Osteoprotegerin Is more than a Possible Serum Marker in Liver Fibrosis: A Study into Its Function in Human and Murine Liver

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Human Liver Tissue
2.3. Liver Fibrosis Model
2.4. Precision-Cut Liver Slices
2.5. Cell Culture
2.6. Generation of Liver Tissue Lysates
2.7. OPG Analysis
2.8. mRNA Analysis
2.9. Single-Cell RNA Sequencing and Analyses
2.10. Immunohistochemistry
2.11. Statistics
3. Results
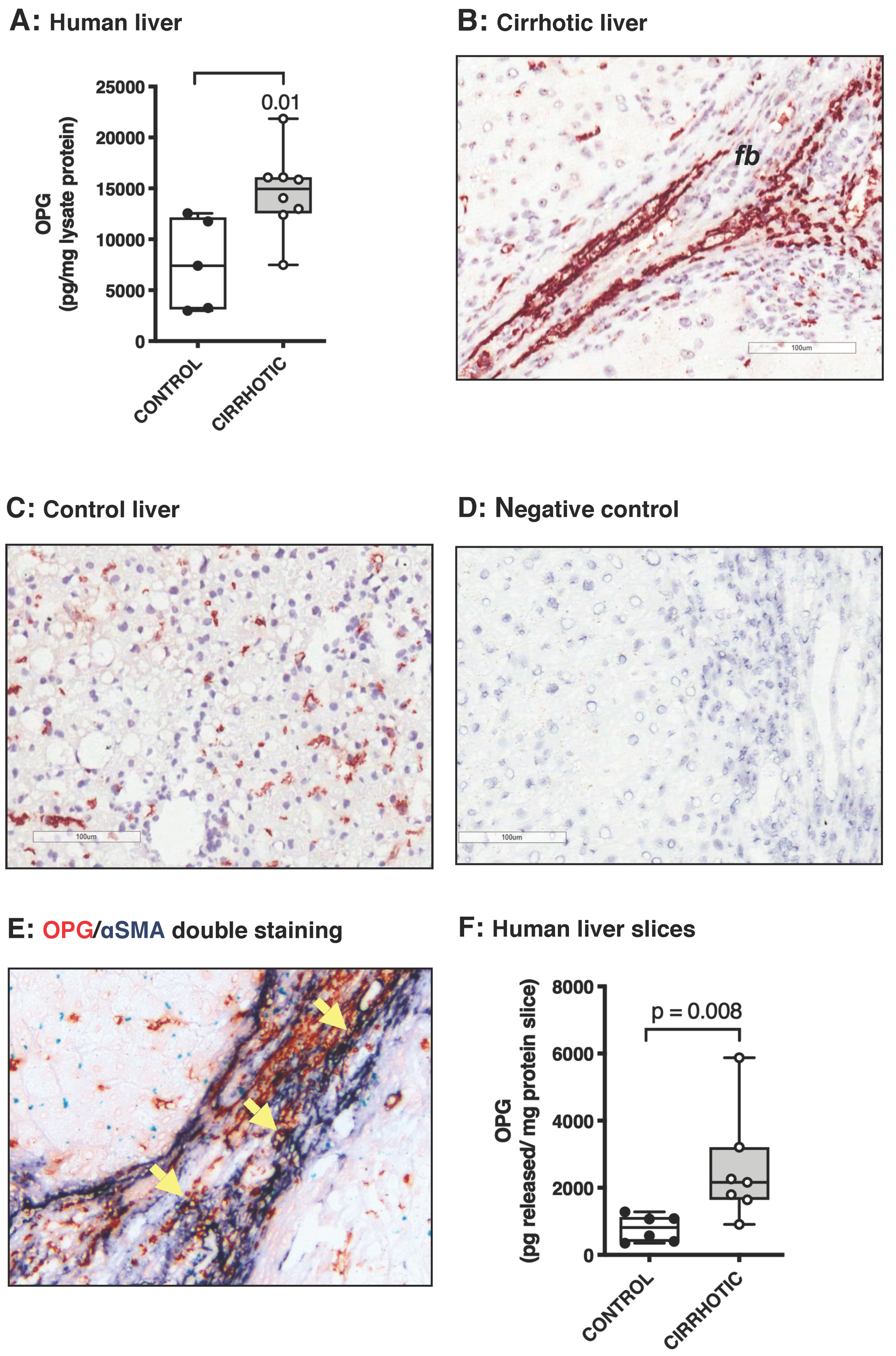
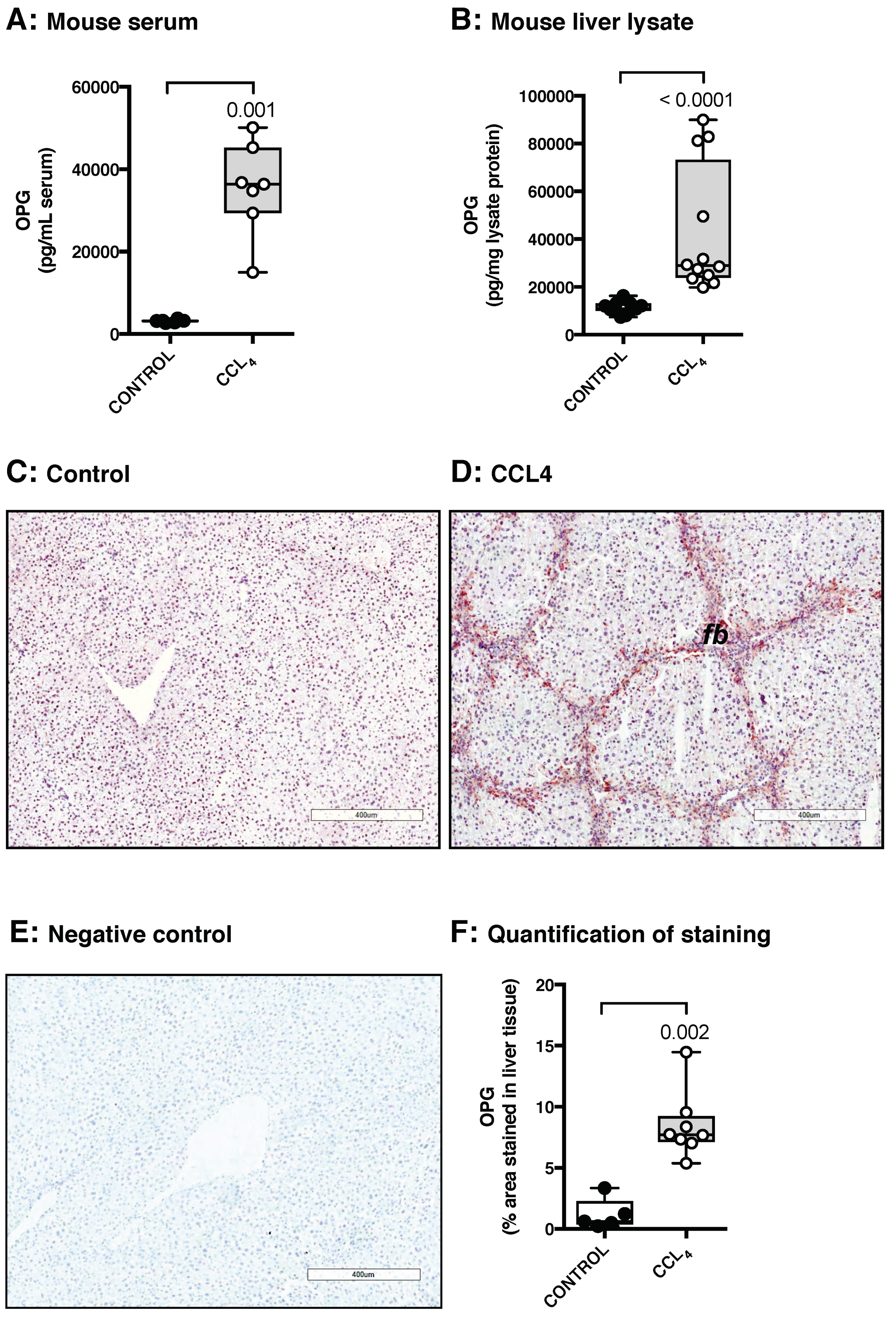
3.1. Osteoprotegerin Expression Is Higher in Human and Murine Fibrotic Livers
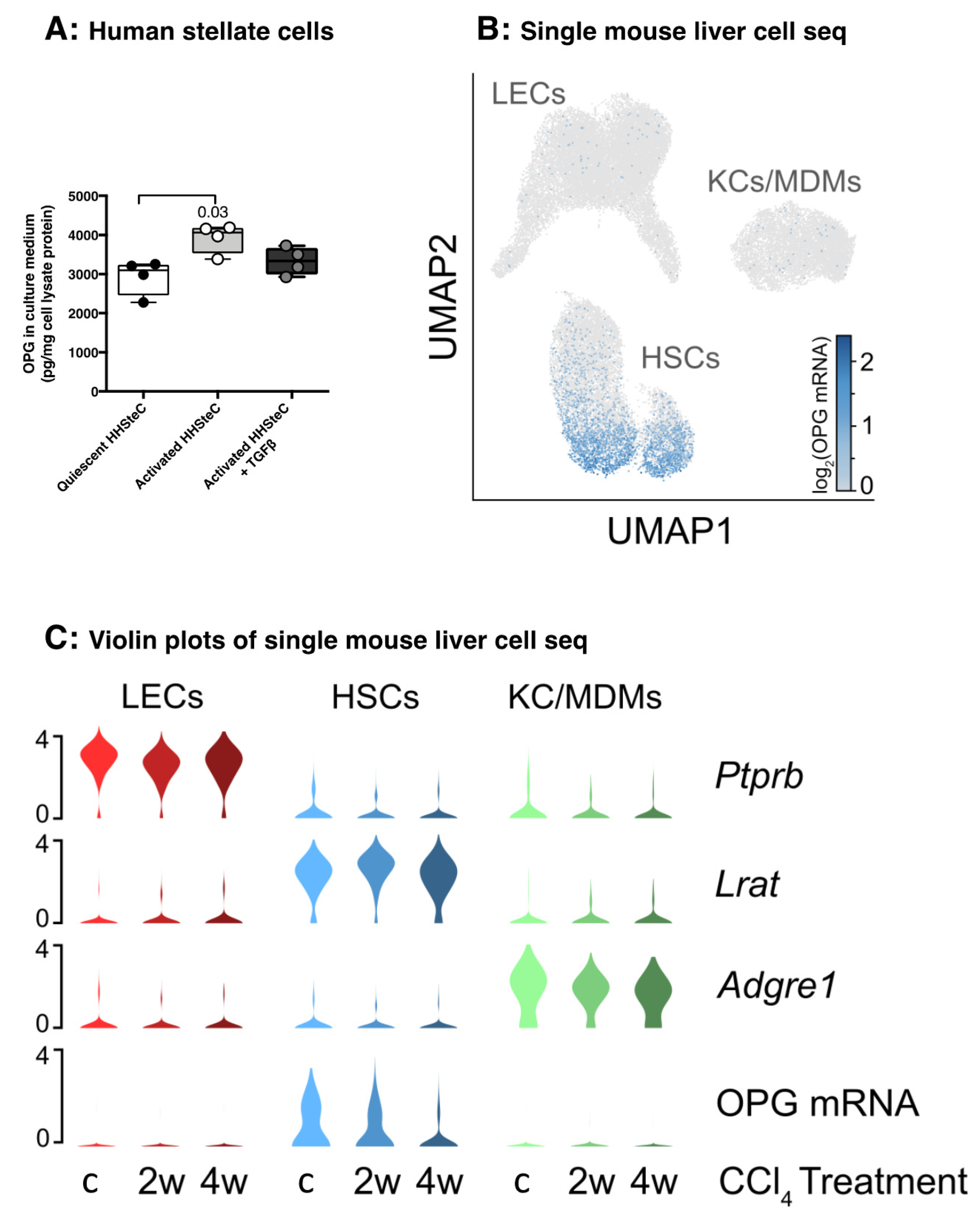
3.2. Hepatic Stellate Cells Produce Copious Amounts of OPG
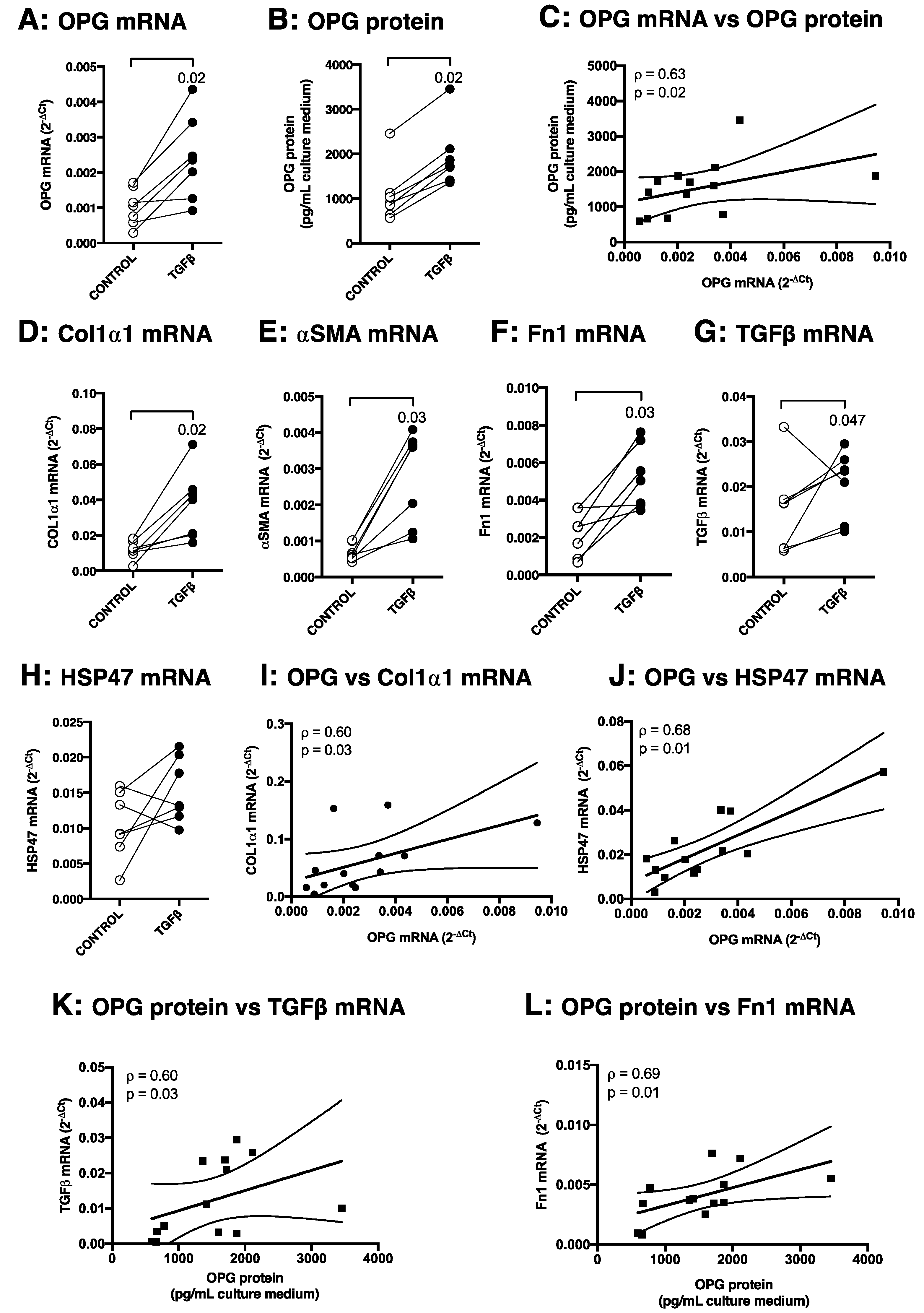
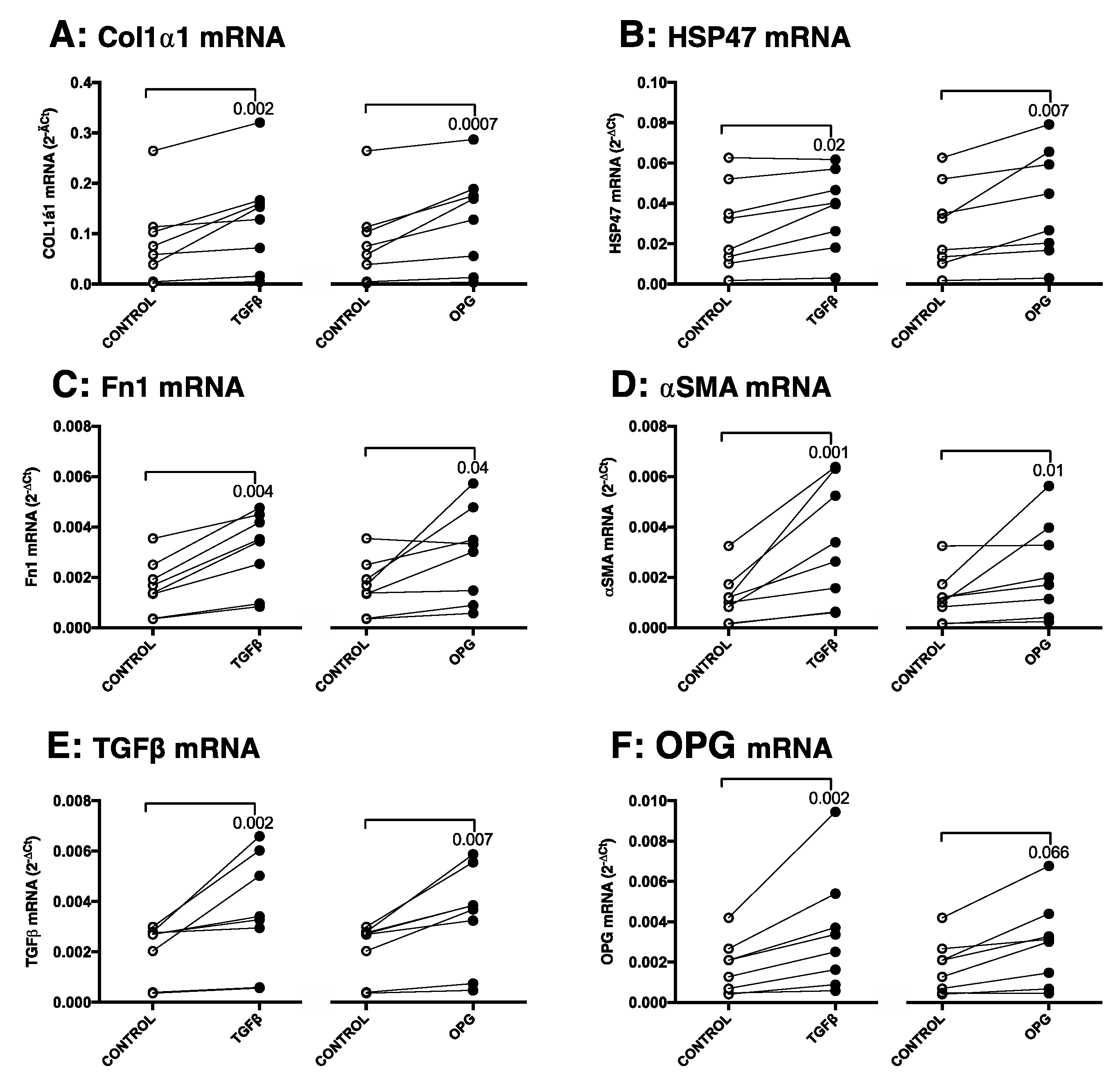
3.3. TGFβ1 Induces OPG mRNA and Protein Production in Murine Precision-Cut Liver Slices, Which Correlates with Other Markers of Fibrosis
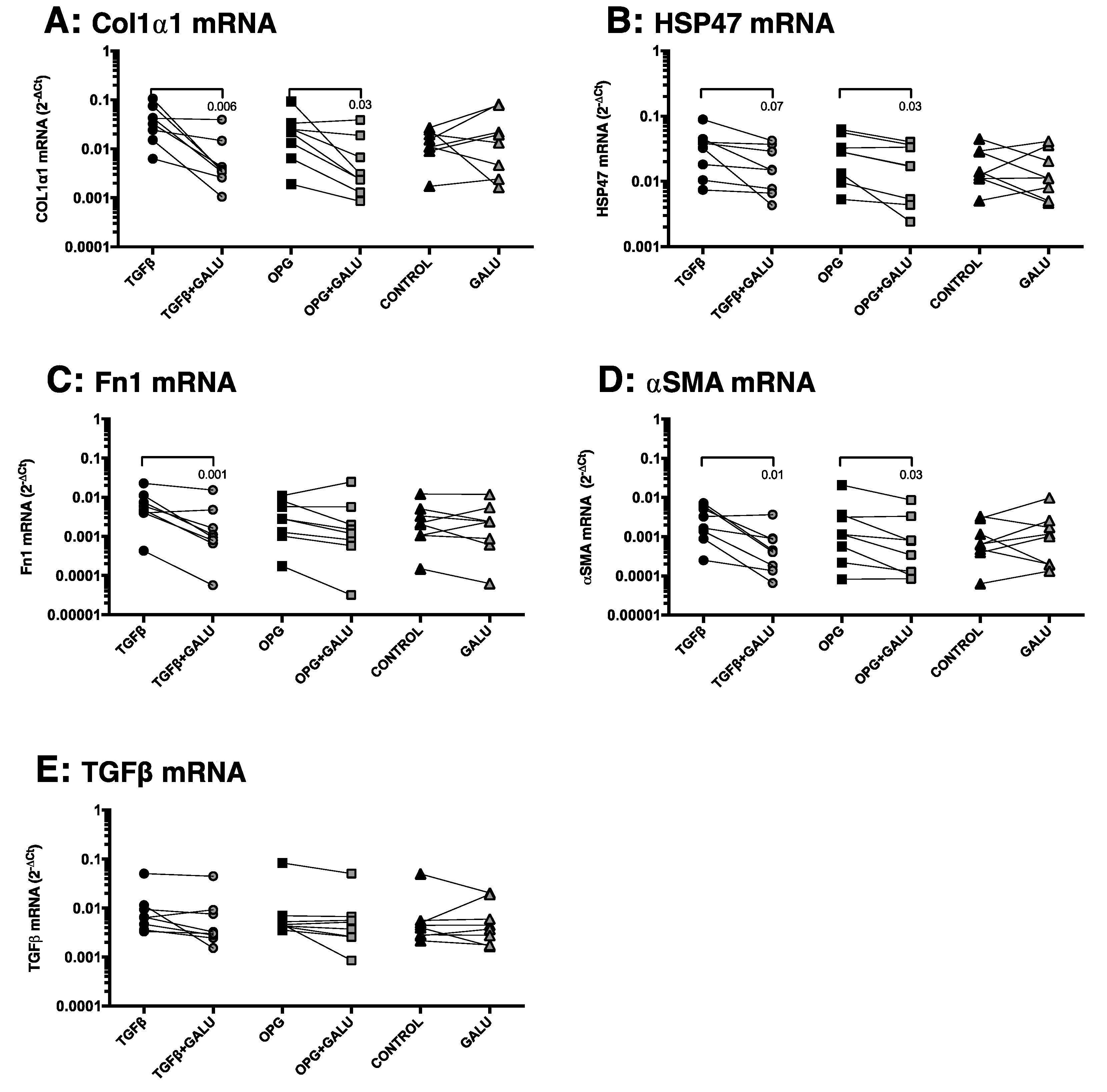
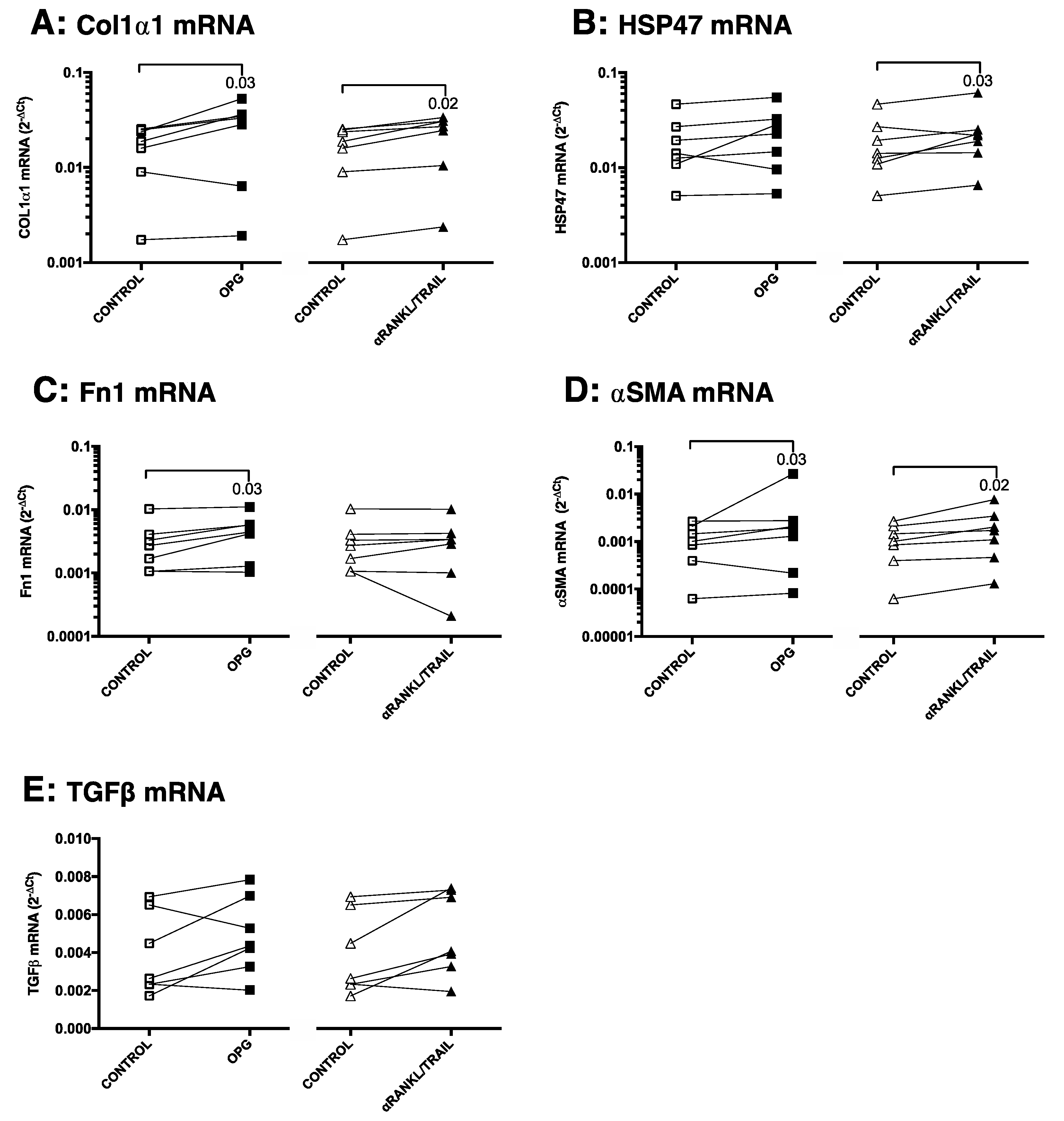
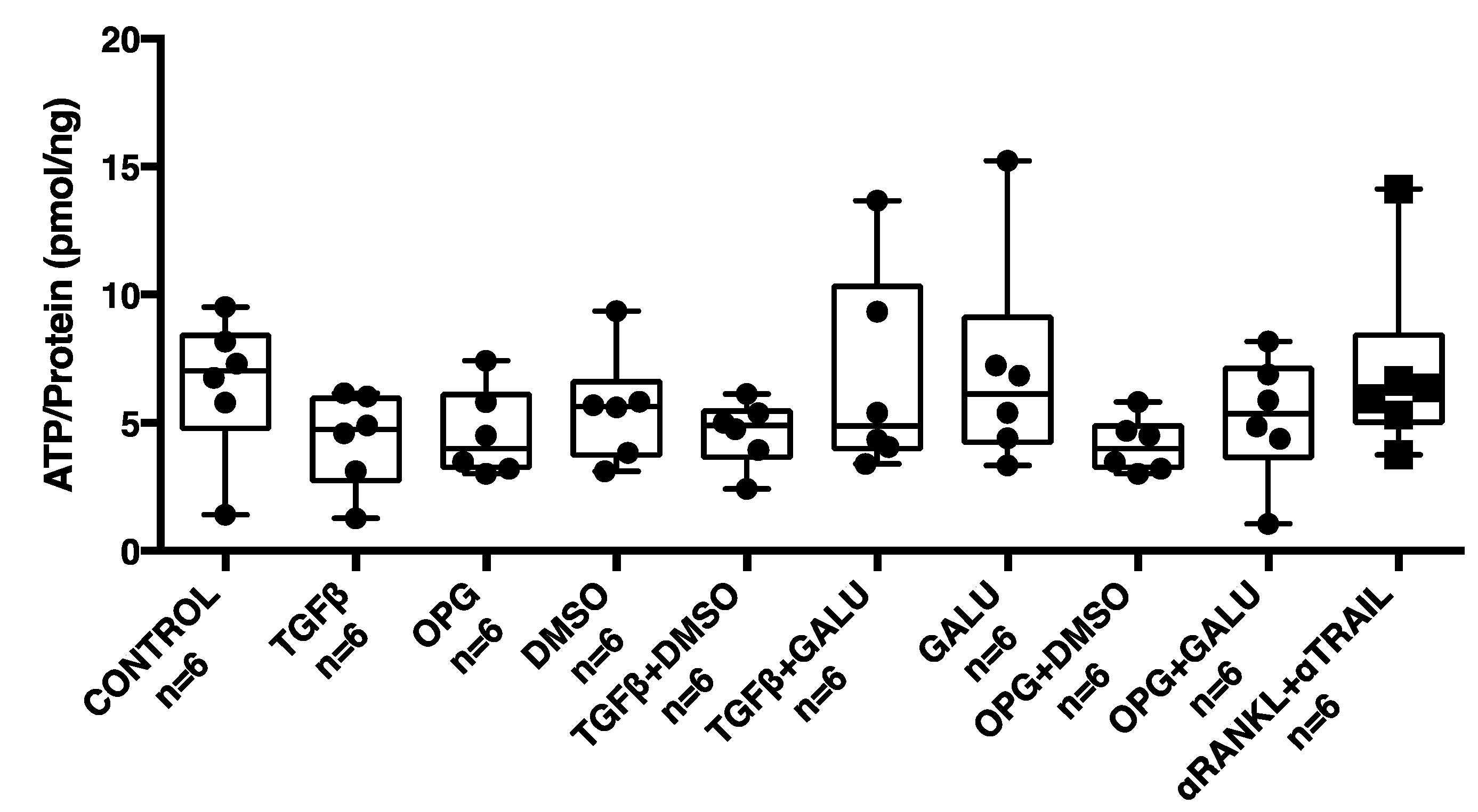
3.4. OPG Treatment of Mouse Precision-Cut Liver Slices Results in Higher Expression of Fibrosis-Associated Markers through TGFβ1
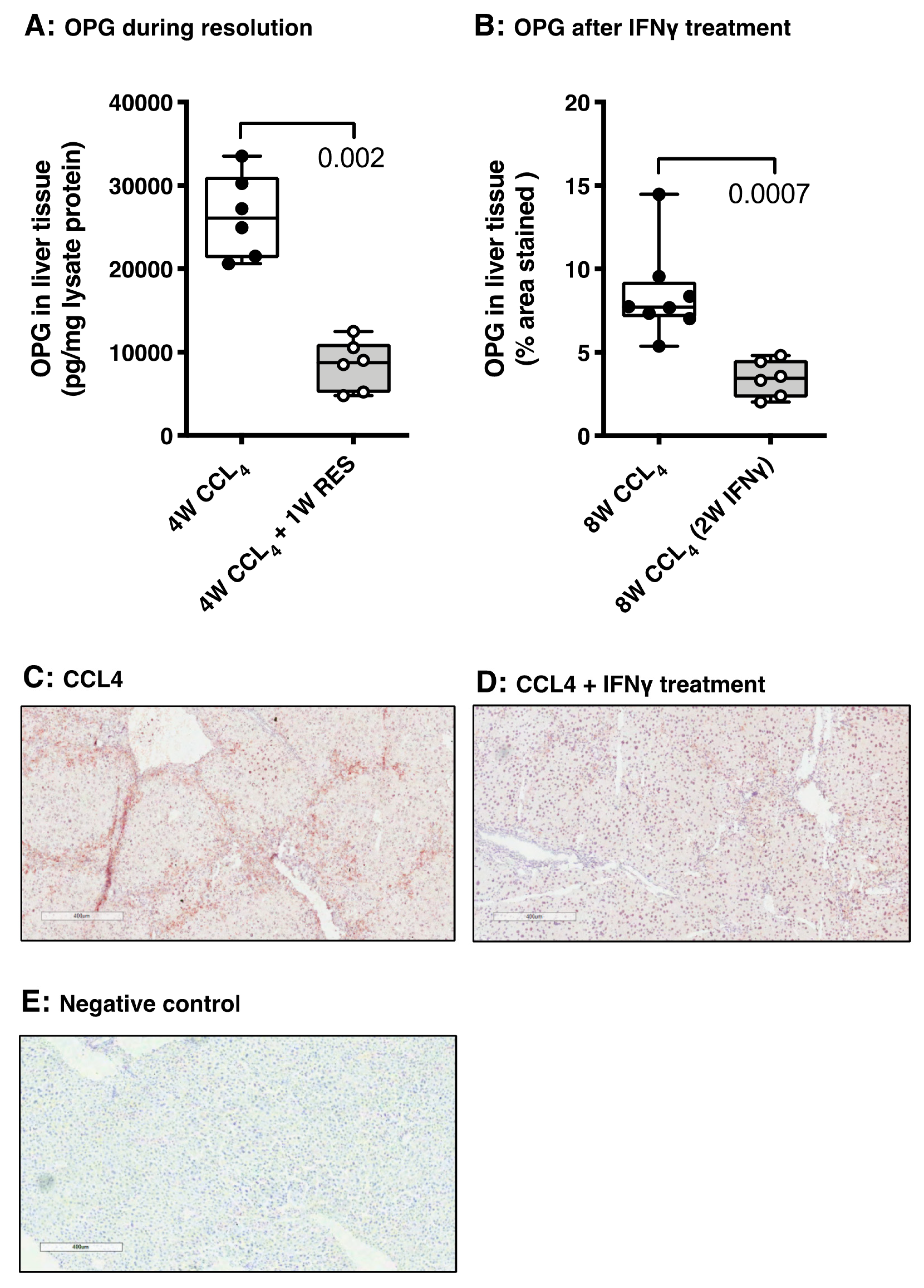
3.5. OPG Expression Responds to Spontaneous and Drug-Induced Fibrosis Resolution
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| β-actin | ATCGTGCGTGACATCAAAGA | ATGCCACAGGATTCCATACC |
| Col1α1 | TGACTGGAAGAGCGGAGAGT | ATCCATCGGTCATGCTCTCT |
| αSMA | ACTACTGCCGAGCGTGAGAT | CCAATGAAAGATGGCTGGAA |
| Fn1 | CGGAGAGAGTGCCCCTACTA | CGATATTGGTGAATCGCAGA |
| OPG | ACAGTTTGCCTGGGACCAAA | CTGTGGTGAGGTTCGAGTGG |
| TGFβ1 | AGGGCTACCATGCCAACTTC | GTTGGACAACTGCTCCACCT |
| HSP47 | AGGTCACCAAGGATGTGGAG | CAGCTTCTCCTTCTCGTCGT |

References
- Friedman, S.L. Liver fibrosis—from bench to bedside. J. Hepatol. 2003, 38, S38–S53. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Ge, P.S.; Runyon, B.A. Treatment of patients with cirrhosis. N. Engl. J. Med. 2016, 375, 767–777. [Google Scholar] [CrossRef] [Green Version]
- Schuppan, D.; Kim, Y.O. Evolving therapies for liver fibrosis. J. Clin. Investig. 2013, 123, 1887–1901. [Google Scholar] [CrossRef] [Green Version]
- Fallatah, H.I. Noninvasive biomarkers of liver fibrosis: An overview. Adv. Hepatol. 2014, 2014, 357287. [Google Scholar] [CrossRef]
- Motola, D.L.; Caravan, P.; Chung, R.T.; Fuchs, B.C. Noninvasive biomarkers of liver fibrosis: Clinical applications and future directions. Curr. Pathobiol. Rep. 2015, 2, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, Y.; Yonal, O.; Kurt, R.; Oral, A.Y.; Eren, F.; Ozdogan, O.; Ari, F.; Celikel, C.A.; Korkmaz, S.; Ulukaya, E.; et al. Serum levels of osteoprotegerin in the spectrum of nonalcoholic fatty liver disease. Scand. J. Clin. Lab. Investig. 2010, 70, 541–546. [Google Scholar] [CrossRef]
- García-Valdecasas-Campelo, E.; González-Reimers, E.; Santolaria-Fernández, F.; De La Vega-Prieto, M.J.; Milena-Abril, A.; Sanchez-Perez, M.J.; Martínez-Riera, A.; Gómez-Rodríguez, M.D.L.A. Serum osteoprotegerin and RANKL levels in chronic alcoholic liver disease. Alcohol Alcohol. 2006, 41, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Prystupa, A.; Dąbrowska, A.; Sak, J.J.; Toruń-Jurkowska, A.; Lachowska-Kotowska, P.; Dzida, G. Concentration of fetuin-A, osteoprotegerin and α-Klotho in patients with alcoholic liver cirrhosis. Exp. Ther. Med. 2016, 12, 3464–3470. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Xu, D.; Liu, Y.; Guo, X.; Li, W.; Guo, C.; Zhang, H.; Gao, Y.; Mao, Y.; Zhao, J. Combined serum biomarkers in non-invasive diagnosis of non-alcoholic steatohepatitis. PLoS ONE 2015, 10, e0131664. [Google Scholar] [CrossRef] [Green Version]
- Guañabens, N.; Enjuanes, A.; Alvarez, L.; Peris, P.; Caballería, L.; Jesús martinez de Osaba, M.; Cerdá, D.; Monegal, A.; Pons, F.; Parés, A. High osteoprotegerin serum levels in primary billiary cirrhosis are associated with disease severity but not with the mRNA gene expression in liver tissue. J. Bone Miner. Metab. 2009, 27, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Monegal, A.; Navasa, M.; Peris, P.; Alvarez, L.; Pons, F.; Rodés, J.; Guañabens, N. The RANKL/OPG system and bone mineral density in patients with chronic liver disease. Liver Int. 2007, 27, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Kaser, A.; Stadlmann, S.; Millonig, G.; Kaser, S.; Mühllechner, P.; Habior, A.; Graziadei, I.; Vogel, W.; Tilg, H. The RANKL/OPG system and bone mineral density in patients with chronic liver disease. J. Hepatol. 2005, 43, 973–983. [Google Scholar] [CrossRef]
- Fábrega, E.; Orive, A.; García-Suarez, C.; García-Unzueta, M.; Antonio Amado, J.; Pons-Romero, F. Osteoprotegerin and RANKL in alcoholic liver cirrhosis. Liver Int. 2005, 25, 305–310. [Google Scholar] [CrossRef]
- Szalay, F.; Hegedus, D.; Lakatos, P.L.; Tornai, I.; Bajnok, E.; Dunkel, K.; Lakatos, P. High serum osteoprotegerin and low RANKL in primary billary cirrhosis. J. Hepatol. 2003, 38, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Bosselut, N.; Taibi, L.; Guéchot, J.; Zarski, J.P.; Sturm, N.; Gelineau, M.C.; Poggi, B.; Thoret, S.; Lasnier, E.; Baudin, B.; et al. The ANRS HCEP 23 Fibrostar Group. Including osteoprotegerin and collagen IV in a score-based blood test for liver fibrosis increases diagnostic accuracy. Clin. Chim. Acta 2013, 415, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Vitovski, S.; Phillips, J.S.; Sayers, J.; Croucher, P.I. Investigating the interaction between osteoprotegerin and receptor activator of NFκB or tumor necrosis factor-related apoptosis-inducing ligand: Evidence for a pivotal role for osteoprotegerin in regulating two distinct pathways. J. Biol. Chem. 2007, 282, 31601–31609. [Google Scholar] [CrossRef] [Green Version]
- Boyce, B.F.; Xing, L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res. Ther. 2007, 9 (Suppl. S1), S1. [Google Scholar] [CrossRef] [Green Version]
- Bock, O.; Loch, G.; Schade, U.; Büsche, G.; von Wasielewski, R.; Wiese, B.; Kreipe, H. Osteosclerosis in advanced chronic idiopathic myelofibrosis is associated with endothelial overexpression of osteoprotegerin. Br. J. Haematol. 2005, 130, 76–82. [Google Scholar] [CrossRef]
- Toffoli, B.; Pickering, R.J.; Tsorotes, D.; Wang, B.; Bernardi, S.; Kantharidis, P.; Fabris, B.; Zauli, G.; Secchiero, P.; Thomas, M.C. Osteoprotegerin promotes vascular fibrosis via a TGF-I1 autocrine loop. Atherosclerosis 2011, 218, 61–68. [Google Scholar] [CrossRef]
- Cross, S.S.; Yang, Z.; Brown, N.J.; Balasubramanian, S.P.; Evans, C.A.; Woodward, J.K.; Neville-Webbe, H.L.; Lippitt, J.M.; Reed, M.W.R.; Coleman, R.E.; et al. Osteoprotegerin (OPG)—a potential new role in the regulation of endothelial cell phenotype and tumour angiogenesis? Int. J. Cancer 2006, 118, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Holen, I.; Cross, S.S.; Neville-Webbe, H.L.; Cross, N.A.; Balasubramanian, S.P.; Croucher, P.I.; Evans, C.A.; Lippitt, J.M.; Coleman, R.E.; Eaton, C.L. Osteoprotegerin (OPG) expression by breast cancer cells in vitro and breast tumours in vivo—a role in tumour survival? Breast Cancer Res. Treat. 2005, 92, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Ölander, M.; Wisniewski, J.R.; Artursson, P. Cell-type-resolved proteomic analysis of human liver. Liver Int. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boorsma, C.E.; Draijer, C.; Cool, R.; Brandsma, C.; Nossent, G.; Brass, D.M.; Timens, W.; Melgert, B.M. A possible role for the RANK/RANKL/OPG axis in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2014, 189, A1252. [Google Scholar]
- Liu, W.; Feng, W.; Wang, F.; Li, W.; Gao, C.; Zhou, B.; Ma, M. Osteoprotegerin/RANK/RANKL axis in cardiac remodelling due to immuno-inflammatory myocardiac disease. Exp. Mol. Pathol. 2008, 84, 213–217. [Google Scholar] [CrossRef]
- Vidal, K.; Serrant, P.; Schlosser, B.; van den Broek, P.; Lorget, F.; Donnet-Hughes, A. Osteoprotegerin production by human interstitial epithelial cells: A potential regulator of mucosal immune responses. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G836–G844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Fu, M.; Myles, D.; Zhu, X.; Du, J.; Cao, X.; Chen, Y.E. PDGF induces osteoprotegerin expression in vascular smooth muscle cells by multiple signal pathways. FEBS Lett. 2002, 521, 180–184. [Google Scholar] [CrossRef] [Green Version]
- Bansal, R.; Prakash, J.; Post, E.; Beljaars, L.; Schuppan, D.; Poelstra, K. Novel engineered targeted interferron-gamma blocks hepatic fibrogenesis in mice. Hepatology 2011, 54, 586–596. [Google Scholar] [CrossRef]
- Beljaars, L.; Schippers, M.; Reker-smit, C.; Martinez, F.O.; Helming, L.; Poelstra, K.; Melgert, B.N. Hepatic localization of macrophage phenotypes during fibrogenesis and resolution of fibrosis in mice and humans. Front. Immunol. 2014, 5, 430. [Google Scholar] [CrossRef] [Green Version]
- De Graaf, I.A.M.; Olinga, P.; De Jager, M.H.; Merema, M.T.; De Kanter, R.; Van de Kerkhof, E.G.; Groothuis, G.M.M. Preparation and incubation of precision-cut liver and intestinal slices for application in drug metabolism and toxicity studies. Nat. Protoc. 2010, 5, 1540–1551. [Google Scholar] [CrossRef]
- Hadi, M.; Chen, Y.; Starokozhko, V.; Merema, M.T.; Groothuis, G.M.M. Mouse precision-cut liver slices as an ex vivo model to study idiosyncratic drug-induced liver injury. Chem. Res. Toxicol. 2012, 25, 1938–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terkelsen, M.K.; Bendixen, S.M.; Hansen, D.; Scott, E.A.H.; Moeller, A.F.; Nielsen, R.; Mandrup, S.; Schlosser, A.; Andersen, T.L.; Sorensen, G.L.; et al. Transcriptional dynamics of hepatic sinusoid-associated cells after liver injury. Hepatology 2020. accepted author manuscript. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimifar, S.B.; Nagaraj, N.; Cox, J.; Mann, M. Cell-type-resolved quantitative proteomics of murine liver. Cell Metab. 2014, 20, 1076–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, C.; Li, Y.; Guo, F.; Jiang, Y.; Ying, W.; Li, D.; Yang, D.; Xia, X.; Liu, W.; Zhao, Y.; et al. A cell type-resolved liver proteome. Mol. Cell. Proteom. 2016, 15, 3190–3202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, N.; Van Sweringen, H.L.; Schuster, R.; Blanchard, J.; Burns, J.M.; Tevar, A.D.; Edwards, M.J.; Lentsch, A.B. Receptor Activator of Nuclear Factor-κB ligand (RANKL) protects against hepatic ischemia/reperfusion injury in mice. Hepatology 2012, 55, 888–897. [Google Scholar] [CrossRef] [Green Version]
- Dawson, S.; Lawrie, A. From bones to blood pressure, developing novel biologic approaches targeting the osteoprotegerin pathway for pulmonary vascular disease. Pharmacol. Ther. 2017, 169, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Lemke, J.; von Karstedt, S.; Zinngrebe, J.; Walczak, H. Getting TRAIL back on track for cancer therapy. Cell Death Differ. 2014, 21, 1350–1364. [Google Scholar] [CrossRef] [Green Version]
- Taimr, P.; Higuchi, H.; Kocova, E.; Rippe, R.A.; Friedman, S.; Gores, G.J. Activated stellate cells express the TRAIL receptor-2/death receptor-5 cancer-specific apoptosis within a therapeutic window. Hepatology 2007, 45, 649–658. [Google Scholar]
- Chamoux, E.; Houde, N.; L’Eriger, K.; Roux, S. Osteoprotegerin decreases human osteoclast apoptosis by inhibiting the TRAIL pathway. J. Cell. Physiol. 2008, 216, 536–542. [Google Scholar] [CrossRef]
- Marshall, R.P.; Simpson, J.K.; Lukey, P.T. Strategies for biomarker discovery in fibrotic disease. Biochim. Biophys. Acta Mol. Basis Dis. 2013, 1832, 1079–1087. [Google Scholar] [CrossRef] [Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhyatmika, A.; Beljaars, L.; Putri, K.S.S.; Habibie, H.; Boorsma, C.E.; Reker-Smit, C.; Luangmonkong, T.; Guney, B.; Haak, A.; Mangnus, K.A.; et al. Osteoprotegerin Is more than a Possible Serum Marker in Liver Fibrosis: A Study into Its Function in Human and Murine Liver. Pharmaceutics 2020, 12, 471. https://doi.org/10.3390/pharmaceutics12050471
Adhyatmika A, Beljaars L, Putri KSS, Habibie H, Boorsma CE, Reker-Smit C, Luangmonkong T, Guney B, Haak A, Mangnus KA, et al. Osteoprotegerin Is more than a Possible Serum Marker in Liver Fibrosis: A Study into Its Function in Human and Murine Liver. Pharmaceutics. 2020; 12(5):471. https://doi.org/10.3390/pharmaceutics12050471
Chicago/Turabian StyleAdhyatmika, Adhyatmika, Leonie Beljaars, Kurnia S. S. Putri, Habibie Habibie, Carian E. Boorsma, Catharina Reker-Smit, Theerut Luangmonkong, Burak Guney, Axel Haak, Keri A. Mangnus, and et al. 2020. "Osteoprotegerin Is more than a Possible Serum Marker in Liver Fibrosis: A Study into Its Function in Human and Murine Liver" Pharmaceutics 12, no. 5: 471. https://doi.org/10.3390/pharmaceutics12050471