

Secondary Metabolite Profile and Pharmacological Opportunities of Lettuce Plants following Selenium and Sulfur Enhancement

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lettuce Plant Growth and Treatments
2.2. Extraction
2.3. UPLC-MS Analyses
2.4. Nitric Oxide (NO) Production Inhibition Assay in RAW 264.7 Murine Macrophages
2.4.1. Cell Preparation
2.4.2. Nitrite Measurement
2.4.3. Determination of Cell Viability
2.5. Minimum Inhibitory Concentration (MIC)
2.6. Cytotoxicity against Vero Cells
2.7. Statistical Analysis
3. Results
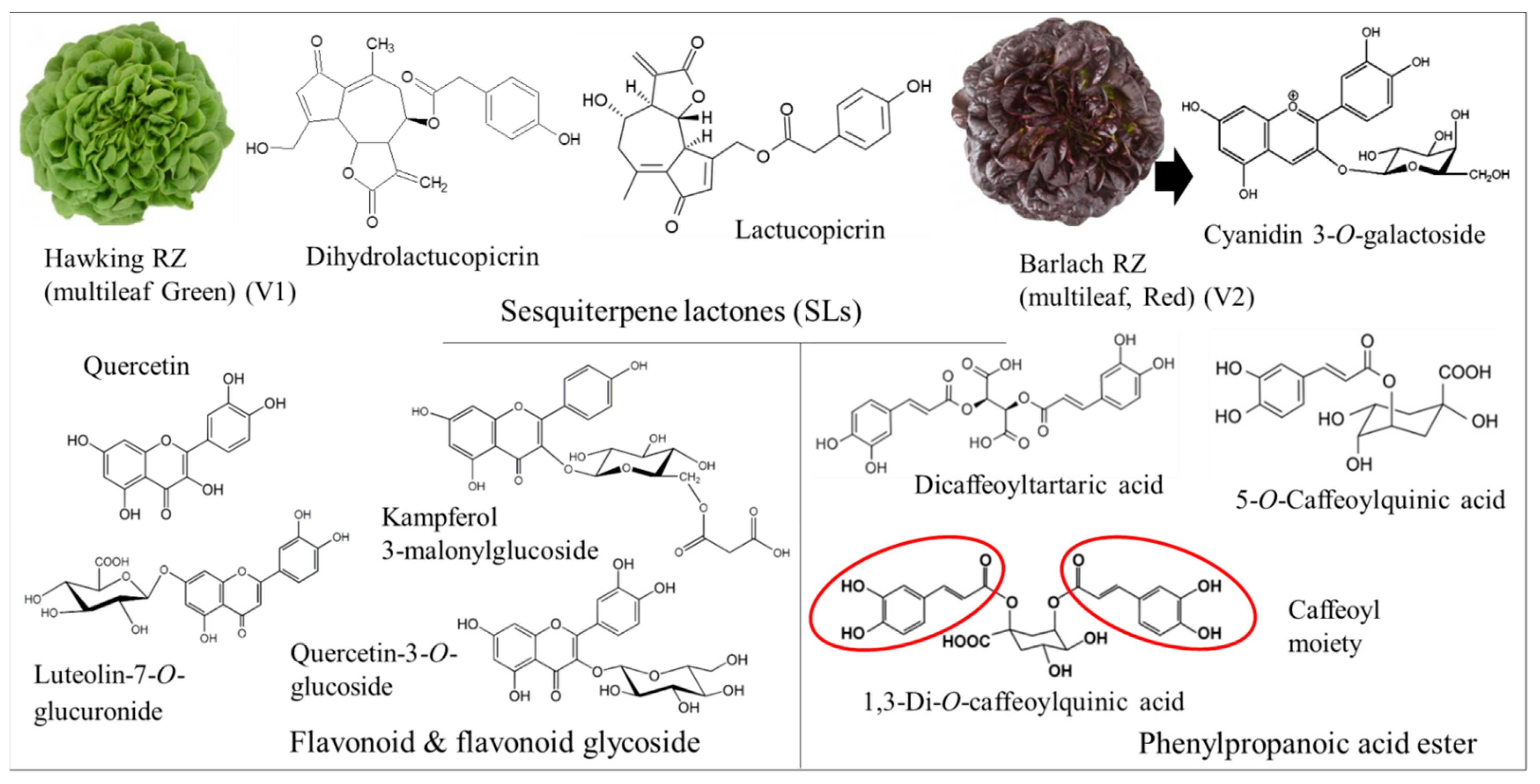
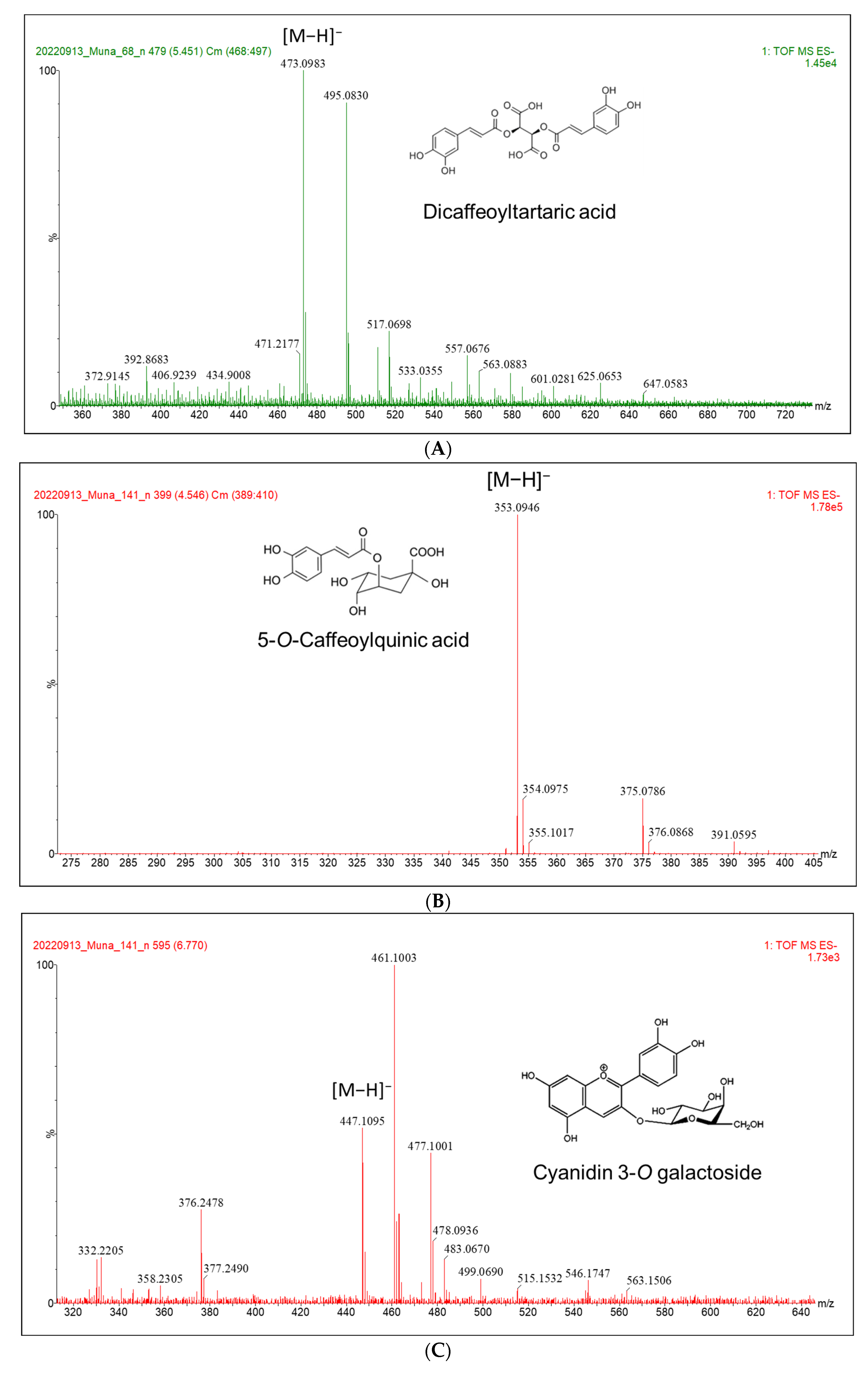
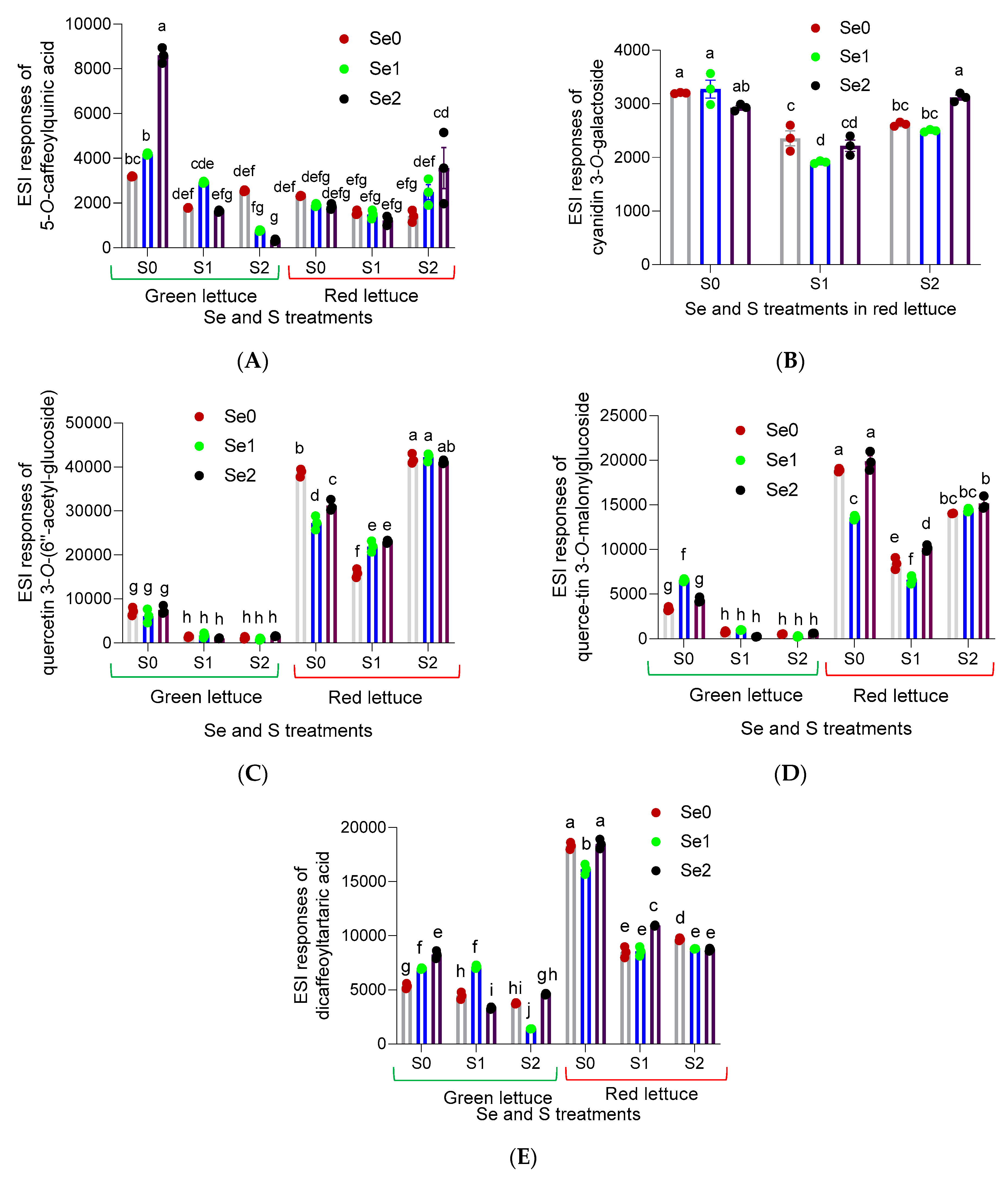
3.1. Secondary Metabolite Profile Following Se and S Enrichment
3.2. The Effect of Se and S Interaction on the NO Inhibitory Activity
3.3. The Effect of Se and S Interaction on the Antibacterial Activity
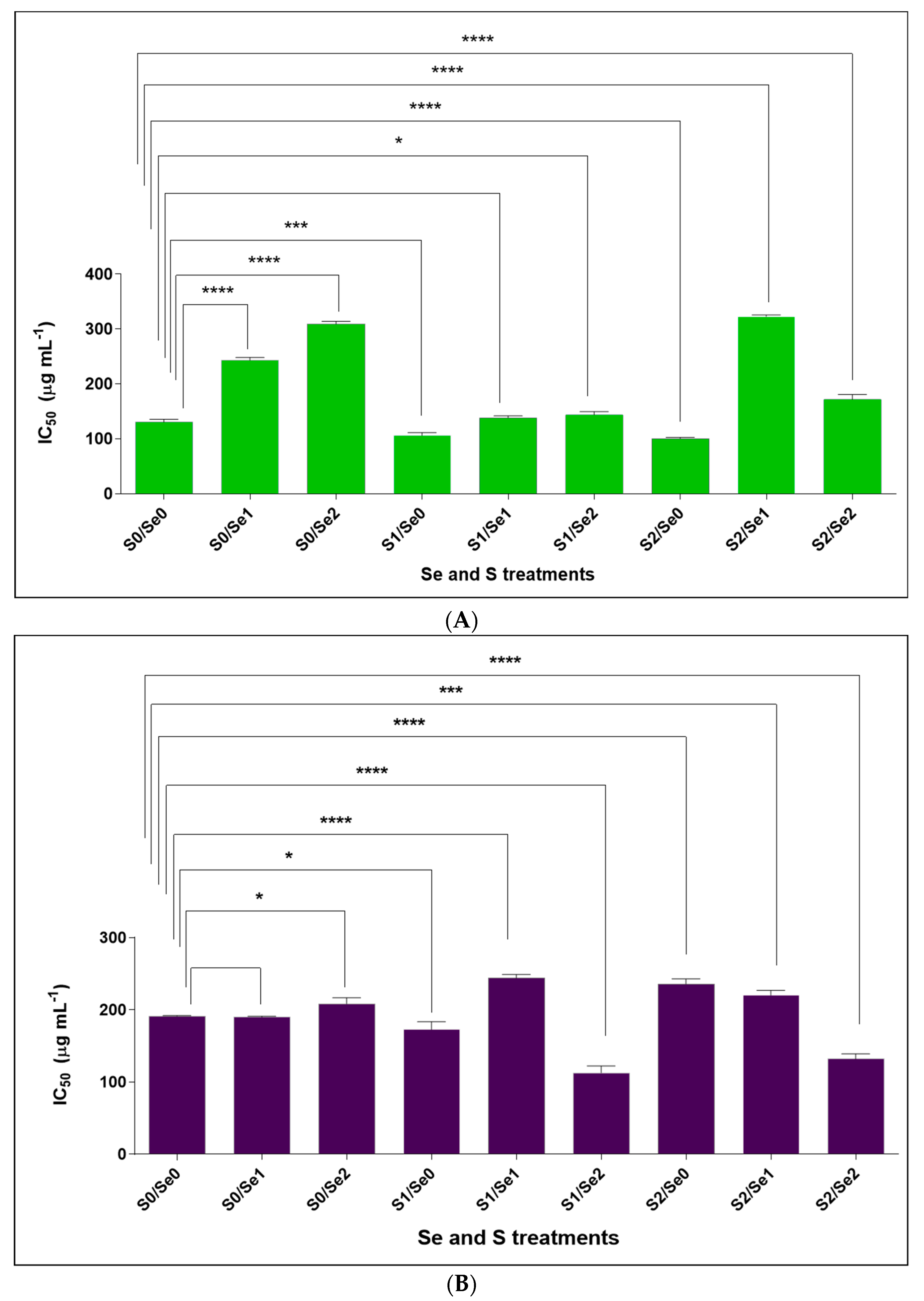
3.4. Cytotoxicity (LC50) Assay against Monkey Kidney (Vero) Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duntas, L.H. Selenium and inflammation: Underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009, 41, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barchielli, G.; Capperucci, A.; Tanini, D. The role of selenium in pathologies: An updated review. Antioxidants 2022, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Castro, W.M. Selenium in critically ill patients with systemic inflammatory response. Nutr. Hosp. 2007, 22, 295–306. [Google Scholar]
- Cremonini, E.; Zonaro, E.; Donini, M.; Lampis, S.; Boaretti, M.; Dusi, S.; Melotti, P.; Lleo, M.M.; Vallini, G. Biogenic selenium nanoparticles: Characterization, antimicrobial activity and effects on human dendritic cells and fibroblasts. Microb. Biotechnol. 2016, 9, 758–771. [Google Scholar] [CrossRef] [Green Version]
- Filipović, N.; Ušjak, D.; Milenković, M.T.; Zheng, K.; Liverani, L.; Boccaccini, A.R.; Stevanović, M.M. Comparative study of the antimicrobial activity of selenium nanoparticles with different surface chemistry and structure. Front. Bioeng. Biotechnol. 2021, 8, 624621. [Google Scholar] [CrossRef]
- Lee, D.Y.; Li, H.; Lim, H.J.; Lee, H.J.; Jeon, R.; Ryu, J.-H. Anti-inflammatory activity of sulfur-containing compounds from garlic. J. Med. Food. 2012, 15, 992–999. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Cao, L.; Zhang, W.; Lu, R.; Bian, J.S.; Nie, X. Therapeutic potential of sulfur-containing natural products in inflammatory diseases. Pharmacol. Ther. 2020, 216, 107687. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Li, F.; Wenzel-Storjohann, A.; Sulieman, S.; Tasdemir, D.; Mühling, K.H. Comparative metabolite profile, biological activity and overall quality of three lettuce (Lactuca sativa L., Asteraceae) cultivars in response to sulfur nutrition. Pharmaceutics 2021, 13, 713. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Regulation of adaptive immunity by the innate immune system. Science 2010, 327, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Fouladseresht, H.; Ghamar Talepoor, A.; Eskandari, N.; Norouzian, M.; Ghezelbash, B.; Beyranvand, M.R.; Nejadghaderi, S.A.; Carson-Chahhoud, K.; Kolahi, A.-A.; Safiri, S. Potential immune indicators for predicting the prognosis of COVID-19 and trauma: Similarities and disparities. Front. Immunol. 2022, 12, 785946. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, M.K.; Lee, H.J.; Kim, J.S.; Ryu, J.-H. A curcuminoid and two sesquiterpenoids from Curcuma zedoaria as inhibitors of nitric oxide synthesis in activated macrophages. Arch. Pharm. Res. 2004, 27, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Ivanescu, B.; Miron, A.; Corciova, A. Sesquiterpene lactones from Artemisia genus: Biological activities and methods of analysis. J. Anal. Methods Chem. 2015, 12, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespo, I.; García-Mediavilla, M.V.; Gutiérrez, B.; Sánchez-Campos, S.; Tuñón, M.J.; González-Gallego, J. A comparison of the effects of kaempferol and quercetin on cytokine-induced pro-inflammatory status of cultured human endothelial cells. Br. J. Nutr. 2008, 100, 968–976. [Google Scholar] [CrossRef] [Green Version]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A.; et al. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Badary, O.A.; Yassin, N.A.Z.; El-Shenawy, S.M.A.; EL-Moneem, M.A.; AL-Shafeiy, H.M. Study of the effect of Allium porrum on hypertension induced in rats. Rev. Latinoam. Quím. 2013, 41, 3. [Google Scholar]
- Dabeek, W.M.; Marra, M.V. Dietary quercetin and kaempferol: Bioavailability and potential cardiovascular-related bioactivity in human. Nutrients 2019, 11, 2288. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.A.; Wick, J.E.; Famuyide, I.M.; McGaw, L.J.; Mühling, K.H. Selenium enrichment of green and red lettuce and the induction of radical scavenging potential. Horticulturae 2021, 7, 488. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Lentz, C.; Mühling, K.H. Selenium and sulfur crosstalk is associated with changes in primary metabolism in lettuce plants grown under Se and S enrichment. Plants 2022, 11, 927. [Google Scholar] [CrossRef] [PubMed]
- Famuyide, I.M.; Aro, A.O.; Fasina, F.O.; Eloff, J.N.; McGaw, L.J. Antibacterial and antibiofilm activity of acetone leaf extracts of nine under-investigated south African Eugenia and Syzygium (Myrtaceae) species and their selectivity indices. BMC Complement. Altern. Med. 2019, 19, 141. [Google Scholar] [CrossRef] [PubMed]
- Mehrbod, P.; Abdalla, M.A.; Fotouhi, F.; Heidarzadeh, M.; Aro, A.O.; Eloff, J.N.; McGaw, L.J.; Fasina, F.O. Immunomodulatory properties of quercetin-3-O-α-L-rhamnopyranoside from Rapanea melanophloeos against influenza a virus. BMC Complement. Altern. Med. 2018, 18, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasonga, T.K.; Coetzee, M.A.A.; Van Zijl, C.; Benteke Momba, M.N. Data on UPLC/MS method validation for the biodegradation of pharmaceuticals and intermediates by a fungal consortium and on T47DK-Bluc reporter gene assay to assess the reduction of their estrogenic activity. Data Brief. 2019, 25, 104336. [Google Scholar] [CrossRef]
- Mu, M.M.; Chakravortty, D.; Sugiyama, T.; Koide, N.; Takahashi, K.; Mori, I.; Yoshida, T.; Yokochi, T. The inhibitory action of quercetin on lipopolysaccharide-induced nitric oxide production in RAW 264. 7 macrophage cells. J. Endotoxin Res. 2001, 7, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Ncube, S.F.; McGaw, L.J.; Njoya, E.M.; Ndagurwa, H.G.T.; Mundy, P.J.; Sibanda, S. In vitro antioxidant activity of crude extracts of Harpagophytum zeyheri and their anti-inflammatory and cytotoxicity activity compared with diclofenac. BMC Complement. Med. Ther. 2021, 21, 238. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotox-icity assays. J. Immunol. Methods 1983, 16, 55–63. [Google Scholar] [CrossRef]
- Eloff, J.N. A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta Med. 1998, 64, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Gado, D.A.; Abdalla, M.A.; Ahmed, A.S.; Madikizela, B.; Nkadimeng, S.M.; Ehlers, M.M.; McGaw, L.J. In vitro antibacterial activity of Loxostylis alata extracts and isolated compounds against Salmonella species. BMC Complement. Med. Ther. 2021, 21, 121. [Google Scholar] [CrossRef]
- McGaw, L.J.; Steenkamp, V.; Eloff, J.N. Evaluation of Athrixia bush tea for cytotoxicity, antioxidant activity, caffeine content andpresence of pyrrolizidine alkaloids. J. Ethnopharmcol. 2007, 110, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.A.; Lawande, K.E.; Patil, V.B.; Mani, S. Relationship between selenium and sulfur assimilation and resultant interaction on quality parameters in onion. Commun. Soil Sci. Plant Anal. 2012, 43, 1625–1631. [Google Scholar] [CrossRef]
- Wessjohann, L.A.; Schneider, A.; Abbas, M.; Brandt, W. Selenium in chemistry and biochemistry in comparison to sulfur. Biol. Chem. 2007, 388, 997–1006. [Google Scholar] [CrossRef]
- Adebayo, S.A.; Dzoyem, J.P.; Shai, L.J.; Eloff, J.N. The anti-inflammatory and antioxidant activity of 25 plant species used traditionally to treat pain in southern African. BMC Complement. Altern. Med. 2015, 15, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzoyem, J.P.; Nkuete, A.H.L.; Ngameni, B.; Eloff, J.N. Anti-inflammatory and anticholinesterase activity of six flavonoids isolated from Polygonum and Dorstenia species. Arch. Pharm. Res. 2017, 40, 1129–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niranjan, R.; Zafar, S.; Lochab, B.; Priyadarshini, R. Synthesis and characterization of sulfur and sulfur-selenium nanoparticles loaded on reduced graphene oxide and their antibacterial activity against gram-positive pathogens. Nanomaterials 2022, 12, 191. [Google Scholar] [CrossRef]
- Olawuwo, O.S.; Famuyide, I.M.; McGaw, L.J. Antibacterial and antibiofilm activity of selected medicinal plant leaf extracts against pathogens implicated in poultry diseases. Front. Vet. Sci. 2022, 9, 820304. [Google Scholar] [CrossRef]
- Shai, L.J.; McGaw, L.J.; Aderogba, M.A.; Mdee, L.K.; Eloff, J.N. Four pentacyclic triterpenoids with antifungal and antibacterial activity from Curtisia dentata (Burm.f) C.A. Sm. leaves. J. Ethnopharmacol. 2008, 119, 238–244. [Google Scholar] [CrossRef]
- Fernández Freire, P.; Peropadre, A.; Pérez Martín, J.M.; Herrero, O.; Hazen, M.J. An integrated cellular model to evaluate cytotoxic effects in mammalian cell lines. Toxicol. In Vitro 2009, 23, 1553–1558. [Google Scholar] [CrossRef]
- McGaw, L.J.; Elgorashi, E.E.; Eloff, J.N. Cytotoxicity of African medicinal plants against normal animal and human cells. In Toxicological Survey of African Medicinal Plants; Kuete, V., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 181–233. [Google Scholar]
- Hu, W.; Zhao, C.; Hu, H.; Yin, S. Food Sources of Selenium and Its Relationship with Chronic Diseases. Nutrients 2021, 13, 1739. [Google Scholar] [CrossRef]
- Abdalla, M.A.; Sulieman, S.; Mühling, K.H. Regulation of selenium/sulfur interactions to enhance chemopreventive effects: Lessons to learn from Brassicaceae. Molecules 2020, 25, 5846. [Google Scholar] [CrossRef]
- D’Amato, R.; Regni, L.; Falcinelli, B.; Mattioli, S.; Benincasa, P.; Dal Bosco, A.; Pacheco, P.; Proietti, P.; Troni, E.; Santi, C.; et al. Current knowledge on selenium biofortification to improve the nutraceutical profile of food: A comprehensive review. J. Agric. Food Chem. 2020, 68, 4075–4097. [Google Scholar] [CrossRef] [PubMed]
- Galić, L.; Špoljarević, M.; Jakovac, E.; Ravnjak, B.; Teklić, T.; Lisjak, M.; Perić, K.; Nemet, F.; Lončarić, Z. Selenium biofortification of soybean seeds influences physiological responses of seedlings to osmotic stress. Plants 2021, 10, 1498. [Google Scholar] [CrossRef] [PubMed]
- Benincasa, P.; D’Amato, R.; Falcinelli, B.; Troni, E.; Fontanella, M.C.; Frusciante, S.; Guiducci, M.; Beone, G.M.; Businelli, D.; Diretto, G. Grain endogenous selenium and moderate salt stress work as synergic elicitors in the enrichment of bioactive compounds in maize sprouts. Agronomy 2020, 10, 735. [Google Scholar] [CrossRef]
- Nikiforova, V.J.; Kopka, J.; Tolstikov, V.; Fiehn, O.; Hopkins, L.; Hawkesford, M.L.; Hesse, H.; Hoefgen, R. Systems re-balancing of metabolism in response to sulfur deprivation, as revealed by metabolome analysis of Arabidopsis plants. Plant Physiol. 2005, 138, 304–318. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.-S.; Kim, H.-C.; Yun, S.-M.; Kim, H.-J.; Kim, C.-S.; Lee, J.-J. Metabolite analysis of lettuce in response to sulfur nutrition. Horticulturae 2022, 8, 734. [Google Scholar] [CrossRef]
- Singh, J.; Lee, Y.; Kellum, J.A. A new perspective on NO pathway in sepsis and ADMA lowering as a potential therapeutic approach. Crit. Care 2022, 26, 246. [Google Scholar] [CrossRef]
- Sp, N.; Kang, D.Y.; Kim, H.D.; Rugamba, A.; Jo, E.S.; Park, J.-C.; Bae, S.W.; Lee, J.-M.; Jang, K.-J. Natural sulfurs inhibit LPS-induced inflammatory responses through NF-κB signaling in CCD-986Sk skin fibroblasts. Life 2021, 11, 427. [Google Scholar] [CrossRef]
- Mattmiller, S.A.; Carlson, B.A.; Sordillo, L.M. Regulation of inflammation by selenium and selenoproteins: Impact on eicosanoid biosynthesis. J. Nutr. Sci. 2013, 2, e28. [Google Scholar] [CrossRef] [Green Version]
- Martinez, S.S.; Huang, Y.; Acuna, L.; Laverde, E.; Trujillo, D.; Barbieri, M.A.; Tamargo, J.; Campa, A.; Baum, M.K. Role of selenium in viral infections with a major focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 23, 280. [Google Scholar] [CrossRef]
- Sappey, C.; Legrand-Poelsl, S.; Best-Belpomme, M.; Favier, A.; Rentier, B.; Piette, J. Stimulation of glutathione peroxidase activity decreases HIV type 1 activation after oxidative stress. AIDS Res. Hum. Retrovir. 1994, 10, 1451–1461. [Google Scholar] [CrossRef]
- Medina-Lozano, I.; RamónBertolín, J.; Díaz, A. Nutritional value of commercial and traditional lettuce (Lactuca sativa L.) and wild relatives: Vitamin C and anthocyanin content. Food Chem. 2021, 359, 129864. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Liang, H.; Guo, Y.; Yang, D. Cyanidin 3-O-galactoside: A natural compound with multiple health benefits. Int. J. Mol. Sci. 2021, 22, 2261. [Google Scholar] [CrossRef] [PubMed]
- Cisowska, A.; Wojnicz, D.; Hendrich, A.B. Anthocyanins as antimicrobial agents of natural plant origin. Nat. Prod. Commun. 2011, 6, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.B. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Bhatt, S.K.; Javagal, R.M.; Nanjarajurs, M.S.; Eligar, S.M. In vitro anti-inflammatory property of a Quercetin-3-O-diglucoside-7-O-glucoside characterized from fresh leaves of Trigonella foenum-graecum L. Int. J. Food Prop. 2021, 24, 1438–1452. [Google Scholar] [CrossRef]
- Abdel Motaal, A.; Ezzat, S.M.; Tadros, M.G.; El-Askary, H.I. In vivo anti-inflammatory activity of caffeoylquinic acid derivatives from Solidago virgaurea in rats. Pharm. Biol. 2016, 54, 2864–2870. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Sun, Q.; Gao, R.; Park, Y. AAK-2 and SKN-1 are involved in chicoric-acid-induced lifespan extension in Caenorhabditis elegans. J. Agric. Food Chem. 2019, 67, 9178–9186. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Observed m/z | Mass Error (mDa) | Observed Retention Time (min) | Detector Counts | Adducts |
|---|---|---|---|---|---|
| Luteolin-7,4′-di-O-glucoside | 611.1626 | 1.9 | 6.14 | 552 | [M + H]+, [M + Na]+, [M + K]+, [M − H]− |
| Quercetin | 303.0472 | −2.7 | 8.47 | 389 | [M + H]+ |
| 5-O-Caffeoylquinic acid | 377.0826 | −1.7 | 4.52 | 26,433 | [M + Na]+, [M + H]+, [M − H2O]+, [M − H]− |
| Cyanidin 3-O-galactoside | 447.0939 | 0.6 | 6.78 | 3923 | [M − H]−, [M + H]+ |
| Kaempferol 3-(6′′-malonylglucoside) | 535.11 | 1.8 | 7.71 | 43 | [M + H]+ |
| Luteolin-7-O-glucuronide | 463.0853 | −1.8 | 6.81 | 8080 | [M + H]+, [M + Na]+, [M − H]− |
| Quercetin 3-O-(6′′-acetyl-glucoside) | 507.1166 | 3.2 | 7.65 | 43 | [M + H]+, [M − H]− |
| Quercetin 3-O-malonylglucoside | 551.1044 | 1.2 | 7.2 | 5378 | [M + H]+, [M + Na]+, [M − H]− |
| Quercetin-3-O-glucoside | 487.0842 | −0.5 | 7.02 | 4520 | [M + Na]+, [M + H]+, [M + K]+, [M − H]− |
| Dihydrolactucopicrin | 413.1585 | 0.7 | 4.5 | 26,433 | [M + H]+, [M − H]− |
| Lactucopicrin | 411.1405 | −3.4 | 8.45 | 71 | [M + H]+, [M − H]− |
| Quercetin 3,4′-diglucoside | 625.1426 | 1.5 | 4.81 | 518 | [M-H]-, [M + H]+, [M + Na]+ |
| Caffeic acid hexoside | 341.0885 | 0.7 | 3.7 | 536 | [M − H]− |
| Dicaffeoylquininic acid | 515.1209 | 1.4 | 6.52 | 6430 | [M − H]− |
| Dicaffeoyltartaric acid | 473.0723 | −0.3 | 5.62 | 11,271 | [M − H]− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, M.A.; Famuyide, I.; Wooding, M.; McGaw, L.J.; Mühling, K.H. Secondary Metabolite Profile and Pharmacological Opportunities of Lettuce Plants following Selenium and Sulfur Enhancement. Pharmaceutics 2022, 14, 2267. https://doi.org/10.3390/pharmaceutics14112267
Abdalla MA, Famuyide I, Wooding M, McGaw LJ, Mühling KH. Secondary Metabolite Profile and Pharmacological Opportunities of Lettuce Plants following Selenium and Sulfur Enhancement. Pharmaceutics. 2022; 14(11):2267. https://doi.org/10.3390/pharmaceutics14112267
Chicago/Turabian StyleAbdalla, Muna Ali, Ibukun Famuyide, Madelien Wooding, Lyndy J. McGaw, and Karl H. Mühling. 2022. "Secondary Metabolite Profile and Pharmacological Opportunities of Lettuce Plants following Selenium and Sulfur Enhancement" Pharmaceutics 14, no. 11: 2267. https://doi.org/10.3390/pharmaceutics14112267