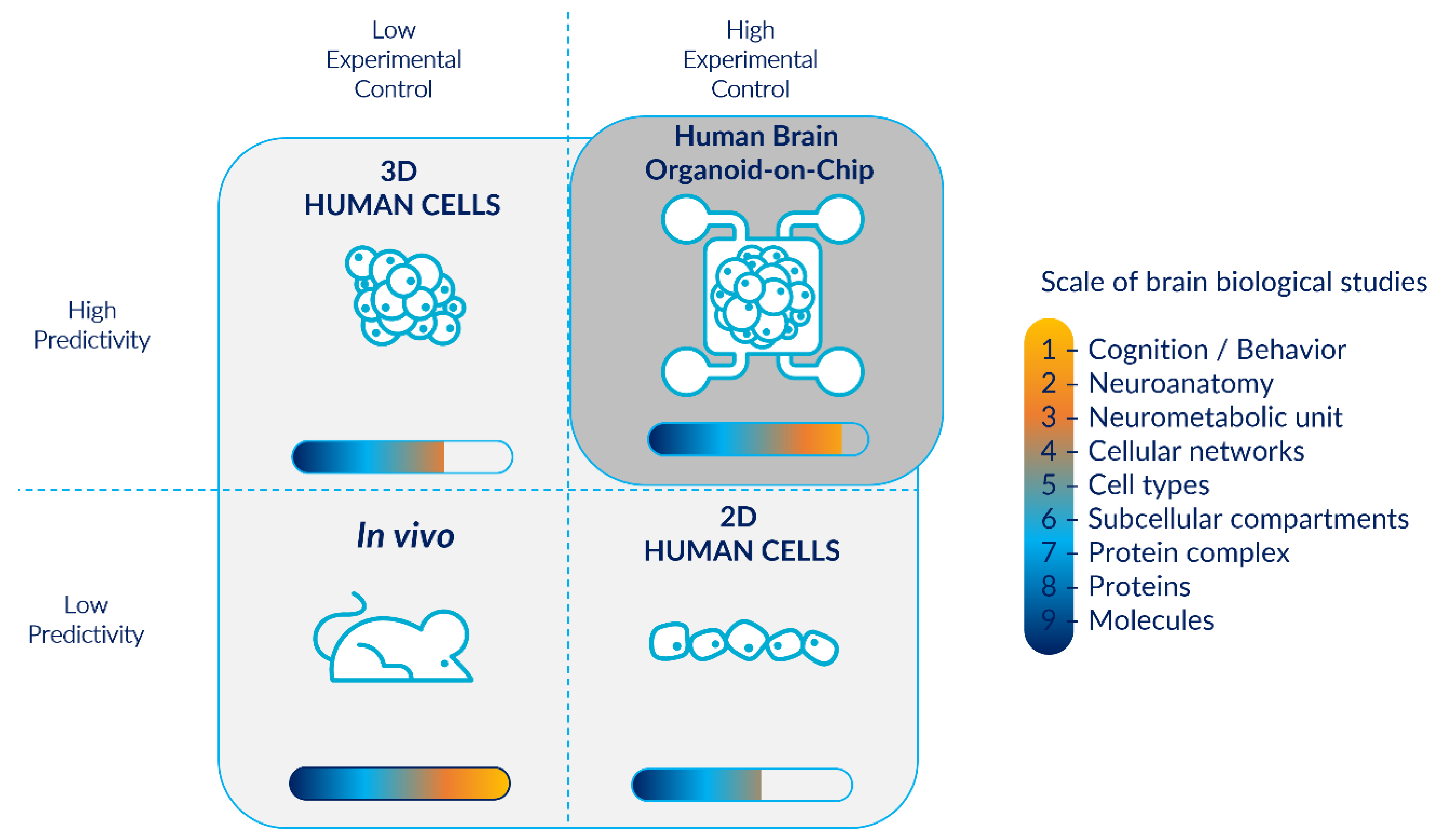
2. Cerebral Organoids: Advantages, Limitations, and Microfluidic Technology as a Solution
2.1. Cerebral Organoids: Promising In Vitro Models of Human Brain Organogenesis
2.2. Cerebral Organoids: A Diversity of Methodologies
2.3. Cerebral Organoids: Examples of Applications in Neurological Disease Modeling
2.4. Challenges for the Adoption of Cerebral Organoids in Preclinical Applications
2.5. Microfluidic Systems: Promising Technologies to Tackle Cerebral Organoid Limitations
3. Brain Organoids-on Chips: State-of-the Art of Promising Models
- (i).
- Microfluidic devices with 3D culture areas and channels: composed of 3D cell culture areas, and channels for culture medium flows (Figure 2i);
- (ii).
- Microfluidic devices with micropillar arrays: comprising micropillars between which cells are cultured in 3D, from iPSCs seeding, to organoid generation, and further growth and expansion (Figure 2ii);
- (iii).
- Microfluidic device with an air-liquid interface: air-liquid integrated to the culture platform (Figure 2iii).
3.1. Microfluidic Devices with 3D-Culture Areas and Channels
3.2. Microfluidic Devices with Micropillar Arrays
3.3. Microfluidic Device with an Air-Liquid Interface
4. Discussion: Current Insight towards Industrial Applications of Brain Organoids-on-Chips in the Field of Pharmacology
4.1. Standardization Methods for Reproducible Cerebral Organoid Generation
4.2. Standardization of Microfluidic Fabrication Process for Reproducible Devices
4.3. Compatibility of Microfluidic Technologies with High-Throughput Pharmacological Assays: Need for Relevant Read-Outs and Data Collection Methods
4.4. Compatibility of Microfluidic Technologies with High-Throughput Pharmacological Assays: Electrophysiology as Relevant Non-Invasive Read-Out
- (i).
- Patch-clamp: provides high temporal resolution, which enables to monitor responses to pharmacological compounds, but little spatial resolution, which prevents information about neuronal networks’ connectivity within organoids ([24]);
- (ii).
- Calcium imaging: enables larger-scale activity information with higher spatial resolution, permitting neural networks analysis within organoids, but with a low temporal resolution, and dependent on imaging capabilities ([20]);
- (iii).
- MEA technologies: provide both high temporal resolution and spatial resolution ([66]). In 2D MEA, cerebral organoids are positioned on the MEA support, and action potentials can be detected and recorded from neurons located on the surface of the organoid in contact with the electrodes.
4.5. Enhancing Capabilities by Coupling 3D Cell Culture with the Blood-Brain Barrier
5. Conclusions
- -
- The establishment of standardized cell culture conditions with defined and reproducible validation and characterization criteria;
- -
- Proof of concept that the technology contributes efficiently and reliably to clinical success for novel therapeutics and improves translational research by providing evidence of successfully predicted human responses;
- -
- Evidence that the model will reduce the need for animal testing while remaining consistent with the scientific aims of the study.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Feigin, V.L.; Nichols, E.; Alam, T.; Bannick, M.S.; Beghi, E.; Blake, N.; Culpepper, W.J.; Dorsey, E.R.; Elbaz, A.; Ellenbogen, R.G.; et al. Global, regional, and national burden of neurological disorders, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 459–480. [Google Scholar] [CrossRef] [Green Version]
- Kesselheim, A.S.; Hwang, T.J.; Franklin, J.M. Two decades of new drug development for central nervous system disorders. Nat. Rev. Drug Discov. 2015, 14, 815–816. [Google Scholar] [CrossRef] [PubMed]
- Abou-El-Enein, M.; Elsanhoury, A.; Reinke, P. Overcoming Challenges Facing Advanced Therapies in the EU Market. Cell Stem Cell 2016, 19, 293–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, F.A.C.; Carvalho, L.R.B.; Grinberg, L.T.; Farfel, J.M.; Ferretti, R.E.L.; Leite, R.E.P.; Filho, W.J.; Lent, R.; Herculano-Houzel, S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol. 2009, 513, 532–541. [Google Scholar] [CrossRef]
- Herculano-Houzel, S. The human brain in numbers: A linearly scaled-up primate brain. Front. Hum. Neurosci. 2009, 3, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Pakkenberg, B.; Pelvig, D.; Marner, L.; Bundgaard, M.J.; Gundersen, H.J.G.; Nyengaard, J.R.; Regeur, L. Aging and the human neocortex. Exp. Gerontol. 2003, 38, 95–99. [Google Scholar] [CrossRef]
- Mateos-Aparicio, P.; Rodríguez-Moreno, A. The impact of studying brain plasticity. Front. Cell. Neurosci. 2019, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Forrester, J.V.; McMenamin, P.G.; Dando, S.J. CNS infection and immune privilege. Nat. Rev. Neurosci. 2018, 19, 655–671. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Prat, A. The blood—Brain barrier. Cold Spring Harb. Perpsectives Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed]
- Holloway, P.M.; Willaime-Morawek, S.; Siow, R.; Barber, M.; Owens, R.M.; Sharma, A.D.; Rowan, W.; Hill, E.; Zagnoni, M. Advances in microfluidic In Vitro systems for neurological disease modeling. J. Neurosci. Res. 2021, 99, 1276–1307. [Google Scholar] [CrossRef] [PubMed]
- Nikolakopoulou, P.; Rauti, R.; Voulgaris, D.; Shlomy, I.; Maoz, B.M.; Herland, A. Recent progress in translational engineered In Vitro models of the central nervous system. Brain 2021, 143, 3181–3213. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Song, H.; Ming, G.L. Brain organoids: Advances, applications and challenges. Development 2019, 146, dev166074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costamagna, G.; Comi, G.P.; Corti, S. Advancing drug discovery for neurological disorders using ipsc-derived neural organoids. Int. J. Mol. Sci. 2021, 22, 2659. [Google Scholar] [CrossRef]
- Coluccio, M.L.; Perozziello, G.; Malara, N.; Parrotta, E.; Zhang, P.; Gentile, F.; Limongi, T.; Raj, P.M.; Cuda, G.; Candeloro, P.; et al. Microfluidic platforms for cell cultures and investigations. Microelectron. Eng. 2019, 208, 14–28. [Google Scholar] [CrossRef]
- Pașca, S.P.; Arlotta, P.; Bateup, H.S.; Camp, J.G.; Cappello, S.; Gage, F.H.; Knoblich, J.A.; Kriegstein, A.R.; Lancaster, M.A.; Ming, G.-L.; et al. A nomenclature consensus for nervous system organoids and assembloids. Nature 2022, 609, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, L.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Rauti, R.; Renous, N.; Maoz, B.M. Mimicking the Brain Extracellular Matrix In Vitro: A Review of Current Methodologies and Challenges. Isr. J. Chem. 2019, 60, 1141–1151. [Google Scholar] [CrossRef]
- Goto-Silva, L.; Ayad, N.M.E.; Herzog, I.L.; Silva, N.P.; Lamien, B.; Orlande, H.R.B.; Da Costa Souza, A.; Ribeiro, S.; Martins, M.; Domont, G.B.; et al. Computational fluid dynamic analysis of physical forces playing a role in brain organoid cultures in two different multiplex platforms. BMC Dev. Biol. 2019, 19, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Su, J.; Wei, J.; Jiang, N.; Ge, X. Recent Advances in Three-Dimensional Stem Cell Culture Systems and Applications. Stem Cells Int. 2021, 2021, 9477332. [Google Scholar] [CrossRef] [PubMed]
- Chiaradia, I.; Lancaster, M.A. Brain organoids for the study of human neurobiology at the interface of In Vitro and In Vivo. Nat. Neurosci. 2020, 23, 1496–1508. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Knoblich, J.A. Generation of Cerebral Organoids from Human Pluripotent Stem Cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [Green Version]
- Pașca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.-Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Susaimanickam, P.J.; Kiral, F.R.; Park, I.H. Region Specific Brain Organoids to Study Neurodevelopmental Disorders. Int. J. Stem Cells 2022, 15, 26–40. [Google Scholar] [CrossRef]
- Xiang, Y.; Tanaka, Y.; Patterson, B.; Kang, Y.-J.; Govindaiah, G.; Roselaar, N.; Cakir, B.; Kim, K.-Y.; Lombroso, A.P.; Hwang, S.-M.; et al. Fusion of regionally-specified hPSC-derived organoids models human brain development and interneuron migration. Cell Stel Cell 2017, 21, 383–398. [Google Scholar] [CrossRef] [Green Version]
- Velasco, S.; Kedaigle, A.J.; Simmons, S.K.; Nash, A.; Rocha, M.; Quadrato, G.; Paulsen, B.; Nguyen, L.; Adiconis, X.; Regev, A.; et al. Individual brain organoids reproducibly form cell diversity of the human cerebral cortex. Nature 2019, 570, 523–527. [Google Scholar] [CrossRef]
- Xiang, Y.; Tanaka, Y.; Cakir, B.; Patterson, B.; Kim, K.-Y.; Sun, P.; Kang, Y.-J.; Zhong, M.; Liu, X.; Patra, P.; et al. hESC-derived thalamic organoids form reciprocal projections when fused with cortical organoids. Cell Stem Cell 2019, 24, 487–497. [Google Scholar] [CrossRef] [Green Version]
- Jo, J.; Xiao, Y.; Sun, A.X.; Cukuroglu, E.; Tran, H.-D.; Göke, J.; Tan, Z.Y.; Saw, T.Y.; Tan, C.-P.; Lokman, H.; et al. Midbrain-like Organoids from Human Pluripotent Stem Cells Contain Functional Dopaminergic and Neuromelanin-Producing Neurons. Cell Stem Cell 2016, 19, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Muguruma, K.; Nishiyama, A.; Kawakami, H.; Hashimoto, K.; Sasai, Y. Self-organization of polarized cerebellar tissue in 3D culture of human pluripotent stem cells. Cell Rep. 2015, 10, 537–550. [Google Scholar] [CrossRef]
- Sakaguchi, H.; Kadoshima, T.; Soen, M.; Narii, N.; Ishida, Y.; Ohgushi, M.; Takahashi, J.; Eiraku, M.; Sasai, Y. Generation of functional hippocampal neurons from self-organizing human embryonic stem cell-derived dorsomedial telencephalic tissue. Nat. Commun. 2015, 6, 8896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Nguyen, H.N.; Song, M.M.; Hadiono, C.; Ogden, S.C.; Hammack, C.; Yao, B.; Hamersky, G.R.; Jacob, F.; Zhong, C.; et al. Brain Region-specific Organoids using Mini-bioreactors for Modeling ZIKV Exposure. Cell 2016, 165, 1238–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Brawner, A.T.; Li, S.; Liu, J.-J.; Kim, H.; Xue, H.; Pang, Z.P.; Kim, W.-Y.; Hart, R.P.; Liu, Y.; et al. OLIG2 Drives Abnormal Neurodevelopmental Phenotypes in Human iPSC-Based Organoid and Chimeric Mouse Models of Down Syndrome. Cell Stel Cell 2019, 24, 908–926. [Google Scholar] [CrossRef]
- Tang, X.Y.; Xu, L.; Wang, J.; Hong, Y.; Wang, Y.; Zhu, Q.; Wang, D.; Zhang, X.-Y.; Liu, C.-Y.; Fang, K.-H.; et al. DSCAM/PAK1 pathway suppression reverses neurogenesis deficits in iPSC-derived cerebral organoids from patients with down syndrome. J. Clin. Invest. 2021, 131, e135763. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Zhu, Y.; Qin, J. Human brain organoid-on-a-chip to model prenatal nicotine exposure. Lab Chip 2018, 18, 851–860. [Google Scholar] [CrossRef]
- Ao, Z.; Cai, H.; Havert, D.J.; Wu, Z.; Gong, Z.; Beggs, J.M.; Mackie, K.; Guo, F. One-Stop Microfluidic Assembly of Human Brain Organoids to Model Prenatal Cannabis Exposure. Anal. Chem. 2020, 92, 4630–4638. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.; Repic, M.; Guo, Z.; Kavirayani, A.; Burkard, T. Genetically engineered cerebral organoids model brain tumour formation. Nat. Methods 2018, 15, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Raja, W.K.; Mungenast, A.E.; Lin, Y.-T.; Ko, T.; Abdurrob, F.; Seo, J.; Tsai, L.-H. Self-organizing 3D human neural tissue derived from induced pluripotent stem cells recapitulate Alzheimer’s disease phenotypes. PLoS ONE 2016, 11, e0161969. [Google Scholar] [CrossRef] [Green Version]
- Nassor, F.; Jarray, R.; Biard, D.S.F.; Maïza, A.; Papy-Garia, D.; Pavoni, S.; Deslys, J.-P.; Yates, F. Long Term Gene Expression in Human Induced Pluripotent Stem Cells and Cerebral Organoids to Model a Neurodegenerative Disease. Front. Cell. Neurosci. 2020, 14, 14. [Google Scholar] [CrossRef]
- Pavoni, S.; Jarray, R.; Nassor, F.; Guyot, A.-C.; Cottin, S.; Rontard, J.; Mikol, J.; Mabondzo, A.; Deslys, J.-P.; Yates, F. Small-molecule induction of Aβ-42 peptide production in human cerebral organoids to model Alzheimer’s disease associated phenotypes. PLoS ONE 2018, 13, e0209150. [Google Scholar] [CrossRef]
- Smits, L.M.; Reinhardt, L.; Reinhardt, P.; Glatza, M.; Monzel, A.S.; Stanslowsky, N.; Rosato-Siri, M.D.; Zanon, A.; Antony, P.M.; Bellman, J.; et al. Modeling Parkinson’s disease in midbrain-like organoids. NPJ Park. Dis. 2019, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Park, H.J.; Choi, H.; Chang, Y.; Park, H.; Shin, J.; Kim, J.; Lengner, C.J.; Lee, Y.K.; Kim, J. Modeling G2019S-LRRK2 Sporadic Parkinson’s Disease in 3D Midbrain Organoids. Stem Cell Rep. 2019, 12, 518–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quadrato, G.; Nguyen, T.; Macosko, E.Z.; Sherwood, J.L.; Yang, S.M.; Berger, D.R.; Maria, N.; Scholvin, J.; Goldman, M.; Kinney, J.P.; et al. Cell diversity and network dynamics in photosensitive human brain organoids. Nature 2017, 545, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giandomenico, S.L.; Sutcliffe, M.; Lancaster, M.A. Generation and long-term culture of advanced cerebral organoids for studying later stages of neural development. Nat. Protoc. 2021, 16, 579–602. [Google Scholar] [CrossRef]
- Ingber, D.E. Reverse Engineering Human Pathophysiology with Organs-on-Chips. Cell 2016, 164, 1105–1109. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.J.; Cho, H. Human brain organoids in Alzheimer’s disease. Organoid 2021, 1, e5. [Google Scholar] [CrossRef]
- Rambani, K.; Vukasinovic, J.; Glezer, A.; Potter, S.M. Culturing thick brain slices: An interstitial 3D microperfusion system for enhanced viability. J. Neurosci. 2009, 180, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Park, S.E.; Georgescu, A.; Huh, D. Organoids-on-a-chip. Science 2019, 364, 960–965. [Google Scholar] [CrossRef]
- Jeon, N.L.; Baskaran, H.; Dertinger, S.K.W.; Whitesides, G.M. Neutrophil chemotaxis in linear and complex gradients of interleukin-8 formed in a microfabricated device. Nat. Biotechnol. 2002, 20, 826–830. [Google Scholar] [CrossRef]
- Chung, B.G.; Flanagan, L.A.; Rhee, S.W.; Schwartz, P.H.; Lee, A.P.; Monuki, E.S.; Jeon, N.L. Human neural stem cell growth and differentiation in a gradient-generating microfluidic device. Lab Chip 2005, 5, 401–406. [Google Scholar] [CrossRef]
- Cassel de Camps, C.; Aslani, S.; Stylianesis, N.; Nami, H.; Mohamed, N.-V.; Durcan, T.M.; Moraes, C. Hydrogel Mechanics Influence the Growth and Development of Embedded Brain Organoids. ACS Appl. Bio Mater. 2022, 5, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Blurton-jones, M.; Rhee, S.W.; Cribbs, D.H.; Cotman, C.W. A microfluidic culture platform for CNS axonal injury, regeneration and transport. Nat. Methods 2005, 2, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, Q.; Batut, A.; Gleyzes, M.; Rontard, J.; Miny, L.; Libralato, M.; Vieira, J.; Debis, D.; Larramendy, F.; Honegger, T.; et al. Co-culture of Glutamatergic Neurons and Pediatric High-Grade Glioma Cells into Microfluidic Devices to Assess Electrical Interactions. J. Vis. Exp. 2021, 177, 1–15. [Google Scholar] [CrossRef]
- Honegger, T.; Thielen, M.I.; Feizi, S.; Sanjana, N.E.; Voldman, J. Microfluidic neurite guidance to study structure-function relationships in topologically-complex population-based neural networks. Sci. Rep. 2016, 6, 28384. [Google Scholar] [CrossRef] [Green Version]
- Maisonneuve, B.G.C.; Libralesso, L.; Miny, L.; Batut, A.; Rontard, J.; Gleyzes, M.; Boudra, B.; Viera, J.; Debis, D.; Larramendy, F.; et al. Deposition chamber technology as building blocks for a standardized brain-on-chip framework. Microsyst. Nanoeng. 2022, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Miny, L.; Maisonneuve, B.G.C.; Quadrio, I.; Honegger, T. Modeling Neurodegenerative Diseases Using In Vitro Compartimentalized Microfluidic Devices. Front. Bioeng. Biotechnol. 2022, 10, 919646. [Google Scholar] [CrossRef]
- Sokolowska, P.; Zuchowska, A.; Brzozka, Z. Why Can Organoids Improve Current Organ-on-Chip Platforms? Organoids 2022, 1, 69–84. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Guo, Y.; Zhu, Y.; Qin, J. Engineering stem cell-derived 3D brain organoids in a perfusable organ-on-a-chip system. RSC Adv. 2018, 8, 1677–1685. [Google Scholar] [CrossRef] [Green Version]
- Cho, A.-N.; Jin, Y.; An, Y.; Kim, J.; Choi, Y.S.; Lee, J.S.; Kim, J.; Choi, W.-Y.; Koo, D.-J.; Yu, W.; et al. Microfluidic device with brain extracellular matrix promotes structural and functional maturation of human brain organoids. Nat. Commun. 2021, 12, 4730. [Google Scholar] [CrossRef]
- Karzbrun, E.; Kshirsagar, A.; Cohen, S.R.; Hanna, J.H.; Reiner, O. Human brain organoids on a chip reveal the physics of folding. Nat. Phys. 2018, 14, 515–522. [Google Scholar] [CrossRef]
- Salmon, I.; Grebenyuk, S.; Abdel Fattah, A.R.; Rustandi, G.; Pilkington, T.; Verfaillie, C.; Ranga, A. Engineering neurovascular organoids with 3D printed microfluidic chips. Lab Chip 2022, 22, 1615–1629. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, L.; Yu, H.; Yin, F.; Wang, Y.; Liu, H.; Jiang, L.; Qin, J. In Situ generation of human brain organoids on a micropillar array. Lab Chip 2017, 17, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Yin, F.; Zhu, Y.; Wang, Y.; Qin, J. Engineering Brain Organoids to Probe Impaired Neurogenesis Induced by Cadmium. ACS Biomater. Sci. Eng. 2018, 4, 1908–1915. [Google Scholar] [CrossRef]
- Cui, K.; Wang, Y.; Zhu, Y.; Tao, T.; Yin, F.; Guo, Y.; Liu, H.; Li, F.; Wang, P.; Chen, Y.; et al. Neurodevelopmental impairment induced by prenatal valproic acid exposure shown with the human cortical organoid-on-a-chip model. Microsyst. Nanoeng. 2020, 6, 49. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Chen, W.; Cao, R.; Xie, Y.; Wang, P.; Wu, Y.; Wang, Y.; Qin, J. Brain organoid-on-chip system to study the effects of breast cancer derived exosomes on the neurodevelopment of brain. Cell Regen. 2022, 11, 7. [Google Scholar] [CrossRef]
- Giandomenico, S.L.; Mierau, S.B.; Gibbons, G.M.; Wenger, L.M.D.; Masullo, L.; Sit, T.; Sutcliffe, M.; Boulanger, J.; Tripodi, M.; Derivery, E.; et al. Cerebral organoids at the air—Liquid interface generate diverse nerve tracts with functional output. Nat. Neurosci. 2019, 22, 669–679. [Google Scholar] [CrossRef]
- Raj, M.K.; Chakraborty, S. PDMS microfluidics: A mini review. J. Appl. Polym. Sci. 2020, 137, 48958. [Google Scholar] [CrossRef]
- Grant, J.; Özkan, A.; Oh, C.; Mahajan, G.; Prantil-Baun, R.; Ingber, D.E. Simulating drug concentrations in PDMS microfluidic organ chips. 2021, 21, 3509–3519. Lab Chip. [CrossRef]
- Durens, M.; Nestor, J.; Williams, M.; Herold, K.; Niescier, R.F.; Lunden, J.W.; Philips, A.W.; Lin, Y.-C.; Dykxhoorn, D.M.; Nestor, M.W. High-throughput screening of human induced pluripotent stem cell-derived brain organoids. J. Neurosci. Methods 2020, 335, 108627. [Google Scholar] [CrossRef]
- Kiaee, K.; Jodat, Y.A.; Bassous, N.J.; Matharu, N.; Shin, S.R. Transcriptomic mapping of neural diversity, differentiation and functional trajectory in ipsc-derived 3d brain organoid models. Cells 2021, 10, 3422. [Google Scholar] [CrossRef]
- Marx, V. Method of the Year: Spatially resolved transcriptomics. Nat. Methods 2021, 18, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.B.; Chane, C.S.; Clouchoux, C.; Histace, A. Recent Trends and Perspectives in Cerebral Organoids Imaging and Analysis. Front. Neurosci. 2021, 15, 629067. [Google Scholar] [CrossRef] [PubMed]
- Dries, R.; Chen, J.; del Rossi, N.; Khan, M.M.; Sistig, A.; Yuan, G.C. Advances in spatial transcriptomic data analysis. Genome Res. 2021, 31, 1706–1718. [Google Scholar] [CrossRef] [PubMed]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. Ilastik: Interactive Machine Learning for (Bio)Image Analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Passaro, A.P.; Stice, S.L. Electrophysiological Analysis of Brain Organoids: Current Approaches and Advancements. Front. Neurosci. 2021, 14, 622137. [Google Scholar] [CrossRef]
- Tasnim, K.; Liu, J. Emerging Bioelectronics for Brain Organoid Electrophysiology. J. Mol. Biol. 2022, 434, 167165. [Google Scholar] [CrossRef]
- Schröter, M.; Wang, C.; Terrigno, M.; Hornauer, P.; Huang, Z.; Jagasia, R.; Hierlemann, A. Functional imaging of brain organoids using high-density microelectrode arrays. MRS Bull. 2022, 47, 530–544. [Google Scholar] [CrossRef]
- Bridges, D.C.; Tovar, K.R.; Wu, B.; Hansma, P.K.; Kosik, K.S. MEA Viewer: A high-performance interactive application for visualizing electrophysiological data. PLoS ONE 2018, 13, e0192477. [Google Scholar] [CrossRef] [Green Version]
- Carton, F.; Malatesta, M. In Vitro Models of Biological Barriers for Nanomedical Research. Int. J. Mol. Sci. 2022, 23, 8910. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Device | Fields of Study | Protocol for Organoid Generation | Characteristics of the Microfluidic Device | Advantages of Microfluidic Technology for Cerebral Organoids Culture | References |
|---|---|---|---|---|---|
| Microfluidic devices with 3D culture areas and channels | Neurodevelopmental toxicity: tests of prenatal nicotine exposure effects on neurodevelopment | Unguided (Lancaster’s protocol) |
|
| [35,58] |
| Modeling of the cerebrospinal fluid flow | Unguided (Lancaster’s protocol) |
|
| [59] | |
| Modeling of cerebral folding and diseased modeling of lissencephaly | Unguided (not Lancaster’s protocol) |
|
| [60] | |
| Vascularized cerebral organoid in a microfluidic device | Unguided (adapted from Lancaster’s protocol) |
|
| [61] | |
| Microfluidic devices with micropillar arrays | Neurodevelopmental toxicity: tests of prenatal cadmium exposure effects on neurodevelopment | Unguided (Lancaster’s protocol) |
|
| [62,63] |
| Neurodevelopmental toxicity: tests of prenatal exposure effects on neurodevelopment with valproic acid and breast cancer-derived exosomes | Guided (cortical organoid) |
|
| [64,65] | |
| Microfluidic devices with air-liquid interface | Neurodevelopmental toxicity: tests of prenatal cannabis exposure effects on neurodevelopment | Unguided (STEMdiff cerebral organoid kit), adapted from Lancaster’s protocol) |
|
| [36] |
| Type of Device | References | Scalability | Reproducibility | Maturity * | Functionality ** | Drug Permeability (BBB Modeling) |
|---|---|---|---|---|---|---|
| Microfluidic devices with 3D culture areas and channels | [35,58]: Perfusable device with culture channels + perfusion channels Prenatal nicotine exposure | Possible | Yes | 33 days | No | No |
| [59]: Device modeling cerebrospinal fluid flow + with brain ECM | Possible | Yes | 120 days | +/− (Ca2+, patch-clamp) | No | |
| [60]: Device with constrained culture chamber Cerebral folding and wrinkles Lissencephaly modelling | Does not seem possible | Yes | 20 days | No | No | |
| [61]: Device to develop vascularized cerebral organoids | Possible | Yes | 30 days | No | +/− | |
| Microfluidic devices with micropillar arrays | [62,63]: Prenatal cadmium exposure | Possible | Yes | 40 days | No | No |
| [64,65]: Prenatal exposure with valproic acid and breast cancer-derived exosomes | Possible | Yes | 70 days | No | No | |
| Microfluidic devices with air-liquid interface | [36]: Prenatal cannabis exposure | Yes | Yes | 90 days | +/− (2D MEA) | +/− |
| Type of Device | References | Structural Visualization | Functional Activity Analysis | Transcriptomic Analysis | Proteomic Analysis | Metabolomic Analysis | Cellular Viability Analysis |
|---|---|---|---|---|---|---|---|
| Microfluidic devices with 3D culture areas and channels | [35,58]: Perfusable device with culture channels + perfusion channels Prenatal nicotine exposure | IF staining (on organoid cryosections) | Ø | RT-qPCR | Ø | Ø | TUNEL assay (on organoid cryosections) |
| [59]: Device modeling cerebrospinal fluid flow + with brain ECM | IF staining (on organoid cryosections) + image-based quantifications |
|
| LC-MS |
|
| |
| [60]: Device with constrained culture chamber Cerebral folding and wrinkles Lissencephaly modelling |
| Ø |
| Ø | Ø | Ø | |
| [61]: Device to develop vascularized cerebral organoids | IF staining (directly in the device) | Ø | RT-qPCR | Ø | Ø | Ø | |
| Microfluidic devices with micropillar arrays | [62,63]: Prenatal cadmium exposure | IF staining (on organoid cryosections) | Ø | RT-qPCR | Ø | Ø | Ø |
| [64,65]: Prenatal exposure with valproic acid and breast cancer-derived exosomes | IF staining (on organoid cryosections) + image-based quantifications | Ø |
| Ø | Ø | Cellular viability + hypoxia (directly in the device) | |
| Microfluidic devices with air-liquid interface | [36]: Prenatal cannabis exposure | IF staining (on organoid cryosections) | 2D MEA | RT-qPCR | Ø | Ø | Ø |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castiglione, H.; Vigneron, P.-A.; Baquerre, C.; Yates, F.; Rontard, J.; Honegger, T. Human Brain Organoids-on-Chip: Advances, Challenges, and Perspectives for Preclinical Applications. Pharmaceutics 2022, 14, 2301. https://doi.org/10.3390/pharmaceutics14112301
Castiglione H, Vigneron P-A, Baquerre C, Yates F, Rontard J, Honegger T. Human Brain Organoids-on-Chip: Advances, Challenges, and Perspectives for Preclinical Applications. Pharmaceutics. 2022; 14(11):2301. https://doi.org/10.3390/pharmaceutics14112301
Chicago/Turabian StyleCastiglione, Héloïse, Pierre-Antoine Vigneron, Camille Baquerre, Frank Yates, Jessica Rontard, and Thibault Honegger. 2022. "Human Brain Organoids-on-Chip: Advances, Challenges, and Perspectives for Preclinical Applications" Pharmaceutics 14, no. 11: 2301. https://doi.org/10.3390/pharmaceutics14112301