
Immunotherapeutic Implications of Toll-like Receptors Activation in Tumor Microenvironment

Abstract
:1. Introduction
2. Activation of Toll-like Receptors
2.1. Toll-like Receptors
2.2. TLR-Activated Pathways
3. Tumor Microenvironment and TLRs Activation
3.1. Tumor Microenvironment and Cancer Development
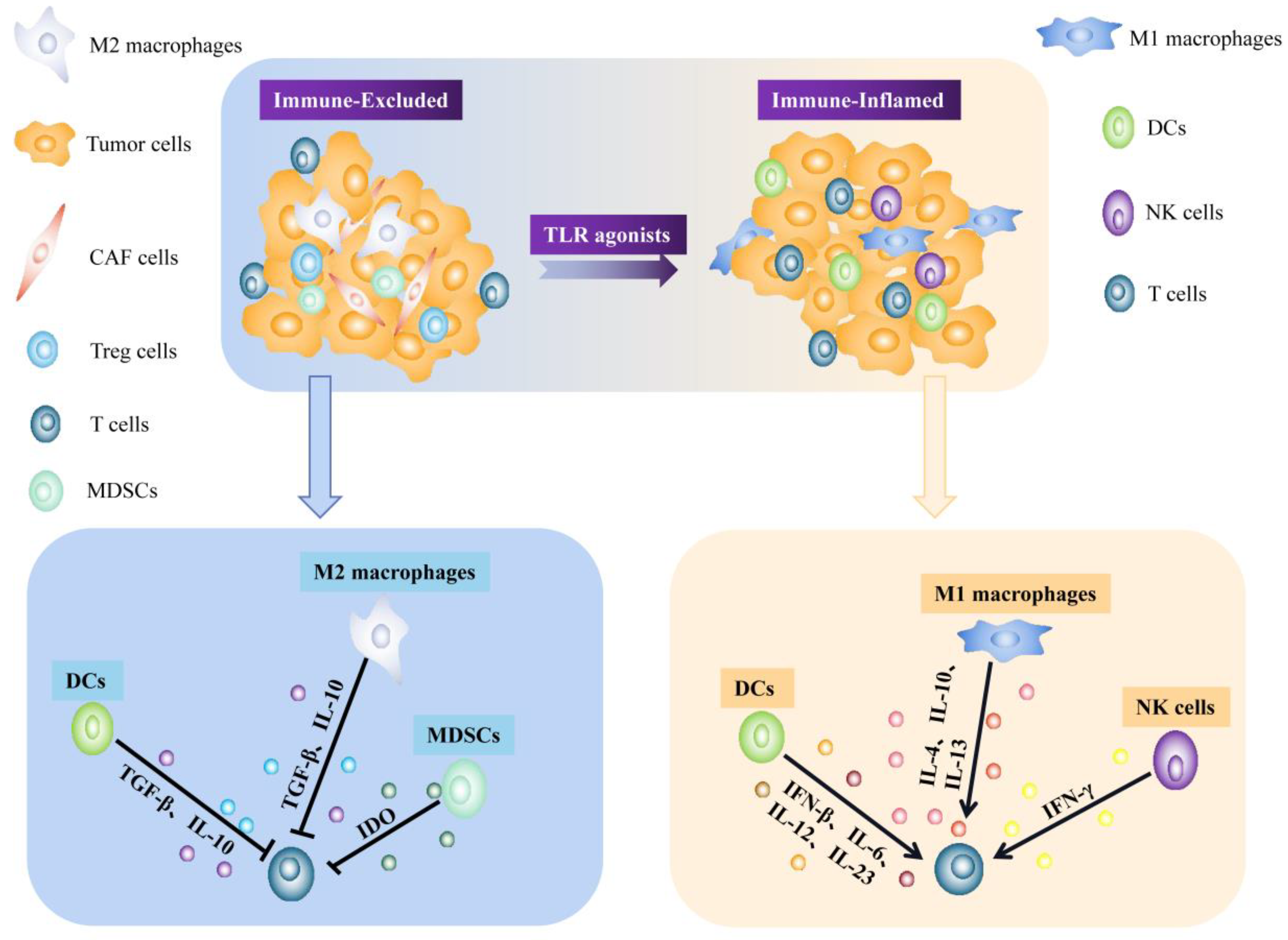
3.2. Tumor Microenvironment and Cancer Immunotherapy
3.3. Expression and Activation of TLRs in Tumor Microenvironment
- (1)
- Dendritic cells
- (2)
- Natural killer cells
- (3)
- Tumor-associated macrophages
- (4)
- T cells
- (5)
- Mast cells
4. Dual Role of TLRs Activation in the Tumor Microenvironment
5. Immunotherapy Applications
5.1. TLR Agonists and TLR Inhibitors
5.1.1. Functions of TLR Agonists and TLR Inhibitors
5.1.2. Developed TLR Agonists and TLR Inhibitors

{kind=link}
{kind=link}
| TLR | TLR Agonist | Structural Characteristic | Applications | Combination Therapy | Results | Refs. |
|---|---|---|---|---|---|---|
| TLR7/8 | Telratolimod (Tel) | Hydrophobic long chain | Significantly prevent B16F10 or 4T-1 tumor postoperative recurrence and metastasis | Tel@PGE in combination with chemotherapy and radiotherapy | Recruitment of effector CD8+ T lymphocytes and the polarization of MDSCs | [138] |
| TLR7/8 | MEDI9197 (3M-052) | Lipophilicity | Intratumoral injection for local TLR7/8 agonism | Combined with T-cell targeted immunotherapy | Th1 polarization, enrichment and activation of NK cells and CD8+ T cells | [139] |
| TLR9 | EnanDIM | L-nucleotide-protected | CT26 colon cancer tumor, MC38 colon cancer, B16 melanoma, A20 lymphoma and EMT-6 breast cancer | EnanDIM® Combination Immunotherapy | Secretion of IFN-α and IP-10 and activation of immune cells | [140] |
| TLR9 | MGN1703 | DNA-based | Multiple solid tumors | As an alternative to cancer immunotherapy | Activation of innate immune response with DAMP | [141] |
| TLR9 | CMP-001 (vidutolimod) | Virus-like particles | Activate immune cells in peripheral blood | As a novel immunotherapy | Anti-Qβ antibody reaction causes uptake of particles by pDCs, producing IFN-α | [142] |
| TLR9 | IMO-2125 | Containing a phosphorothioate backbone | Intratumoral injection | Co-administration of anti-CTLA-4 and anti-PD-1 | Create a long-lived tumor-specific immune memory of CT26 antigens | [143] |
| TLR2 | TLR2-L Amplivant (AV) | Splicing of Pam3CSK4 with synthetic long peptide (SLP) | Cancer Vaccination | AV-SLP couples in combination with chemotherapy or photodynamic therapy | Enhance activation of DCs in vitro | [144] |
| TLR1/TLR2 | Diprovocims | With structure-activity (SAR) relationships | As vaccine adjuvant and cancer treatment | Synergistic action with checkpoint inhibitors | Promotes the release of TNF-α from human THP-1 bone marrow cells | [145] |
| TLR3 | ARNAX | Nucleic acid adjuvants | Safe for older cancer patients receiving immunotherapy | As vaccine immunotherapy | Induces tumor-specific memory T cells and durable anti-tumor immunity without inducing systemic inflammation | [146,147,148] |
| TLR5 | Mobilan | Adenovirus | Injected into the prostate of transgenic mice (TRAMP) | Combined with radiation and vaccination | Constitutive activation of NF-κB in vitro and in vivo | [149] |
5.1.3. TLR Agonists and Inhibitors in Clinical Trials
5.2. Delivery Methods for TLR Agonists and TLR Inhibitors
5.2.1. Nano-Delivery System
5.2.2. Exosome Delivery System
5.2.3. Liposomal Delivery System
5.3. Combination Applications for TLR Agonists and TLR Inhibitors
5.3.1. TLR Agonists in Combination with Other Therapies
5.3.2. TLR Agonists as Vaccine Adjuvants
5.3.3. Co-Delivery of TLR Agonists/Antagonists with Nanoparticle for Cancer Treatment
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Z.; Hou, B. TLR signaling in B-cell development and activation. Cell Mol. Immunol. 2013, 10, 103–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basith, S.; Manavalan, B.; Yoo, T.H.; Kim, S.G.; Choi, S. Roles of toll-like receptors in cancer: A double-edged sword for defense and offense. Arch. Pharm. Res. 2012, 35, 1297–1316. [Google Scholar] [CrossRef] [PubMed]
- Dajon, M.; Iribarren, K.; Cremer, I. Toll-like receptor stimulation in cancer: A pro- and anti-tumor double-edged sword. Immunobiology 2017, 222, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, L.; Chen, S. Dual character of Toll-like receptor signaling: Pro-tumorigenic effects and anti-tumor functions. Biochim. Biophys. Acta 2013, 1835, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Anandappa, A.J.; Wu, C.J.; Ott, P.A. Directing Traffic: How to Effectively Drive T Cells into Tumors. Cancer Discov. 2020, 10, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Bourquin, C.; Pommier, A.; Hotz, C. Harnessing the immune system to fight cancer with Toll-like receptor and RIG-I-like receptor agonists. Pharmacol. Res. 2020, 154, 104192. [Google Scholar] [CrossRef] [Green Version]
- Keshavarz, A.; Pourbagheri-Sigaroodi, A.; Zafari, P.; Bagheri, N.; Ghaffari, S.H.; Bashash, D. Toll-like receptors (TLRs) in cancer; with an extensive focus on TLR agonists and antagonists. IUBMB Life 2021, 73, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Tartey, S.; Takeuchi, O. Pathogen recognition and Toll-like receptor targeted therapeutics in innate immune cells. Int. Rev. Immunol. 2017, 36, 57–73. [Google Scholar] [CrossRef]
- Lim, K.H.; Staudt, L.M. Toll-like receptor signaling. Cold Spring Harb. Perspect. Biol. 2013, 5, a011247. [Google Scholar] [CrossRef] [PubMed]
- Pradere, J.P.; Dapito, D.H.; Schwabe, R.F. The Yin and Yang of Toll-like receptors in cancer. Oncogene 2014, 33, 3485–3495. [Google Scholar] [CrossRef] [Green Version]
- Javaid, N.; Choi, S. Toll-like Receptors from the Perspective of Cancer Treatment. Cancers 2020, 12, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.S.; Lee, J.O. Structures of the toll-like receptor family and its ligand complexes. Immunity 2008, 29, 182–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, H.; Qian, C.; Cao, X. Regulation of Toll-like receptor signaling in the innate immunity. Sci. Chin. Life Sci. 2010, 53, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.J.; Gilmore, T.D. Evolutionary Origins of Toll-like Receptor Signaling. Mol. Biol. Evol. 2018, 35, 1576–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, K.J.; Coban, C.; Akira, S. Manifold mechanisms of Toll-like receptor-ligand recognition. J. Clin. Immunol. 2005, 25, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.C.; Mosaheb, M.M.; Mohme, M.; McKay, Z.P.; Holl, E.K.; Kastan, J.P.; Yang, Y.; Beasley, G.M.; Hwang, E.S.; Ashley, D.M.; et al. Viral infection of cells within the tumor microenvironment mediates antitumor immunotherapy via selective TBK1-IRF3 signaling. Nat. Commun. 2021, 12, 1858. [Google Scholar] [CrossRef] [PubMed]
- Robinet, M.; Maillard, S.; Cron, M.A.; Berrih-Aknin, S.; Le Panse, R. Review on Toll-Like Receptor Activation in Myasthenia Gravis: Application to the Development of New Experimental Models. Clin. Rev. Allergy Immunol. 2017, 52, 133–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamdar, K.; Nguyen, V.; DePaolo, R.W. Toll-like receptor signaling and regulation of intestinal immunity. Virulence 2013, 4, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Li, H.; Wang, H.; Zhang, T.; Hutchinson, M.R.; Yin, H.; Wang, X. Small-Molecule Modulators of Toll-like Receptors. Acc. Chem. Res. 2020, 53, 1046–1055. [Google Scholar] [CrossRef]
- Kirtland, M.E.; Tsitoura, D.C.; Durham, S.R.; Shamji, M.H. Toll-Like Receptor Agonists as Adjuvants for Allergen Immunotherapy. Front. Immunol. 2020, 11, 599083. [Google Scholar] [CrossRef] [PubMed]
- Suhir, H.; Etzioni, A. The role of Toll-like receptor signaling in human immunodeficiencies. Clin. Rev. Allergy Immunol. 2010, 38, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Bockamp, E.; Rosigkeit, S.; Siegl, D.; Schuppan, D. Nano-Enhanced Cancer Immunotherapy: Immunology Encounters Nanotechnology. Cells 2020, 9, 2102. [Google Scholar] [CrossRef]
- Pearce, O.M.T.; Delaine-Smith, R.M.; Maniati, E.; Nichols, S.; Wang, J.; Bohm, S.; Rajeeve, V.; Ullah, D.; Chakravarty, P.; Jones, R.R.; et al. Deconstruction of a Metastatic Tumor Microenvironment Reveals a Common Matrix Response in Human Cancers. Cancer Discov. 2018, 8, 304–319. [Google Scholar] [CrossRef] [Green Version]
- Parker, T.M.; Gupta, K.; Palma, A.M.; Yekelchyk, M.; Fisher, P.B.; Grossman, S.R.; Won, K.J.; Madan, E.; Moreno, E.; Gogna, R. Cell competition in intratumoral and tumor microenvironment interactions. EMBO J. 2021, 40, e107271. [Google Scholar] [CrossRef] [PubMed]
- Gasser, S.; Lim, L.H.K.; Cheung, F.S.G. The role of the tumour microenvironment in immunotherapy. Endocr. Relat. Cancer 2017, 24, T283–T295. [Google Scholar] [CrossRef] [PubMed]
- Casey, S.C.; Amedei, A.; Aquilano, K.; Azmi, A.S.; Benencia, F.; Bhakta, D.; Bilsland, A.E.; Boosani, C.S.; Chen, S.; Ciriolo, M.R.; et al. Cancer prevention and therapy through the modulation of the tumor microenvironment. Semin. Cancer Biol. 2015, 35, S199–S223. [Google Scholar] [CrossRef] [PubMed]
- Rameshbabu, S.; Labadie, B.W.; Argulian, A.; Patnaik, A. Targeting Innate Immunity in Cancer Therapy. Vaccines 2021, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Clubb, J.D.; Chen, Y.Y. Engineering CAR-T Cells for Next-Generation Cancer Therapy. Cancer Cell 2020, 38, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Grosser, R.; Cherkassky, L.; Chintala, N.; Adusumilli, P.S. Combination Immunotherapy with CAR T Cells and Checkpoint Blockade for the Treatment of Solid Tumors. Cancer Cell 2019, 36, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, S. Toll-like receptors expressed in tumor cells: Targets for therapy. Cancer Immunol. Immunother. 2008, 57, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Xun, Y.; Yang, H.; Kaminska, B.; You, H. Toll-like receptors and toll-like receptor-targeted immunotherapy against glioma. J. Hematol. Oncol. 2021, 14, 176. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhu, S.; Qiao, Y.; Liu, Y.J.; Chen, W.; Zhao, G.; Chen, J. Recent advances in the role of toll-like receptors and TLR agonists in immunotherapy for human glioma. Protein Cell 2014, 5, 899–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Wang, L.; Chen, S. Exogenous or endogenous Toll-like receptor ligands: Which is the MVP in tumorigenesis? Cell Mol. Life Sci. 2012, 69, 935–949. [Google Scholar] [CrossRef]
- El-Omar, E.M.; Ng, M.T.; Hold, G.L. Polymorphisms in Toll-like receptor genes and risk of cancer. Oncogene 2008, 27, 244–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, D.; Yusuf, N. Expression of toll-like receptors on breast tumors: Taking a toll on tumor microenvironment. Int. J. Breast Cancer 2012, 2012, 716564. [Google Scholar] [CrossRef] [Green Version]
- Jang, G.Y.; Lee, J.W.; Kim, Y.S.; Lee, S.E.; Han, H.D.; Hong, K.J.; Kang, T.H.; Park, Y.M. Interactions between tumor-derived proteins and Toll-like receptors. Exp. Mol. Med. 2020, 52, 1926–1935. [Google Scholar] [CrossRef] [PubMed]
- Yeh, D.W.; Huang, L.R.; Chen, Y.W.; Huang, C.F.; Chuang, T.H. Interplay between Inflammation and Stemness in Cancer Cells: The Role of Toll-Like Receptor Signaling. J. Immunol. Res. 2016, 2016, 4368101. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Zhao, J.; Unkeless, J.C.; Feng, Z.H.; Xiong, H. TLR signaling by tumor and immune cells: A double-edged sword. Oncogene 2008, 27, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, H.; Peng, G. TLR-mediated metabolic reprogramming in the tumor microenvironment: Potential novel strategies for cancer immunotherapy. Cell Mol. Immunol. 2018, 15, 428–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Meng, T.; Chen, L.; Wei, W.; Wang, P. The role of ubiquitination in tumorigenesis and targeted drug discovery. Signal Transduct. Target Ther. 2020, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatehchand, K.; Ren, L.; Elavazhagan, S.; Fang, H.; Mo, X.; Vasilakos, J.P.; Dietsch, G.N.; Hershberg, R.M.; Tridandapani, S.; Butchar, J.P. Toll-like Receptor 4 Ligands Down-regulate Fcgamma Receptor IIb (FcgammaRIIb) via MARCH3 Protein-mediated Ubiquitination. J. Biol. Chem. 2016, 291, 3895–3904. [Google Scholar] [CrossRef] [Green Version]
- Gerhard, G.M.; Bill, R.; Messemaker, M.; Klein, A.M.; Pittet, M.J. Tumor-infiltrating dendritic cell states are conserved across solid human cancers. J. Exp. Med. 2021, 218, e20200264. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.; Chintala, S.; Dey, M. Plasmacytoid dendritic cell in immunity and cancer. J. Neuroimmunol. 2018, 322, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Wimmers, F.; Subedi, N.; van Buuringen, N.; Heister, D.; Vivie, J.; Beeren-Reinieren, I.; Woestenenk, R.; Dolstra, H.; Piruska, A.; Jacobs, J.F.M.; et al. Single-cell analysis reveals that stochasticity and paracrine signaling control interferon-alpha production by plasmacytoid dendritic cells. Nat. Commun. 2018, 9, 3317. [Google Scholar] [CrossRef] [Green Version]
- Watts, C.; Zaru, R.; Prescott, A.R.; Wallin, R.P.; West, M.A. Proximal effects of Toll-like receptor activation in dendritic cells. Curr. Opin. Immunol. 2007, 19, 73–78. [Google Scholar] [CrossRef]
- Krawczyk, C.M.; Holowka, T.; Sun, J.; Blagih, J.; Amiel, E.; DeBerardinis, R.J.; Cross, J.R.; Jung, E.; Thompson, C.B.; Jones, R.G.; et al. Toll-like receptor-induced changes in glycolytic metabolism regulate dendritic cell activation. Blood 2010, 115, 4742–4749. [Google Scholar] [CrossRef] [Green Version]
- Lovgren, T.; Sarhan, D.; Truxova, I.; Choudhary, B.; Maas, R.; Melief, J.; Nystrom, M.; Edback, U.; Vermeij, R.; Scurti, G.; et al. Enhanced stimulation of human tumor-specific T cells by dendritic cells matured in the presence of interferon-gamma and multiple toll-like receptor agonists. Cancer Immunol. Immunother. 2017, 66, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Takeda, Y.; Seya, T. Targeting Toll-like receptor 3 in dendritic cells for cancer immunotherapy. Expert Opin. Biol. Ther. 2020, 20, 937–946. [Google Scholar] [CrossRef]
- Roselli, E.; Araya, P.; Nunez, N.G.; Gatti, G.; Graziano, F.; Sedlik, C.; Benaroch, P.; Piaggio, E.; Maccioni, M. TLR3 Activation of Intratumoral CD103(+) Dendritic Cells Modifies the Tumor Infiltrate Conferring Anti-tumor Immunity. Front. Immunol. 2019, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.; Yong, M.; Raju, J.; Willemsen, N.; Black, M.; Trent, A.; Tirrell, M.; Olive, C. Toll-like receptor-mediated adjuvanticity and immunomodulation in dendritic cells: Implications for peptide vaccines. Hum. Vaccin. 2011, 7 (Suppl. 1), 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bald, T.; Krummel, M.F.; Smyth, M.J.; Barry, K.C. The NK cell-cancer cycle: Advances and new challenges in NK cell-based immunotherapies. Nat. Immunol. 2020, 21, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hu, Y.; Shi, C. Targeting Natural Killer Cells for Tumor Immunotherapy. Front. Immunol. 2020, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, J.Y.; Yoon, S.R.; Kim, T.D.; Choi, I.; Jung, H. Toll-Like Receptors in Natural Killer Cells and Their Application for Immunotherapy. J. Immunol. Res. 2020, 2020, 2045860. [Google Scholar] [CrossRef]
- Veneziani, I.; Alicata, C.; Pelosi, A.; Landolina, N.; Ricci, B.; D’Oria, V.; Fagotti, A.; Scambia, G.; Moretta, L.; Maggi, E. Toll-like receptor 8 agonists improve NK-cell function primarily targeting CD56(bright)CD16(-) subset. J. Immunother. Cancer 2022, 10, e003385. [Google Scholar] [CrossRef]
- Cheng, N.; Bai, X.; Shu, Y.; Ahmad, O.; Shen, P. Targeting tumor-associated macrophages as an antitumor strategy. Biochem. Pharmacol. 2021, 183, 114354. [Google Scholar] [CrossRef]
- Ngambenjawong, C.; Gustafson, H.H.; Pun, S.H. Progress in tumor-associated macrophage (TAM)-targeted therapeutics. Adv. Drug Deliv. Rev. 2017, 114, 206–221. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Q.; Jewell, C.M. Directing toll-like receptor signaling in macrophages to enhance tumor immunotherapy. Curr. Opin. Biotechnol. 2019, 60, 138–145. [Google Scholar] [CrossRef]
- Safarzadeh, E.; Mohammadi, A.; Mansoori, B.; Duijf, P.H.G.; Hashemzadeh, S.; Khaze, V.; Kazemi, T.; Derakhshani, A.; Silvestris, N.; Baradaran, B. STAT3 Silencing and TLR7/8 Pathway Activation Repolarize and Suppress Myeloid-Derived Suppressor Cells From Breast Cancer Patients. Front. Immunol. 2020, 11, 613215. [Google Scholar] [CrossRef]
- Deng, Y.; Yang, J.; Qian, J.; Liu, R.; Huang, E.; Wang, Y.; Luo, F.; Chu, Y. TLR1/TLR2 signaling blocks the suppression of monocytic myeloid-derived suppressor cell by promoting its differentiation into M1-type macrophage. Mol. Immunol. 2019, 112, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xie, Y.; Xiong, Y.; Liu, S.; Qiu, C.; Zhu, Z.; Mao, H.; Yu, M.; Wang, X. TLR 7/8 agonist reverses oxaliplatin resistance in colorectal cancer via directing the myeloid-derived suppressor cells to tumoricidal M1-macrophages. Cancer Lett. 2020, 469, 173–185. [Google Scholar] [CrossRef]
- Adzavon, Y.M.; Zhao, P.; Lv, B.; Liu, M.; Zhang, X.; Xie, F.; Yang, L.; Shang, L.; Zhang, M.; Li, Q.; et al. TLR7 and TLR8 agonist resiquimod (R848) differently regulates MIF expression in cells and organs. Cytokine 2017, 97, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Won, H.; Moreira, D.; Gao, C.; Duttagupta, P.; Zhao, X.; Manuel, E.; Diamond, D.; Yuan, Y.C.; Liu, Z.; Jones, J.; et al. TLR9 expression and secretion of LIF by prostate cancer cells stimulates accumulation and activity of polymorphonuclear MDSCs. J. Leukoc. Biol. 2017, 102, 423–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ubil, E.; Caskey, L.; Holtzhausen, A.; Hunter, D.; Story, C.; Earp, H.S. Tumor-secreted Pros1 inhibits macrophage M1 polarization to reduce antitumor immune response. J. Clin. Investig. 2018, 128, 2356–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, S.; Kohanbash, G.; Liu, S.J.; Alvarado, B.; Carrera, D.; Bhaduri, A.; Watchmaker, P.B.; Yagnik, G.; Di Lullo, E.; Malatesta, M.; et al. Single-cell profiling of human gliomas reveals macrophage ontogeny as a basis for regional differences in macrophage activation in the tumor microenvironment. Genome Biol. 2017, 18, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Yan, L.; Wang, G.J.; Jin, R. Resistin effects on pancreatic cancer progression and chemoresistance are mediated through its receptors CAP1 and TLR4. J. Cell Physiol. 2019, 234, 9457–9466. [Google Scholar] [CrossRef]
- Gong, W.J.; Liu, J.Y.; Yin, J.Y.; Cui, J.J.; Xiao, D.; Zhuo, W.; Luo, C.; Liu, R.J.; Li, X.; Zhang, W.; et al. Resistin facilitates metastasis of lung adenocarcinoma through the TLR4/Src/EGFR/PI3K/NF-kappaB pathway. Cancer Sci. 2018, 109, 2391–2400. [Google Scholar] [CrossRef]
- Reichel, D.; Tripathi, M.; Perez, J.M. Biological Effects of Nanoparticles on Macrophage Polarization in the Tumor Microenvironment. Nanotheranostics 2019, 3, 66–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Mu, R.; Wang, Z.; Xing, P.; Zhang, J.; Dong, L.; Wang, C. A toll-like receptor agonist mimicking microbial signal to generate tumor-suppressive macrophages. Nat. Commun. 2019, 10, 2272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Y.; Li, J.; Zhu, X.; Liu, Y.; Wang, X.; Wang, H.; Yao, Y.; Gao, Y.; Chen, Z. Development of Toll-like Receptor Agonist-Loaded Nanoparticles as Precision Immunotherapy for Reprogramming Tumor-Associated Macrophages. ACS Appl. Mater. Interfaces 2021, 13, 24442–24452. [Google Scholar] [CrossRef] [PubMed]
- Nouri, Y.; Weinkove, R.; Perret, R. T-cell intrinsic Toll-like receptor signaling: Implications for cancer immunotherapy and CAR T-cells. J. Immunother. Cancer 2021, 9, e003065. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, X.; Sanders, K.L.; Edwards, J.L.; Ye, J.; Si, F.; Gao, A.; Huang, L.; Hsueh, E.C.; Ford, D.A.; et al. TLR8-Mediated Metabolic Control of Human Treg Function: A Mechanistic Target for Cancer Immunotherapy. Cell Metab. 2019, 29, 103–123.e5. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Wu, M.; Liu, S.; Shang, W.; Li, R.; Xu, J.; Huang, L.; Wang, F. Glucose metabolism characteristics and TLR8-mediated metabolic control of CD4(+) Treg cells in ovarian cancer cells microenvironment. Cell Death Dis. 2021, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Chodaczek, G.; Pagni, P.P.; Christoffersson, G.; Ratliff, S.S.; Toporkiewicz, M.; Wegrzyn, A.S.; von Herrath, M. The effect of Toll-like receptor stimulation on the motility of regulatory T cells. J. Autoimmun. 2021, 116, 102563. [Google Scholar] [CrossRef]
- Lai, Y.; Weng, J.; Wei, X.; Qin, L.; Lai, P.; Zhao, R.; Jiang, Z.; Li, B.; Lin, S.; Wang, S.; et al. Toll-like receptor 2 costimulation potentiates the antitumor efficacy of CAR T Cells. Leukemia 2018, 32, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Doorduijn, E.M.; Sluijter, M.; Salvatori, D.C.; Silvestri, S.; Maas, S.; Arens, R.; Ossendorp, F.; van der Burg, S.H.; van Hall, T. CD4(+) T Cell and NK Cell Interplay Key to Regression of MHC Class I(low) Tumors upon TLR7/8 Agonist Therapy. Cancer Immunol. Res. 2017, 5, 642–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichterman, J.N.; Reddy, S.M. Mast Cells: A New Frontier for Cancer Immunotherapy. Cells 2021, 10, 1270. [Google Scholar] [CrossRef] [PubMed]
- Dudeck, A.; Koberle, M.; Goldmann, O.; Meyer, N.; Dudeck, J.; Lemmens, S.; Rohde, M.; Roldan, N.G.; Dietze-Schwonberg, K.; Orinska, Z.; et al. Mast cells as protectors of health. J. Allergy Clin. Immunol. 2019, 144, S4–S18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, P.; Zhou, Y.; Yang, Y.; Wu, J.; Zhou, H.; Quan, W.; Sun, J.; Yao, Y.; Shang, A.; Gu, C.; et al. Myeloid cell-derived LL-37 promotes lung cancer growth by activating Wnt/beta-catenin signaling. Theranostics 2019, 9, 2209–2223. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Jing, Y.Y.; Gao, L.; Li, R.; Yang, X.; Pan, X.R.; Yang, Y.; Meng, Y.; Hou, X.J.; Zhao, Q.D.; et al. Lipopolysaccharide induces the differentiation of hepatic progenitor cells into myofibroblasts constitutes the hepatocarcinogenesis-associated microenvironment. Cell Death Differ. 2020, 27, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Khajeh Alizadeh Attar, M.; Anwar, M.A.; Eskian, M.; Keshavarz-Fathi, M.; Choi, S.; Rezaei, N. Basic understanding and therapeutic approaches to target toll-like receptors in cancerous microenvironment and metastasis. Med. Res. Rev. 2018, 38, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Ridnour, L.A.; Cheng, R.Y.; Switzer, C.H.; Heinecke, J.L.; Ambs, S.; Glynn, S.; Young, H.A.; Trinchieri, G.; Wink, D.A. Molecular pathways: Toll-like receptors in the tumor microenvironment—Poor prognosis or new therapeutic opportunity. Clin. Cancer Res. 2013, 19, 1340–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Li, S.; Yang, H. Roles of Toll-Like Receptor 3 in Human Tumors. Front. Immunol. 2021, 12, 667454. [Google Scholar] [CrossRef]
- Husseinzadeh, N.; Davenport, S.M. Role of toll-like receptors in cervical, endometrial and ovarian cancers: A review. Gynecol. Oncol. 2014, 135, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Jeon, S.B.; Yoon, H.J.; Choi, B.K.; Kim, S.S.; Oshima, M.; Park, E.J. Glial TLR2-driven innate immune responses and CD8(+) T cell activation against brain tumor. Glia 2019, 67, 1179–1195. [Google Scholar] [CrossRef]
- Qian, J.; Luo, F.; Yang, J.; Liu, J.; Liu, R.; Wang, L.; Wang, C.; Deng, Y.; Lu, Z.; Wang, Y.; et al. TLR2 Promotes Glioma Immune Evasion by Downregulating MHC Class II Molecules in Microglia. Cancer Immunol. Res. 2018, 6, 1220–1233. [Google Scholar] [CrossRef] [Green Version]
- Fehri, E.; Ennaifer, E.; Bel Haj Rhouma, R.; Guizani-Tabbane, L.; Guizani, I.; Boubaker, S. The role of Toll-like receptor 9 in gynecologic cancer. Curr. Res. Transl. Med. 2016, 64, 155–159. [Google Scholar] [CrossRef]
- Alvarado, A.G.; Thiagarajan, P.S.; Mulkearns-Hubert, E.E.; Silver, D.J.; Hale, J.S.; Alban, T.J.; Turaga, S.M.; Jarrar, A.; Reizes, O.; Longworth, M.S.; et al. Glioblastoma Cancer Stem Cells Evade Innate Immune Suppression of Self-Renewal through Reduced TLR4 Expression. Cell Stem Cell 2017, 20, 450–461.e4. [Google Scholar] [CrossRef] [Green Version]
- Da Cruz, L.L.P.; de Souza, P.O.; Dal Pra, M.; Falchetti, M.; de Abreu, A.M.; Azambuja, J.H.; Bertoni, A.P.S.; Paz, A.H.R.; Araujo, A.B.; Visioli, F.; et al. TLR4 expression and functionality are downregulated in glioblastoma cells and in tumor-associated macrophages: A new mechanism of immune evasion? Biochim. Biophys. Acta Mol. Basis. Dis. 2021, 1867, 166155. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, Z.; Xiao, X.Y.; Zhang, Z.Y.; Gao, S.L.; Lu, C.; Zuo, L.; Zhang, L.F. Toll-like receptor 2 (TLR2) is a candidate prognostic factor in testicular germ cell tumors as well as an indicator of immune function in the tumor microenvironment. Bioengineered 2021, 12, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Scatozza, F.; D’Amore, A.; Fontanella, R.A.; De Cesaris, P.; Marampon, F.; Padula, F.; Ziparo, E.; Riccioli, A.; Filippini, A. Toll-Iike Receptor-3 Activation Enhances Malignant Traits in Human Breast Cancer Cells Through Hypoxia-inducible Factor-1alpha. Anticancer Res. 2020, 40, 5379–5391. [Google Scholar] [CrossRef] [PubMed]
- Kohtz, P.D.; Halpern, A.L.; Eldeiry, M.A.; Hazel, K.; Kalatardi, S.; Ao, L.; Meng, X.; Reece, T.B.; Fullerton, D.A.; Weyant, M.J. Toll-Like Receptor-4 Is a Mediator of Proliferation in Esophageal Adenocarcinoma. Ann. Thorac. Surg. 2019, 107, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Fels Elliott, D.R.; Perner, J.; Li, X.; Symmons, M.F.; Verstak, B.; Eldridge, M.; Bower, L.; O’Donovan, M.; Gay, N.J.; Consortium, O.; et al. Impact of mutations in Toll-like receptor pathway genes on esophageal carcinogenesis. PLoS Genet. 2017, 13, e1006808. [Google Scholar] [CrossRef] [Green Version]
- Zu, Y.; Ping, W.; Deng, T.; Zhang, N.; Fu, X.; Sun, W. Lipopolysaccharide-induced toll-like receptor 4 signaling in esophageal squamous cell carcinoma promotes tumor proliferation and regulates inflammatory cytokines expression. Dis. Esophagus 2017, 30, 1–8. [Google Scholar] [CrossRef]
- Mohamed, F.E.; Al-Jehani, R.M.; Minogue, S.S.; Andreola, F.; Winstanley, A.; Olde Damink, S.W.; Habtesion, A.; Malago, M.; Davies, N.; Luong, T.V.; et al. Effect of toll-like receptor 7 and 9 targeted therapy to prevent the development of hepatocellular carcinoma. Liver Int. 2015, 35, 1063–1076. [Google Scholar] [CrossRef]
- Loh, J.J.; Li, T.W.; Zhou, L.; Wong, T.L.; Liu, X.; Ma, V.W.S.; Lo, C.M.; Man, K.; Lee, T.K.; Ning, W.; et al. FSTL1 Secreted by Activated Fibroblasts Promotes Hepatocellular Carcinoma Metastasis and Stemness. Cancer Res. 2021, 81, 5692–5705. [Google Scholar] [CrossRef]
- Zhou, S.; Du, R.; Wang, Z.; Shen, W.; Gao, R.; Jiang, S.; Fang, Y.; Shi, Y.; Chang, A.; Liu, L.; et al. TLR4 increases the stemness and is highly expressed in relapsed human hepatocellular carcinoma. Cancer Med. 2019, 8, 2325–2337. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Shapiro, B.; Vucic, E.A.; Vogt, S.; Bar-Sagi, D. Tumor Cell-Derived IL1beta Promotes Desmoplasia and Immune Suppression in Pancreatic Cancer. Cancer Res. 2020, 80, 1088–1101. [Google Scholar] [CrossRef] [Green Version]
- Rapoport, B.L.; Steel, H.C.; Theron, A.J.; Heyman, L.; Smit, T.; Ramdas, Y.; Anderson, R. High Mobility Group Box 1 in Human Cancer. Cells 2020, 9, 1664. [Google Scholar] [CrossRef]
- Bianchi, F.; Milione, M.; Casalini, P.; Centonze, G.; Le Noci, V.M.; Storti, C.; Alexiadis, S.; Truini, M.; Sozzi, G.; Pastorino, U.; et al. Toll-like receptor 3 as a new marker to detect high risk early stage Non-Small-Cell Lung Cancer patients. Sci. Rep. 2019, 9, 14288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Wang, J.; Wei, F.; Zhao, N.; Yang, F.; Ren, X. Expression of TLR4 in Non-Small Cell Lung Cancer Is Associated with PD-L1 and Poor Prognosis in Patients Receiving Pulmonectomy. Front. Immunol. 2017, 8, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Zhou, R.; Wang, H.; Li, W.; Pan, M.; Yao, X.; Zhan, W.; Yang, S.; Xu, L.; Ding, Y.; et al. Gut microbiota-stimulated cathepsin K secretion mediates TLR4-dependent M2 macrophage polarization and promotes tumor metastasis in colorectal cancer. Cell Death. Differ. 2019, 26, 2447–2463. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liao, M.; Wang, J. TLR4 signaling in the development of colitis-associated cancer and its possible interplay with microRNA-155. Cell Commun. Signal 2021, 19, 90. [Google Scholar] [CrossRef]
- Nie, M.; Yang, L.; Bi, X.; Wang, Y.; Sun, P.; Yang, H.; Liu, P.; Li, Z.; Xia, Y.; Jiang, W. Neutrophil Extracellular Traps Induced by IL8 Promote Diffuse Large B-cell Lymphoma Progression via the TLR9 Signaling. Clin. Cancer Res. 2019, 25, 1867–1879. [Google Scholar] [CrossRef]
- Muniz-Bongers, L.R.; McClain, C.B.; Saxena, M.; Bongers, G.; Merad, M.; Bhardwaj, N. MMP2 and TLRs modulate immune responses in the tumor microenvironment. JCI Insight 2021, 6, e144913. [Google Scholar] [CrossRef]
- Chen, J.; Qiao, Y.; Chen, G.; Chang, C.; Dong, H.; Tang, B.; Cheng, X.; Liu, X.; Hua, Z. Salmonella flagella confer anti-tumor immunological effect via activating Flagellin/TLR5 signalling within tumor microenvironment. Acta Pharm. Sin. B 2021, 11, 3165–3177. [Google Scholar] [CrossRef]
- Roychowdhury, A.; Jondhale, M.; Saldanha, E.; Ghosh, D.; Kumar Panda, C.; Chandrani, P.; Mukherjee, N. Landscape of toll-like receptors expression in tumor microenvironment of triple negative breast cancer (TNBC): Distinct roles of TLR4 and TLR8. Gene 2021, 792, 145728. [Google Scholar] [CrossRef]
- Paone, A.; Galli, R.; Gabellini, C.; Lukashev, D.; Starace, D.; Gorlach, A.; De Cesaris, P.; Ziparo, E.; Del Bufalo, D.; Sitkovsky, M.V.; et al. Toll-like receptor 3 regulates angiogenesis and apoptosis in prostate cancer cell lines through hypoxia-inducible factor 1 alpha. Neoplasia 2010, 12, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Paone, A.; Starace, D.; Galli, R.; Padula, F.; De Cesaris, P.; Filippini, A.; Ziparo, E.; Riccioli, A. Toll-like receptor 3 triggers apoptosis of human prostate cancer cells through a PKC-alpha-dependent mechanism. Carcinogenesis 2008, 29, 1334–1342. [Google Scholar] [CrossRef]
- Dietsch, G.N.; Lu, H.; Yang, Y.; Morishima, C.; Chow, L.Q.; Disis, M.L.; Hershberg, R.M. Coordinated Activation of Toll-Like Receptor8 (TLR8) and NLRP3 by the TLR8 Agonist, VTX-2337, Ignites Tumoricidal Natural Killer Cell Activity. PLoS ONE 2016, 11, e0148764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.N.; Jin, S.M.; Shin, H.S.; Lim, Y.T. Chemical Strategies to Enhance the Therapeutic Efficacy of Toll-like Receptor Agonist Based Cancer Immunotherapy. Acc. Chem. Res. 2020, 53, 2081–2093. [Google Scholar] [CrossRef] [PubMed]
- Patra, M.C.; Choi, S. Recent progress in the development of Toll-like receptor (TLR) antagonists. Expert Opin. Ther. Pat. 2016, 26, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Oblak, A.; Jerala, R. Toll-like receptor 4 activation in cancer progression and therapy. Clin. Dev. Immunol. 2011, 2011, 609579. [Google Scholar] [CrossRef] [Green Version]
- Talukdar, A.; Ganguly, D.; Roy, S.; Das, N.; Sarkar, D. Structural Evolution and Translational Potential for Agonists and Antagonists of Endosomal Toll-like Receptors. J. Med. Chem. 2021, 64, 8010–8041. [Google Scholar] [CrossRef]
- Huang, S.; Mei, H.; Zhang, D.; Shi, T.; Chen, L.; Kuang, Z.; Heng, Y.; Pan, X.; Lu, L. Subtle differences in chemical pattern between human toll-like receptor 8 agonists and antagonists: Emerging chemical patterns analysis. Chem. Biol. Drug Des. 2019, 94, 1824–1834. [Google Scholar] [CrossRef]
- Kaczanowska, S.; Joseph, A.M.; Davila, E. TLR agonists: Our best frenemy in cancer immunotherapy. J. Leukoc. Biol. 2013, 93, 847–863. [Google Scholar] [CrossRef] [Green Version]
- Pahlavanneshan, S.; Sayadmanesh, A.; Ebrahimiyan, H.; Basiri, M. Toll-Like Receptor-Based Strategies for Cancer Immunotherapy. J. Immunol. Res. 2021, 2021, 9912188. [Google Scholar] [CrossRef]
- Liu, C.; Han, C.; Liu, J. The Role of Toll-Like Receptors in Oncotherapy. Oncol. Res. 2019, 27, 965–978. [Google Scholar] [CrossRef]
- Meyer, T.; Stockfleth, E. Clinical investigations of Toll-like receptor agonists. Expert Opin. Investig. Drugs 2008, 17, 1051–1065. [Google Scholar] [CrossRef]
- Shetab Boushehri, M.A.; Lamprecht, A. TLR4-Based Immunotherapeutics in Cancer: A Review of the Achievements and Shortcomings. Mol. Pharm. 2018, 15, 4777–4800. [Google Scholar] [CrossRef] [PubMed]
- Bhagchandani, S.; Johnson, J.A.; Irvine, D.J. Evolution of Toll-like receptor 7/8 agonist therapeutics and their delivery approaches: From antiviral formulations to vaccine adjuvants. Adv. Drug Deliv. Rev. 2021, 175, 113803. [Google Scholar] [CrossRef] [PubMed]
- Novak, N.; Yu, C.F.; Bieber, T.; Allam, J.P. Toll-like receptor 7 agonists and skin. Drug News Perspect. 2008, 21, 158–165. [Google Scholar] [PubMed]
- Seya, T.; Shime, H.; Takeda, Y.; Tatematsu, M.; Takashima, K.; Matsumoto, M. Adjuvant for vaccine immunotherapy of cancer—Focusing on Toll-like receptor 2 and 3 agonists for safely enhancing antitumor immunity. Cancer Sci. 2015, 106, 1659–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, F.; Pretto, S.; Tagliabue, E.; Balsari, A.; Sfondrini, L. Exploiting poly(I:C) to induce cancer cell apoptosis. Cancer Biol. Ther. 2017, 18, 747–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Xue, Y.; Pan, Y.; Yao, A.; Wang, G.; Li, D.; Wang, T.; Zhao, S.; Hou, Y. Toll-like receptor 3 agonist poly I:C reinforces the potency of cytotoxic chemotherapy via the TLR3-UNC93B1-IFN-beta signaling axis in paclitaxel-resistant colon cancer. J. Cell Physiol. 2019, 234, 7051–7061. [Google Scholar] [CrossRef]
- Bagaev, A.; Pichugin, A.; Nelson, E.L.; Agadjanyan, M.G.; Ghochikyan, A.; Ataullakhanov, R.I. Anticancer Mechanisms in Two Murine Bone Marrow-Derived Dendritic Cell Subsets Activated with TLR4 Agonists. J. Immunol. 2018, 200, 2656–2669. [Google Scholar] [CrossRef] [Green Version]
- Theile, D.; Wagner, L.; Haefeli, W.E.; Weiss, J. In vitro evidence suggesting that the toll-like receptor 7 and 8 agonist resiquimod (R-848) unlikely affects drug levels of co-administered compounds. Eur. J. Pharm. Sci. 2021, 162, 105826. [Google Scholar] [CrossRef]
- Melisi, D.; Frizziero, M.; Tamburrino, A.; Zanotto, M.; Carbone, C.; Piro, G.; Tortora, G. Toll-Like Receptor 9 Agonists for Cancer Therapy. Biomedicines 2014, 2, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Krieg, A.M. Therapeutic potential of Toll-like receptor 9 activation. Nat. Rev. Drug Discov. 2006, 5, 471–484. [Google Scholar] [CrossRef]
- Holtick, U.; Scheulen, M.E.; von Bergwelt-Baildon, M.S.; Weihrauch, M.R. Toll-like receptor 9 agonists as cancer therapeutics. Expert Opin. Investig. Drugs 2011, 20, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, M.; Smolka, G. Clausal Tableaux for Hybrid PDL. In Electronic Notes in Theoretical Computer Science; Elsevier: Amsterdam, The Netherlands, 2011; Volume 278, pp. 99–113. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; An, M.; Xi, J.; Liu, H. Targeting CpG Adjuvant to Lymph Node via Dextran Conjugate Enhances Antitumor Immunotherapy. Bioconjug. Chem. 2017, 28, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Gallotta, M.; Assi, H.; Degagne, E.; Kannan, S.K.; Coffman, R.L.; Guiducci, C. Inhaled TLR9 Agonist Renders Lung Tumors Permissive to PD-1 Blockade by Promoting Optimal CD4(+) and CD8(+) T-cell Interplay. Cancer Res. 2018, 78, 4943–4956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieg, A.M. Toll-like receptor 9 (TLR9) agonists in the treatment of cancer. Oncogene 2008, 27, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.L.; Sagiv-Barfi, I.; Neuhofer, P.; Czerwinski, D.K.; Artandi, S.E.; Bertozzi, C.R.; Levy, R.; Cochran, J.R. Systemic delivery of a targeted synthetic immunostimulant transforms the immune landscape for effective tumor regression. Cell Chem. Biol. 2022, 29, 451–462.e8. [Google Scholar] [CrossRef]
- Ackerman, S.E.; Pearson, C.I.; Gregorio, J.D.; Gonzalez, J.C.; Kenkel, J.A.; Hartmann, F.J.; Luo, A.; Ho, P.Y.; LeBlanc, H.; Blum, L.K.; et al. Immune-stimulating antibody conjugates elicit robust myeloid activation and durable antitumor immunity. Nat. Cancer 2021, 2, 18–33. [Google Scholar] [CrossRef]
- Liu, Y.; Li, C.; Xia, H.; Bi, J.; Guan, R.; Du, X.; Zhang, H.; He, Z.; Wang, Y.; Liu, H. An injectable superior depot of Telratolimod inhibits post-surgical tumor recurrence and distant metastases. Acta Biomater. 2022, 141, 132–139. [Google Scholar] [CrossRef]
- Mullins, S.R.; Vasilakos, J.P.; Deschler, K.; Grigsby, I.; Gillis, P.; John, J.; Elder, M.J.; Swales, J.; Timosenko, E.; Cooper, Z.; et al. Intratumoral immunotherapy with TLR7/8 agonist MEDI9197 modulates the tumor microenvironment leading to enhanced activity when combined with other immunotherapies. J. Immunother. Cancer 2019, 7, 244. [Google Scholar] [CrossRef]
- Kapp, K.; Volz, B.; Curran, M.A.; Oswald, D.; Wittig, B.; Schmidt, M. EnanDIM—A novel family of L-nucleotide-protected TLR9 agonists for cancer immunotherapy. J. Immunother. Cancer 2019, 7, 5. [Google Scholar] [CrossRef]
- Wittig, B.; Schmidt, M.; Scheithauer, W.; Schmoll, H.J. MGN1703, an immunomodulator and toll-like receptor 9 (TLR-9) agonist: From bench to bedside. Crit. Rev. Oncol. Hematol. 2015, 94, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Sabree, S.A.; Voigt, A.P.; Blackwell, S.E.; Vishwakarma, A.; Chimenti, M.S.; Salem, A.K.; Weiner, G.J. Direct and indirect immune effects of CMP-001, a virus-like particle containing a TLR9 agonist. J. Immunother. Cancer 2021, 9, 1699. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jiang, W.; Zhu, F.; Mao, X.; Agrawal, S. Modulation of the tumor microenvironment by intratumoral administration of IMO-2125, a novel TLR9 agonist, for cancer immunotherapy. Int. J. Oncol. 2018, 53, 1193–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zom, G.G.; Willems, M.; Khan, S.; van der Sluis, T.C.; Kleinovink, J.W.; Camps, M.G.M.; van der Marel, G.A.; Filippov, D.V.; Melief, C.J.M.; Ossendorp, F. Novel TLR2-binding adjuvant induces enhanced T cell responses and tumor eradication. J. Immunother. Cancer 2018, 6, 146. [Google Scholar] [CrossRef] [Green Version]
- Morin, M.D.; Wang, Y.; Jones, B.T.; Mifune, Y.; Su, L.; Shi, H.; Moresco, E.M.Y.; Zhang, H.; Beutler, B.; Boger, D.L. Diprovocims: A New and Exceptionally Potent Class of Toll-like Receptor Agonists. J. Am. Chem. Soc. 2018, 140, 14440–14454. [Google Scholar] [CrossRef] [PubMed]
- Seya, T.; Takeda, Y.; Matsumoto, M. A Toll-like receptor 3 (TLR3) agonist ARNAX for therapeutic immunotherapy. Adv. Drug Deliv. Rev. 2019, 147, 37–43. [Google Scholar] [CrossRef]
- Takeda, Y.; Yoshida, S.; Takashima, K.; Ishii-Mugikura, N.; Shime, H.; Seya, T.; Matsumoto, M. Vaccine immunotherapy with ARNAX induces tumor-specific memory T cells and durable anti-tumor immunity in mouse models. Cancer Sci. 2018, 109, 2119–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, Y.; Kataoka, K.; Yamagishi, J.; Ogawa, S.; Seya, T.; Matsumoto, M. A TLR3-Specific Adjuvant Relieves Innate Resistance to PD-L1 Blockade without Cytokine Toxicity in Tumor Vaccine Immunotherapy. Cell Rep. 2017, 19, 1874–1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mett, V.; Komarova, E.A.; Greene, K.; Bespalov, I.; Brackett, C.; Gillard, B.; Gleiberman, A.S.; Toshkov, I.A.; Aygun-Sunar, S.; Johnson, C.; et al. Mobilan: A recombinant adenovirus carrying Toll-like receptor 5 self-activating cassette for cancer immunotherapy. Oncogene 2018, 37, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, B.A.; Matheny, C.J.; Burns, O.; Struemper, H.; Wang, X.; Washburn, M.L. Safety, Pharmacokinetics, and Pharmacodynamics of the TLR4 Agonist GSK1795091 in Healthy Individuals: Results from a Randomized, Double-blind, Placebo-controlled, Ascending Dose Study. Clin. Ther. 2020, 42, 1519–1534.e33. [Google Scholar] [CrossRef] [PubMed]
- Slingluff, C.L.; Zarour, H.M.; Tawbi, H.A.; Kirkwood, J.M.; Postow, M.A.; Friedlander, P.; Devoe, C.E.; Gaughan, E.M.; Mauldin, I.S.; Olson, W.C.; et al. A phase 1 study of NY-ESO-1 vaccine + anti-CTLA4 antibody Ipilimumab (IPI) in patients with unresectable or metastatic melanoma. Oncoimmunology 2021, 10, 1898105. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Kageyama, S.; Miyahara, Y.; Okayama, T.; Kokura, S.; Wang, L.; Sato, E.; Yagita, H.; Itoh, Y.; Shiku, H. Safety and antibody immune response of CHP-NY-ESO-1 vaccine combined with poly-ICLC in advanced or recurrent esophageal cancer patients. Cancer Immunol. Immunother. 2021, 70, 3081–3091. [Google Scholar] [CrossRef]
- Shayan, G.; Kansy, B.A.; Gibson, S.P.; Srivastava, R.M.; Bryan, J.K.; Bauman, J.E.; Ohr, J.; Kim, S.; Duvvuri, U.; Clump, D.A.; et al. Phase Ib Study of Immune Biomarker Modulation with Neoadjuvant Cetuximab and TLR8 Stimulation in Head and Neck Cancer to Overcome Suppressive Myeloid Signals. Clin. Cancer Res. 2018, 24, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, S.; Britten, C.D.; Chin, S.; Garrett-Mayer, E.; Cloud, C.A.; Li, M.; Scurti, G.; Salem, M.L.; Nelson, M.H.; Thomas, M.B.; et al. Vaccination with poly(IC:LC) and peptide-pulsed autologous dendritic cells in patients with pancreatic cancer. J. Hematol. Oncol. 2017, 10, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meneveau, M.O.; Petroni, G.R.; Salerno, E.P.; Lynch, K.T.; Smolkin, M.; Woodson, E.; Chianese-Bullock, K.A.; Olson, W.C.; Deacon, D.; Patterson, J.W.; et al. Immunogenicity in humans of a transdermal multipeptide melanoma vaccine administered with or without a TLR7 agonist. J. Immunother. Cancer 2021, 9, e002214. [Google Scholar] [CrossRef] [PubMed]
- Salazar, L.G.; Lu, H.; Reichow, J.L.; Childs, J.S.; Coveler, A.L.; Higgins, D.M.; Waisman, J.; Allison, K.H.; Dang, Y.; Disis, M.L. Topical Imiquimod Plus Nab-paclitaxel for Breast Cancer Cutaneous Metastases: A Phase 2 Clinical Trial. JAMA. Oncol. 2017, 3, 969–973. [Google Scholar] [CrossRef]
- Patel, S.P.; Petroni, G.R.; Roszik, J.; Olson, W.C.; Wages, N.A.; Chianese-Bullock, K.A.; Smolkin, M.; Varhegyi, N.; Gaughan, E.; Smith, K.T.; et al. Phase I/II trial of a long peptide vaccine (LPV7) plus toll-like receptor (TLR) agonists with or without incomplete Freund’s adjuvant (IFA) for resected high-risk melanoma. J. Immunother. Cancer 2021, 9, e003220. [Google Scholar] [CrossRef] [PubMed]
- Pavlick, A.; Blazquez, A.B.; Meseck, M.; Lattanzi, M.; Ott, P.A.; Marron, T.U.; Holman, R.M.; Mandeli, J.; Salazar, A.M.; McClain, C.B.; et al. Combined Vaccination with NY-ESO-1 Protein, Poly-ICLC, and Montanide Improves Humoral and Cellular Immune Responses in Patients with High-Risk Melanoma. Cancer Immunol. Res. 2020, 8, 70–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Suh, J.; Surh, Y.J.; Na, H.K. Regulation of the tumor suppressor PTEN by natural anticancer compounds. Ann. N. Y. Acad. Sci. 2017, 1401, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.Q.M.; Morishima, C.; Eaton, K.D.; Baik, C.S.; Goulart, B.H.; Anderson, L.N.; Manjarrez, K.L.; Dietsch, G.N.; Bryan, J.K.; Hershberg, R.M.; et al. Phase Ib Trial of the Toll-like Receptor 8 Agonist, Motolimod (VTX-2337), Combined with Cetuximab in Patients with Recurrent or Metastatic SCCHN. Clin. Cancer Res. 2017, 23, 2442–2450. [Google Scholar] [CrossRef]
- Carbone, C.; Piro, G.; Agostini, A.; Delfino, P.; De Sanctis, F.; Nasca, V.; Spallotta, F.; Sette, C.; Martini, M.; Ugel, S.; et al. Intratumoral injection of TLR9 agonist promotes an immunopermissive microenvironment transition and causes cooperative antitumor activity in combination with anti-PD1 in pancreatic cancer. J. Immunother. Cancer 2021, 9, e002876. [Google Scholar] [CrossRef]
- Karime, C.; Wang, J.; Woodhead, G.; Mody, K.; Hennemeyer, C.T.; Borad, M.J.; Mahadevan, D.; Chandana, S.R.; Babiker, H. Tilsotolimod: An investigational synthetic toll-like receptor 9 (TLR9) agonist for the treatment of refractory solid tumors and melanoma. Expert. Opin. Investig. Drugs. 2022, 31, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Miller, N.J.; Lu, H.; Longino, N.V.; Ibrani, D.; Shinohara, M.M.; Byrd, D.R.; Parvathaneni, U.; Kulikauskas, R.; Ter Meulen, J.; et al. Intratumoral G100, a TLR4 Agonist, Induces Antitumor Immune Responses and Tumor Regression in Patients with Merkel Cell Carcinoma. Clin. Cancer Res. 2019, 25, 1185–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.J.; Reagan, P.M.; Bartlett, N.L.; Gordon, L.I.; Friedberg, J.W.; Czerwinski, D.K.; Long, S.R.; Hoppe, R.T.; Janssen, R.; Candia, A.F.; et al. In Situ Vaccination with a TLR9 Agonist and Local Low-Dose Radiation Induces Systemic Responses in Untreated Indolent Lymphoma. Cancer Discov. 2018, 8, 1258–1269. [Google Scholar] [CrossRef] [Green Version]
- Monk, B.J.; Brady, M.F.; Aghajanian, C.; Lankes, H.A.; Rizack, T.; Leach, J.; Fowler, J.M.; Higgins, R.; Hanjani, P.; Morgan, M.; et al. A phase 2, randomized, double-blind, placebo- controlled study of chemo-immunotherapy combination using motolimod with pegylated liposomal doxorubicin in recurrent or persistent ovarian cancer: A Gynecologic Oncology Group partners study. Ann. Oncol. 2017, 28, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Grewal, E.P.; Erskine, C.L.; Nevala, W.K.; Allred, J.B.; Strand, C.A.; Kottschade, L.A.; Block, M.S. Peptide vaccine with glucopyranosyl lipid A—Stable oil-in-water emulsion for patients with resected melanoma. Immunotherapy 2020, 12, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Brody, J.D.; Ai, W.Z.; Czerwinski, D.K.; Torchia, J.A.; Levy, M.; Advani, R.H.; Kim, Y.H.; Hoppe, R.T.; Knox, S.J.; Shin, L.K.; et al. In situ vaccination with a TLR9 agonist induces systemic lymphoma regression: A phase I/II study. J. Clin. Oncol. 2010, 28, 4324–4332. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Gratzinger, D.; Harrison, C.; Brody, J.D.; Czerwinski, D.K.; Ai, W.Z.; Levy, R. In situ vaccination against mycosis fungoides by intratumoral injection of a TLR9 agonist combined with radiation: A phase 1/2 study. Blood J. Am. Soc. Hematol. 2012, 119, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Li, J.; Gu, P.; Fan, X. The application of nanoparticles in cancer immunotherapy: Targeting tumor microenvironment. Bioact. Mater. 2021, 6, 1973–1987. [Google Scholar] [CrossRef]
- Jin, S.M.; Lee, S.N.; Kim, J.E.; Yoo, Y.J.; Song, C.; Shin, H.S.; Phuengkham, H.; Lee, C.H.; Um, S.H.; Lim, Y.T. Overcoming Chemoimmunotherapy-Induced Immunosuppression by Assemblable and Depot Forming Immune Modulating Nanosuspension. Adv. Sci. 2021, 8, e2102043. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Gu, P.; Yang, Y.; Yu, L.; Jiang, Z.; Li, J.; Le, Y.; Chen, Y.; Ba, Q.; Wang, H. Mesoporous silica nanoparticles inflame tumors to overcome anti-PD-1 resistance through TLR4-NFkappaB axis. J. Immunother. Cancer 2021, 9, e002508. [Google Scholar] [CrossRef]
- Cejudo-Guillen, M.; Ramiro-Gutierrez, M.L.; Labrador-Garrido, A.; Diaz-Cuenca, A.; Pozo, D. Nanoporous silica microparticle interaction with toll-like receptor agonists in macrophages. Acta Biomater. 2012, 8, 4295–4303. [Google Scholar] [CrossRef] [PubMed]
- Schmid, D.; Park, C.G.; Hartl, C.A.; Subedi, N.; Cartwright, A.N.; Puerto, R.B.; Zheng, Y.; Maiarana, J.; Freeman, G.J.; Wucherpfennig, K.W.; et al. T cell-targeting nanoparticles focus delivery of immunotherapy to improve antitumor immunity. Nat. Commun. 2017, 8, 1747. [Google Scholar] [CrossRef] [Green Version]
- Yoo, Y.J.; Lee, C.H.; Park, S.H.; Lim, Y.T. Nanoparticle-based delivery strategies of multifaceted immunomodulatory RNA for cancer immunotherapy. J. Control. Release 2022, 343, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Jiang, C. Delivery strategies for macromolecular drugs in cancer therapy. Acta Pharm. Sin. B 2020, 10, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Raimondo, S.; Urzi, O.; Moschetti, M.; Di Bella, M.A.; Conigliaro, A.; Caccamo, N.; La Manna, M.P.; Fontana, S.; Alessandro, R. Tumor-Derived Small Extracellular Vesicles Induce Pro-Inflammatory Cytokine Expression and PD-L1 Regulation in M0 Macrophages via IL-6/STAT3 and TLR4 Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 2118. [Google Scholar] [CrossRef]
- Domenis, R.; Cifu, A.; Marino, D.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; Curcio, F. Toll-like Receptor-4 Activation Boosts the Immunosuppressive Properties of Tumor Cells-derived Exosomes. Sci. Rep. 2019, 9, 8457. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.F.; Liu, H.; Gao, R.; Zhou, M.; Ma, J.; Zhang, Y.; Zhao, J.; Chen, Y.; Zhang, T.; Huang, F.; et al. Tumor cell-released autophagosomes (TRAPs) promote immunosuppression through induction of M2-like macrophages with increased expression of PD-L1. J. Immunother. Cancer 2018, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Zhang, Q.; Cheng, Y.; Chen, X.; Wang, G.; Shi, M.; Zhang, T.; Cao, Y.; Pan, H.; Zhang, L.; et al. Tumor-derived exosomal HMGB1 fosters hepatocellular carcinoma immune evasion by promoting TIM-1(+) regulatory B cell expansion. J. Immunother. Cancer 2018, 6, 145. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shi, H.; Yuan, X.; Jiang, P.; Qian, H.; Xu, W. Tumor-derived exosomes induce N2 polarization of neutrophils to promote gastric cancer cell migration. Mol. Cancer 2018, 17, 146. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.Q.; Li, P.C.; Pan, N.; Gao, R.; Wen, Z.F.; Zhang, T.Y.; Huang, F.; Wu, F.Y.; Ou, X.L.; Zhang, J.P.; et al. Tumor-released autophagosomes induces CD4(+) T cell-mediated immunosuppression via a TLR2-IL-6 cascade. J. Immunother. Cancer 2019, 7, 178. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, L.; Guo, B.; Deng, J.; Wu, S.; Li, F.; Wang, Y.; Lu, J.; Zhou, Y. Exosomal FMR1-AS1 facilitates maintaining cancer stem-like cell dynamic equilibrium via TLR7/NFkappaB/c-Myc signaling in female esophageal carcinoma. Mol. Cancer 2019, 18, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.; Li, Z.; He, J.; Fu, S.; Duan, Y.; Zhou, Q.; Yan, Y.; Liu, X.; Liu, L.; Feng, C.; et al. Epstein-Barr virus noncoding RNAs from the extracellular vesicles of nasopharyngeal carcinoma (NPC) cells promote angiogenesis via TLR3/RIG-I-mediated VCAM-1 expression. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1201–1213. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, J.; Li, Y.; Xu, F.; Hou, J.; Yang, G.; Zhou, S. Development of a Localized Drug Delivery System with a Step-by-Step Cell Internalization Capacity for Cancer Immunotherapy. ACS Nano 2022, 16, 5778–5794. [Google Scholar] [CrossRef] [PubMed]
- Yuba, E. Liposome-based immunity-inducing systems for cancer immunotherapy. Mol. Immunol. 2018, 98, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Kocabas, B.B.; Almacioglu, K.; Bulut, E.A.; Gucluler, G.; Tincer, G.; Bayik, D.; Gursel, M.; Gursel, I. Dual-adjuvant effect of pH-sensitive liposomes loaded with STING and TLR9 agonists regress tumor development by enhancing Th1 immune response. J. Control Release 2020, 328, 587–595. [Google Scholar] [CrossRef]
- Klauber, T.C.B.; Laursen, J.M.; Zucker, D.; Brix, S.; Jensen, S.S.; Andresen, T.L. Delivery of TLR7 agonist to monocytes and dendritic cells by DCIR targeted liposomes induces robust production of anti-cancer cytokines. Acta Biomater. 2017, 53, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Francian, A.; Widmer, A.; Olsson, T.; Ramirez, M.; Heald, D.; Rasic, K.; Adams, L.; Martinson, H.; Kullberg, M. Delivery of toll-like receptor agonists by complement C3-targeted liposomes activates immune cells and reduces tumour growth. J. Drug Target 2021, 29, 754–760. [Google Scholar] [CrossRef]
- Roses, R.E.; Xu, M.; Koski, G.K.; Czerniecki, B.J. Radiation therapy and Toll-like receptor signaling: Implications for the treatment of cancer. Oncogene 2008, 27, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Walshaw, R.C.; Honeychurch, J.; Choudhury, A.; Illidge, T.M. Toll-Like Receptor Agonists and Radiation Therapy Combinations: An Untapped Opportunity to Induce Anticancer Immunity and Improve Tumor control. Int. J. Radiat. Oncol. Biol. Phys. 2020, 108, 27–37. [Google Scholar] [CrossRef]
- Uthaman, S.; Pillarisetti, S.; Hwang, H.S.; Mathew, A.P.; Huh, K.M.; Rhee, J.H.; Park, I.K. Tumor Microenvironment-Regulating Immunosenescence-Independent Nanostimulant Synergizing with Near-Infrared Light Irradiation for Antitumor Immunity. ACS Appl. Mater. Interfaces 2021, 13, 4844–4852. [Google Scholar] [CrossRef]
- Sagiv-Barfi, I.; Czerwinski, D.K.; Levy, S.; Alam, I.S.; Mayer, A.T.; Gambhir, S.S.; Levy, R. Eradication of spontaneous malignancy by local immunotherapy. Sci. Transl. Med. 2018, 10, eaan4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banstola, A.; Jeong, J.H.; Yook, S. Immunoadjuvants for cancer immunotherapy: A review of recent developments. Acta Biomater. 2020, 114, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Sultan, H.; Salazar, A.M.; Celis, E. Poly-ICLC, a multi-functional immune modulator for treating cancer. Semin. Immunol. 2020, 49, 101414. [Google Scholar] [CrossRef] [PubMed]
- Vermaelen, K. Vaccine Strategies to Improve Anti-cancer Cellular Immune Responses. Front. Immunol. 2019, 10, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Guo, Z. Recent Advances in Toll Like Receptor-Targeting Glycoconjugate Vaccines. Molecules 2018, 23, 1583. [Google Scholar] [CrossRef] [Green Version]
- Kanzler, H.; Barrat, F.J.; Hessel, E.M.; Coffman, R.L. Therapeutic targeting of innate immunity with Toll-like receptor agonists and antagonists. Nat. Med. 2007, 13, 552–559. [Google Scholar] [CrossRef]
- Anwar, M.A.; Shah, M.; Kim, J.; Choi, S. Recent clinical trends in Toll-like receptor targeting therapeutics. Med. Res. Rev. 2019, 39, 1053–1090. [Google Scholar] [CrossRef] [Green Version]
- Rodell, C.B.; Arlauckas, S.P.; Cuccarese, M.F.; Garris, C.S.; Li, R.; Ahmed, M.S.; Weissleder, R. TLR7/8-agonist-loaded nanoparticles promote the polarization of tumour-associated macrophages to enhance cancer immunotherapy. Nat. Biomed. Eng. 2018, 2, 578–588. [Google Scholar] [CrossRef]
- Chen, P.M.; Pan, W.Y.; Wu, C.Y.; Yeh, C.Y.; Korupalli, C.; Luo, P.K.; Chou, C.J.; Chia, W.T.; Sung, H.W. Modulation of tumor microenvironment using a TLR-7/8 agonist-loaded nanoparticle system that exerts low-temperature hyperthermia and immunotherapy for in situ cancer vaccination. Biomaterials 2020, 230, 119629. [Google Scholar] [CrossRef]
- Kim, H.; Khanna, V.; Kucaba, T.A.; Zhang, W.; Ferguson, D.M.; Griffith, T.S.; Panyam, J. Combination of Sunitinib and PD-L1 Blockade Enhances Anticancer Efficacy of TLR7/8 Agonist-Based Nanovaccine. Mol. Pharm. 2019, 16, 1200–1210. [Google Scholar] [CrossRef]
- Zalba, S.; Belsue, V.; Topp, B.; de Alwis, D.; Alvarez, M.; Troconiz, I.F.; Berraondo, P.; Garrido, M.J. Modulation of intratumoural myeloid cells, the hallmark of the anti-tumour efficacy induced by a triple combination: Tumour-associated peptide, TLR-3 ligand and alpha-PD-1. Br. J. Cancer 2021, 124, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Luo, T.; Culbert, A.; Kaufmann, M.; Jiang, X.; Lin, W. Nanoscale Metal-Organic Framework Co-delivers TLR-7 Agonists and Anti-CD47 Antibodies to Modulate Macrophages and Orchestrate Cancer Immunotherapy. J. Am. Chem. Soc. 2020, 142, 12579–12584. [Google Scholar] [CrossRef] [PubMed]
- Nuhn, L.; De Koker, S.; Van Lint, S.; Zhong, Z.; Catani, J.P.; Combes, F.; Deswarte, K.; Li, Y.; Lambrecht, B.N.; Lienenklaus, S.; et al. Nanoparticle-Conjugate TLR7/8 Agonist Localized Immunotherapy Provokes Safe Antitumoral Responses. Adv. Mater. 2018, 30, e1803397. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, P.; Lepland, A.; Scodeller, P.; Fontana, F.; Torrieri, G.; Tiboni, M.; Shahbazi, M.A.; Casettari, L.; Kostiainen, M.A.; Hirvonen, J.; et al. Peptide-guided resiquimod-loaded lignin nanoparticles convert tumor-associated macrophages from M2 to M1 phenotype for enhanced chemotherapy. Acta Biomater. 2021, 133, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Qin, J.; Zhou, C.; Wan, G.; Liu, Y.; Zhang, M.; Yang, X.; Zhang, N.; Wang, Y. Multifunctional nanoparticles based on a polymeric copper chelator for combination treatment of metastatic breast cancer. Biomaterials 2019, 195, 86–99. [Google Scholar] [CrossRef]
- Braunstein, M.J.; Kucharczyk, J.; Adams, S. Targeting Toll-Like Receptors for Cancer Therapy. Target Oncol. 2018, 13, 583–598. [Google Scholar] [CrossRef]
- Federico, S.; Pozzetti, L.; Papa, A.; Carullo, G.; Gemma, S.; Butini, S.; Campiani, G.; Relitti, N. Modulation of the Innate Immune Response by Targeting Toll-like Receptors: A Perspective on Their Agonists and Antagonists. J. Med. Chem. 2020, 63, 13466–13513. [Google Scholar] [CrossRef]


| Tumors | TLRs | Type | Mechanisms | Refs. |
|---|---|---|---|---|
| Glioblastoma | TLR4 | pro- | TLR4 represses RBBP5 to suppress cancer stem cells (CSCs) properties, but CSCs evade innate immunosuppression by reducing TLR4 expression and downregulating its activity | [89,90] |
| TGCT | TLR2 | pro- | A candidate prognostic factor | [91] |
| Breast carcinoma | TLR3 | pro- | Hypoxia-inducible factor 1α (HIF1α)-dependent angiogenesis | [92] |
| EAC | TLR4 | pro- | TLR4-MyD88-TRAF6-NF-κB | [93,94] |
| ESCC | TLR4 | pro- | Production of immunosuppressive cytokines TNF-α and TGF-β | [95] |
| HCC | TLR4, TLR7, TLR9 | pro- | TLR4-AKT-SOX2 signaling pathway enhances cancer stem cell competence; dysregulated AKT/mTOR/4EBP1 signaling | [96,97,98] |
| PDA | TLR4 | pro- | HMGB-1 and S100A8/S100A9 stimulate TLR4 signaling | [99,100] |
| NSCLC | TLR4, TLR3 | pro- | A candidate prognostic factor | [101,102] |
| CRC | TLR4 | pro- | Cathepsin K (CTSK) mediates TLR4-stimulated polarization of M2 macrophages; TLR4-miR-155 positive feedback loop | [103,104] |
| DLBCL | TLR9 | pro- | Formation of neutrophil extracellular traps (NETs) | [105] |
| Melanoma | TLR2, TLR4 | pro- | Overexpression of MMP2 tends toward a potentially tumorigenic phenotype | [106] |
| Melanoma | TLR5 | anti- | Activation of TLR5 signaling by Salmonella flagella | [107] |
| TNBC | TLR4 | anti- | Infiltration and activation of different immune cell types | [108] |
| PC | TLR3 | anti- | Inhibition of proliferation by mitogen-activated protein kinase and induction of apoptosis by cysteine aspartase | [109,110] |
| SCCHN | TLR8 | anti- | Synergistic activation of NK cell activity by motolimod and NLRP3 | [111] |
| Brain tumor | TLR2 | anti- | The glial cell TLR2-MHC I axis contributes to the proliferation and activation of CD8+ T cells | [86] |
| Reagents | Phase(s) | Subjects | Results | Refs. |
|---|---|---|---|---|
| GSK1795091 (TLR4 agonist) | I/II | Health Volunteers | Acceptable safety profile with dose-dependent cytokine and immune cell changes | [150] |
| NY-ESO-1 vaccine + anti-CTLA4 antibody Ipilimumab (IPI) | I | Patients with unresectable or metastatic melanoma | Enhanced immune infiltration | [151] |
| CHP-NY-ESO-1 vaccine combined with poly-ICLC | I | Advanced or recurrent esophageal cancer patients | Acquisition of antibody response | [152] |
| Neoadjuvant Cetuximab and motolimod | Ib | Head and Neck Cancer | Reversal of myeloid-derived suppressor cells (MDSC) inhibition of T cell proliferation | [153] |
| Vaccination With Poly-ICLC and Peptide-pulsed Dendritic Cells | N/A | Patients with pancreatic cancer | Good tolerance and effective production of antigen-specific T cells | [154] |
| A transdermal multipeptide melanoma vaccine administered with or without a TLR7 agonist | N/A | Melanoma | TLR7 agonist imiquimod as a vaccine adjuvant | [155] |
| Topical Imiquimod Plus Nab-paclitaxel | II | Breast Cancer Cutaneous Metastases | Effective in inducing disease regression in refractory breast cancer chest wall metastases, but response is transient | [156] |
| A long peptide vaccine (LPV7) plus toll-like receptor (TLR) agonists | I/II | Resected high-risk melanoma | Enhanced T-cell response | [157] |
| Combined Vaccination with NY-ESO-1 Protein, Poly-ICLC, and Montanide | I/II | Patients with High-Risk MelanomaPatients with High-Risk Melanoma | The combination is safe, well tolerated, and induces integrated antibody and CD4+ T cell responses | [158] |
| GNKG168(TLR9 agonist) | I | children with minimal residual disease positive acute leukemia | Induction of an immediate NK response, followed by adaptive T and B cell responses | [159] |
| TLR8 Agonist, Motolimod (VTX-2337), Combined with Cetuximab | Ib | Patients with Recurrent or Metastatic SCCHN | Motolimod co-administered with cetuximab has safety profile | [160] |
| IMO-2125 (TLR9 agonist) | I/II | Pancreatic cancer | Triggers immune system response and kills local and distant tumors | [161] |
| Tilsotolimod (TLR9 agonist) | II/III | Refractory solid tumors and melanoma | Immunomodulation of TME and therapeutic effects on distant tumor lesions | [162] |
| G100 (TLR4 agonist) | N/A | Patients with Merkel Cell Carcinoma | Induces Antitumor Immune Responses and Tumor Regression | [163] |
| SD-101 (TLR9 agonist) and Local Low-Dose Radiation | I/II | Untreated Indolent Lymphoma | Combination therapy is safe and well tolerated | [164] |
| Motolimod with pegylated liposomal doxorubicin | II | Recurrent or persistent ovarian cancer | Combination therapy was well tolerated, but did not significantly improve clinical outcomes | [165] |
| Peptide vaccine with glucopyranosyl lipid A-stable oil-in-water emulsion | IIA | Patients with resected melanoma | Is well tolerated and leads to an increase in specific T cells | [166] |
| PF-3512676 (TLR9 agonist) | I/II | Lymphoma | Well-tolerated, induces systemic anti-tumor immunity | [167] |
| TLR9 agonist combined with radiation | I/II | Mycosis fungoides | Systemic tumor regression | [168] |
| Combination | Treatment Effect | Refs. |
|---|---|---|
| CDNPs | Promote the polarization of tumor-associated macrophages | [199] |
| R848@NPs+NIR | Effectively internalized by dendritic cells and generate robust anti-tumor memory immunity | [200] |
| Sunitinib/PD-L1 Blockade/TLR7/8 Agonist-Based Nanovaccine | Reduced MDSC and Tregs, and reduced CD8 T cell depletion | [201] |
| HPV E7 long peptide/PIC/anti-PD-1 mAb | Treatment of primary drug-resistant tumors | [202] |
| nMOFs/TLR7 agonist/Anti-CD47 Antibodies | nMOFs are radiosensitizers; the TLR7 agonist imiquimod (IMD) repolarizes immunosuppressive M2 macrophages to immunostimulatory M1 macrophages; and anti-CD47 antibodies block CD47 tumor cell surface markers to promote phagocytosis | [203] |
| Nanoparticle-Conjugate TLR7/8 Agonist/anti-PD-1 mAb/Flt3L | Leads to efficient activation of DCs in sentinel lymph nodes and promotes proliferation of tumor antigen-specific CD8 T cells | [204] |
| R848@LNPs | Promote the conversion of tumor-associated macrophages from the M2 phenotype to the M1 phenotype | [205] |
| RPTDH/R848 | Anti-angiogenesis and immune activation | [206] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, R.; Ma, J. Immunotherapeutic Implications of Toll-like Receptors Activation in Tumor Microenvironment. Pharmaceutics 2022, 14, 2285. https://doi.org/10.3390/pharmaceutics14112285
Zheng R, Ma J. Immunotherapeutic Implications of Toll-like Receptors Activation in Tumor Microenvironment. Pharmaceutics. 2022; 14(11):2285. https://doi.org/10.3390/pharmaceutics14112285
Chicago/Turabian StyleZheng, Run, and Jian Ma. 2022. "Immunotherapeutic Implications of Toll-like Receptors Activation in Tumor Microenvironment" Pharmaceutics 14, no. 11: 2285. https://doi.org/10.3390/pharmaceutics14112285