
Zika Virus: A Neurotropic Warrior against High-Grade Gliomas—Unveiling Its Potential for Oncolytic Virotherapy
 , , and
, , and
Abstract
:1. Introduction
2. Biology of GBM
3. The Oncolytic Viruses
- (i)
- Tumor Tropism: the virus’s ability to selectively target and infect tumor cells.
- (ii)
- Infection of Non-tumoral Cells: whether the virus can infect non-tumoral cells or is specific to cancer cells.
- (iii)
- Encoding Therapeutic Transgenes: the capacity of the virus to carry and express therapeutic transgenes.
- (iv)
- Replication and Viral Progeny Production: the ability of the virus to replicate and generate viral progeny, including viremia in blood and tissues.
- (v)
- Virus Infiltration into Tissues: the extent to which the virus can infiltrate tissues.
- (vi)
- Viral Evasion of the Immune Response: the virus’s ability to evade the host immune response.
- (vii)
- Minimizing Adverse Effects: strategies aimed at minimizing potential adverse effects.
- (viii)
- Virus Pathogenicity and Immunogenicity: the virus’s inherent pathogenicity and its potential to induce an immune response.
4. New Therapies for GBM Treatment
5. ZIKV Therapy in GBM
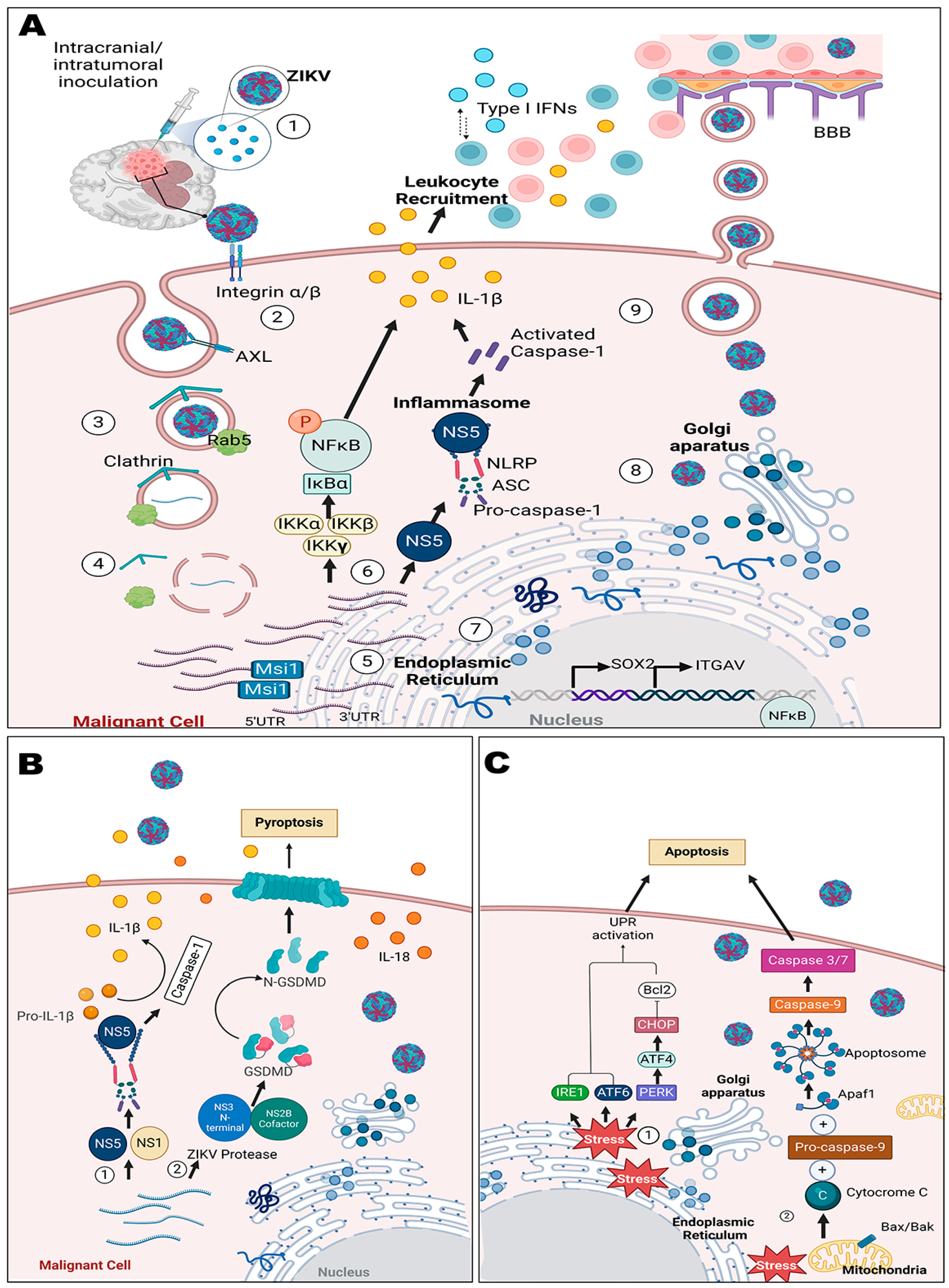
5.1. Mechanisms in CNS Tumors
5.2. Cellular and Animal Models
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AdV | adenovirus |
| bp | base pair |
| BBB | blood–brain barrier |
| CEA | carcinoembryonic antigen |
| CNS | central nervous system |
| CSC | cancer stem cells |
| CLT | Clinicaltrials.gov ID |
| EGFR | epidermal growth factor receptor |
| EMA | European Medicines Agency. |
| Eng | engineered |
| ER | endoplasmic reticulum |
| FDA | United States Food and Drug Administration Agency |
| GBM | glioblastoma multiforme |
| GBM-I | glioblastoma multiforme type 1 |
| GBM-II | glioblastoma multiforme type 2 |
| GBR | glioblastoma recurrence |
| GFAP | glial fibrillary acidic protein |
| GSDMD | gasdermin D protein |
| GSC | glioma stem cells |
| HR2V | human rhinovirus type 2 |
| HSV | herpes simplex virus |
| IDH1 | isocitrate dehydrogenase type 1 enzyme |
| IDH2 | isocitrate dehydrogenase type 2 enzyme |
| IFN | interferon |
| IL-1β | interleukin-1β |
| IL-18 | interleukin-18 |
| IRES | internal ribosomal entry site |
| MGMT | O-6-Methylguanine-DNA-methyltransferase gene |
| MV | measles virus |
| mRNA | messenger RNA |
| ncRNA | non-coding RNAs |
| NPC | neural progenitor cells |
| NP/EE | Nestin Promoter/Enhancer Element |
| NK | natural killer cells |
| nsIL-12 | non-secretory interleukin-12 |
| OVT | oncolytic virotherapy |
| OV | oncolytic virus |
| PDGF | platelet-derived growth factor |
| PDGFRα | receptor alpha of the PDGF |
| PTEN | phosphatase and tensin gene |
| PVS1 | live attenuated poliovirus serotype 1 (SABIN) vaccine |
| RGD | specific amino acid sequence Arg-Gly-Asp |
| REO | reovirus |
| RR | ribonucleotide reductase |
| RRV | retroviral replicating vector |
| RV | rotavirus |
| TK | thymidine kinase |
| TSGs | tumor suppressor genes |
| T-VEC | Talimogene laherparepvec |
| TMZ | Temozolomide |
| VEGF | vascular endothelial growth factor |
| WHO | World Health Organization |
| WT | wild-type |
| ZIKV | Zika virus |
References
- Bhagat, R.; Kaur, G.; Seth, P. Molecular Mechanisms of Zika Virus Pathogenesis: An Update. Indian J. Med. Res. 2021, 154, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Mittal, R.; Nguyen, D.; Debs, L.H.; Patel, A.P.; Liu, G.; Jhaveri, V.M.; Kay, S.I.S.; Mittal, J.; Bandstra, E.S.; Younis, R.T.; et al. Zika Virus: An Emerging Global Health Threat. Front. Cell. Infect. Microbiol. 2017, 7, 486. [Google Scholar] [CrossRef]
- Rabaan, A.A.; Bazzi, A.M.; Al-Ahmed, S.H.; Al-Ghaith, M.H.; Al-Tawfiq, J.A. Overview of Zika Infection, Epidemiology, Transmission and Control Measures. J. Infect. Public Health 2017, 10, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Saiz, J.C.; Martín-Acebes, M.A.; Bueno-Marí, R.; Salomón, O.D.; Villamil-Jiménez, L.C.; Heukelbach, J.; Alencar, C.H.; Armstrong, P.K.; Ortiga-Carvalho, T.M.; Mendez-Otero, R.; et al. Zika Virus: What Have We Learnt since the Start of the Recent Epidemic? Front. Microbiol. 2017, 8, 1554. [Google Scholar] [CrossRef]
- Bhardwaj, U.; Pandey, N.; Rastogi, M.; Singh, S.K. Gist of Zika Virus Pathogenesis. Virology 2021, 560, 86–95. [Google Scholar] [CrossRef]
- Petersen, L.; Jamieson, D.; Powers, A.; Honein, M. Zika virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Song, B.H.; Yun, S.I.; Woolley, M.; Lee, Y.M. Zika Virus: History, Epidemiology, Transmission, and Clinical Presentation. J. Neuroimmunol. 2017, 308, 50–64. [Google Scholar] [CrossRef]
- Zhu, Z.; Gorman, M.J.; McKenzie, L.D.; Chai, J.N.; Hubert, C.G.; Prager, B.C.; Fernandez, E.; Richner, J.M.; Zhang, R.; Shan, C.; et al. Zika Virus Has Oncolytic Activity against Glioblastoma Stem Cells. J. Exp. Med. 2017, 214, 2843–2857. [Google Scholar] [CrossRef]
- Lubin, J.A.; Zhang, R.R.; Kuo, J.S. Zika Virus Has Oncolytic Activity against Glioblastoma Stem Cells. Clin. Neurosurg. 2018, 82, E113–E114. [Google Scholar] [CrossRef]
- Hanif, F.; Muzaffar, K.; Perveen, K.; Malhi, S.M.; Simjee, S.U. Glioblastoma Multiforme: A Review of Its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac. J. Cancer Prev. 2017, 18, 3–9. [Google Scholar] [CrossRef]
- Weller, M.; van den Bent, M.; Preusser, M.; Le Rhun, E.; Tonn, J.C.; Minniti, G.; Bendszus, M.; Balana, C.; Chinot, O.; Dirven, L.; et al. EANO Guidelines on the Diagnosis and Treatment of Diffuse Gliomas of Adulthood. Nat. Rev. Clin. Oncol. 2021, 18, 170–186. [Google Scholar] [CrossRef]
- Contreras, L.E. Epidemiología De Tumores Cerebrales. Rev. Méd. Clín. Condes 2017, 28, 332–338. [Google Scholar] [CrossRef]
- Altshuler, D.B.; Kadiyala, P.; Núñez, F.J.; Núñez, F.M.; Alghamri, M.S.; Garcia-fabiani, M.B.; Asad, A.S.; Candia, A.J.N.; Candolfi, M.; Lahann, J.; et al. Prospects of Biological and Synthetic Pharmacotherapies for Glioblastoma. Expert Opin. Biol. Ther. 2020, 20, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Miyauchi, J.T.; Tsirka, S.E. Advances in Immunotherapeutic Research for Glioma Therapy. J. Neurol. 2018, 265, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Martikainen, M.; Essand, M. Virus-Based Immunotherapy of Glioblastoma. Cancers 2019, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Foreman, P.M.; Friedman, G.K.; Cassady, K.A.; Markert, J.M. Oncolytic Virotherapy for the Treatment of Malignant Glioma. Neurotherapeutics 2017, 14, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.Q.; Wang, S.B.; Cai, M.H.; Wang, X.J.; Chen, J.Y.; Tong, X.M.; Chen, X.Y.; Mou, X.Z. Recent Advances in Oncolytic Virus-Based Cancer Therapy. Virus Res. 2012, 270, 197675. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.N.; Ling, F. Zika Virus Infection and Microcephaly: Evidence for a Causal Link. Int. J. Environ. Res. Public Health 2016, 13, 1031. [Google Scholar] [CrossRef] [PubMed]
- Salacz, M.E.; Watson, K.R.; Schomas, D.A. Glioblastoma: Part I. Current State of Affairs. Mo. Med. 2011, 108, 187–194. [Google Scholar] [PubMed]
- da Cunha, M.L.V.; Maldaun, M.V.C. Metastasis from Glioblastoma Multiforme: A Meta-Analysis. Rev. Assoc. Med. Bras. 2019, 65, 424–433. [Google Scholar] [CrossRef]
- Alexander, B.M.; Cloughesy, T.F. Adult Glioblastoma. J. Clin. Oncol. 2017, 35, 2402–2409. [Google Scholar] [CrossRef]
- Rahman, M.; Dastmalchi, F.; Karachi, A.; Mitchell, D. The Role of CMV in Glioblastoma and Implications for Immunotherapeutic Strategies. Oncoimmunology 2019, 8, e1514921. [Google Scholar] [CrossRef]
- Davis, M.E. Glioblastoma: Overview of Disease and Treatment. Clin. J. Oncol. Nurs. 2016, 20, S2–S8. [Google Scholar] [CrossRef]
- Karpel-Massler, G.; Ishida, C.T.; Bianchetti, E.; Zhang, Y.; Shu, C.; Tsujiuchi, T.; Banu, M.A.; Garcia, F.; Roth, K.A.; Bruce, J.N.; et al. Induction of Synthetic Lethality in IDH1-Mutated Gliomas through Inhibition of Bcl-XL. Nat. Commun. 2017, 8, 1067. [Google Scholar] [CrossRef]
- Lau, D.; Magill, S.T.; Aghi, M.K. Molecularly Targeted Therapies for Recurrent Glioblastoma: Current and Future Targets. Neurosurg. Focus 2014, 37, E15. [Google Scholar] [CrossRef] [PubMed]
- Campos, B.; Olsen, L.R.; Urup, T.; Poulsen, H.S. A Comprehensive Profile of Recurrent Glioblastoma. Oncogene 2016, 35, 5819–5825. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.W.; Chen, C.C. Photodynamic Therapy for the Treatment of Glioblastoma. Front Surg. 2020, 6, 81. [Google Scholar] [CrossRef] [PubMed]
- Parashar, D.; Singh, A.; Gupta, S.; Sharma, A.; Sharma, M.K.; Roy, K.K.; Chauhan, S.C.; Kashyap, V.K. Emerging Roles and Potential Applications of Non-Coding RNAs in Cervical Cancer. Genes 2022, 13, 1254. [Google Scholar] [CrossRef] [PubMed]
- Fukuhara, H.; Ino, Y.; Todo, T. Oncolytic Virus Therapy: A New Era of Cancer Treatment at Dawn. Cancer Sci. 2016, 107, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Prestwich, R.J.D.; Harrington, K.J.; Pandha, H.S.; Vile, R.G.; Melcher, A.A.; Errington, F. Oncolytic Viruses: A Novel Form of Immunotherapy. Expert Rev. Anticancer Ther. 2008, 8, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Stavrakaki, E.; Dirven, C.M.F.; Lamfers, M.L.M. Personalizing Oncolytic Virotherapy for Glioblastoma: In Search of Biomarkers for Response. Cancers 2021, 13, 614. [Google Scholar] [CrossRef]
- Li, K.; Zhao, Y.; Hu, X.; Jiao, J.; Wang, W.; Yao, H. Advances in the Clinical Development of Oncolytic Viruses. Am. J. Transl. Res. 2022, 314, 4192–4206. [Google Scholar]
- Manchester, M.; Eto, D.S.; Valsamakis, A.; Liton, P.B.; Rota, P.A.; Bellini, W.J.; Fernandez-mun, R.; Forthal, D.N.; Oldstone, M.B.A.; Irol, J.V. Clinical Isolates of Measles Virus Use CD46 as a Cellular Receptor. J. Virol. 2000, 74, 3967–3974. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, W.; Cai, W.; Jiang, K.; Yuan, Z. Engineered Measles Virus Edmonston Strain Used as a Novel Oncolytic Viral System against Human Hepatoblastoma. BMC Cancer 2012, 12, 427. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Berkeley, R.; Barr, T.; Ilett, E.; Errington-Mais, F. Past, Present and Future of Oncolytic Reovirus. Cancers 2020, 12, 3219. [Google Scholar] [CrossRef]
- Boisgerault, N.; Tangy, F.; Gregoire, M. New Perspectives in Cancer Virotherapy: Bringing the Immune System into Play. Immunotherapy 2010, 2, 185–199. [Google Scholar] [CrossRef]
- Brammer, J.; Rosenthal, K.S. Therapeutic Smart Bombs. Infect. Dis. Clin. Pract. 2009, 17, 39–43. [Google Scholar] [CrossRef]
- Lauer, U.M. Oncolytic Viruses: Challenges and Considerations in an Evolving Clinical Landscape. Future Oncol. 2022, 18, 2713–2732. [Google Scholar] [CrossRef]
- Buijs, P.R.A.; Verhagen, J.H.E.; van Eijck, C.H.J.; van den Hoogen, B.G. Oncolytic Viruses: From Bench to Bedside with a Focus on Safety. Hum. Vaccines Immunother. 2015, 11, 1573–1584. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, C.; Miao, J.; Wang, Z.; Wang, Z.; Cheng, Z.; Wang, P.; Dunmall, L.S.C.; Lemoine, N.R.; Wang, Y. A Tumor-Targeted Replicating Oncolytic Adenovirus Ad-TD-NsIL12 as a Promising Therapeutic Agent for Human Esophageal Squamous Cell Carcinoma. Cells 2020, 9, 2438. [Google Scholar] [CrossRef]
- Bortolanza, S.; Bunuales, M.; Otano, I.; Gonzalez-Aseguinolaza, G.; Ortiz-de-Solorzano, C.; Perez, D.; Prieto, J.; Hernandez-Alcoceba, R. Treatment of Pancreatic Cancer with an Oncolytic Adenovirus Expressing Interleukin-12 in Syrian Hamsters. Mol. Ther. 2009, 17, 614–622. [Google Scholar] [CrossRef]
- Wang, P.; Li, X.; Wang, J.; Gao, D.; Li, Y.; Li, H.; Chu, Y.; Zhang, Z.; Liu, H.; Jiang, G.; et al. Re-Designing Interleukin-12 to Enhance Its Safety and Potential as an Anti-Tumor Immunotherapeutic Agent. Nat. Commun. 2017, 8, 1395. [Google Scholar] [CrossRef]
- Nassiri, F.; Patil, V.; Yefet, L.S.; Singh, O.; Liu, J.; Dang, R.M.A.; Yamaguchi, T.N.; Daras, M.; Cloughesy, T.F.; Colman, H.; et al. Oncolytic DNX-2401 Virotherapy plus Pembrolizumab in Recurrent Glioblastoma: A Phase 1/2 Trial. Nat. Med. 2013, 29, 1370–1378. [Google Scholar] [CrossRef]
- Faisal, S.M.; Castro, M.G.; Lowenstein, P.R. Combined Cytotoxic and Immune-Stimulatory Gene Therapy Using Ad-TK and Ad-Flt3L: Translational Developments from Rodents to Glioma Patients. Mol. Ther. 2023, 1, 2839–2860. [Google Scholar] [CrossRef]
- Markert, J.M.; Medlock, M.D.; Rabkin, S.D.; Gillespie, G.Y.; Todo, T.; Hunter, W.D.; Palmer, C.A.; Feigenbaum, F.; Tornatore, C.; Tufaro, F.; et al. Conditionally Replicating Herpes Simplex Virus Mutant G207 for the Treatment of Malignant Glioma: Results of a Phase I Trial. Gene Ther. 2000, 7, 867–874. [Google Scholar] [CrossRef]
- Yun, C.O.; Hong, J.W.; Yoon, A.R. Current Clinical Landscape of Oncolytic Viruses as Novel Cancer Immunotherapeutic and Recent Preclinical Advancements. Front. Immunol. 2022, 13, 953410. [Google Scholar] [CrossRef]
- Chiocca, E.A.; Nakashima, H.; Kasai, K.; Fernandez, S.A.; Oglesbee, M. Preclinical Toxicology of RQNestin34.5v.2: An Oncolytic Herpes Virus with Transcriptional Regulation of the ICP34.5 Neurovirulence Gene. Mol. Ther. Methods Clin. Dev. 2020, 17, 871–893. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.M.; Foreman, P.M.; Nabors, L.B.; Riley, K.O.; Gillespie, G.Y.; Markert, J.M. Design of a Phase I Clinical Trial to Evaluate M032, a Genetically Engineered HSV-1 Expressing IL-12, in Patients with Recurrent/Progressive Glioblastoma Multiforme, Anaplastic Astrocytoma, or Gliosarcoma. Hum. Gene Ther. Clin. Dev. 2016, 27, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Rius-Rocabert, S.; García-Romero, N.; García, A.; Ayuso-Sacido, A.; Nistal-Villan, E. Oncolytic Virotherapy in Glioma Tumors. Int. J. Mol. Sci. 2020, 21, 7604. [Google Scholar] [CrossRef] [PubMed]
- Geletneky, K.; Huesing, J.; Rommelaere, J.; Schlehofer, J.R.; Leuchs, B.; Dahm, M.; Krebs, O.; von Knebel Doeberitz, M.; Huber, B.; Hajda, J. Phase I/IIa Study of Intratumoral/Intracerebral or Intravenous/Intracerebral Administration of Parvovirus H-1 (ParvOryx) in Patients with Progressive Primary or Recurrent Glioblastoma Multiforme: ParvOryx01 Protocol. BMC Cancer 2012, 12, 99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Xia, Q.; Liu, L.; Li, S.; Dong, L. Current Opinion on Molecular Characterization for GBM Classification in Guiding Clinical Diagnosis, Prognosis, and Therapy. Front. Mol. Biosci. 2020, 7, 562798. [Google Scholar] [CrossRef]
- Rivera, A.L.; Pelloski, C.E.; Gilbert, M.R.; Colman, H.; De La Cruz, C.; Sulman, E.P.; Bekele, B.N.; Aldape, K.D. MGMT Promoter Methylation Is Predictive of Response to Radiotherapy and Prognostic in the Absence of Adjuvant Alkylating Chemotherapy for Glioblastoma. Neuro-Oncol. 2010, 12, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Ashkan, K.; Baig Mirza, A.; Soumpasis, C.; Syrris, C.; Kalaitzoglou, D.; Sharma, C.; James, Z.J.; Khoja, A.K.; Ahmed, R.; Vastani, A.; et al. MGMT Promoter Methylation: Prognostication beyond Treatment Response. J. Pers. Med. 2023, 13, 999. [Google Scholar] [CrossRef] [PubMed]
- Ricaurte, O.; Neita, K.; Valero, D.; Ortega-Rojas, J.; Arboleda-Bustos, C.E.; Zubieta, C.; Penagos, J.; Arboleda, G. Estudio de Mutaciones en los Genes IDH 1 y 2 en una Muestra de Gliomas en Población Colombiana. Biomédica 2018, 38, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Wang, Y.; Ge, P.; Bailey, C.; Zhang, P.; Zhang, D.; Meng, Z.; Qi, C.; Chen, Q.; Chen, J.; et al. The HIF1α-PDGFD-PDGFRα Axis Controls Glioblastoma Growth at Normoxia/Mild-Hypoxia and Confers Sensitivity to Targeted Therapy by Echinomycin. J. Exp. Clin. Cancer Res. 2021, 40, 278. [Google Scholar] [CrossRef]
- Minniti, G.; De Sanctis, V.; Muni, R.; Filippone, F.; Bozzao, A.; Valeriani, M.; Osti, M.F.; De Paula, U.; Lanzetta, G.; Tombolini, V.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma in Elderly Patients. J. Neurooncol. 2008, 88, 97–103. [Google Scholar] [CrossRef]
- Alonso, M.M.; Jiang, H.; Gomez-Manzano, C.; Fueyo, J. Targeting Brain Tumor Stem Cells with Oncolytic Adenoviruses. Methods Mol. Biol. 2012, 797, 111–125. [Google Scholar] [CrossRef]
- Cattaneo, R.; Russell, S.J. How to Develop Viruses into Anticancer Weapons. PLoS Pathog. 2017, 13, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Boagni, D.A.; Ravirala, D.; Zhang, S.X. Current Strategies in Engaging Oncolytic Viruses with Antitumor Immunity. Mol. Ther. Oncolytics 2021, 22, 98–113. [Google Scholar] [CrossRef]
- Zheng, M.; Huang, J.; Tong, A.; Yang, H. Oncolytic Viruses for Cancer Therapy: Barriers and Recent Advances. Mol. Ther. Oncolytics 2019, 15, 234–247. [Google Scholar] [CrossRef]
- Tang, H.; Hammack, C.; Ogden, S.C.; Wen, Z.; Qian, X.; Li, Y.; Yao, B.; Shin, J.; Zhang, F.; Lee, E.M.; et al. Zika Virus Infects Human Cortical Neural Progenitors and Attenuates Their Growth. Cell Stem Cell 2016, 18, 587–590. [Google Scholar] [CrossRef]
- Rubio-Hernández, E.I.; Comas-García, M.; Coronado-Ipiña, M.A.; Colunga-Saucedo, M.; Sánchez, H.M.G.; Castillo, C.G. Astrocytes Derived from Neural Progenitor Cells Are Susceptible to Zika Virus Infection. PLoS ONE 2023, 18, e0283429. [Google Scholar] [CrossRef]
- Rosenfeld, A.B.; Doobin, D.J.; Warren, A.L.; Racaniello, V.R.; Vallee, R.B. Replication of Early and Recent Zika Virus Isolates throughout Mouse Brain Development. Proc. Natl. Acad. Sci. USA 2017, 114, 12273–12278. [Google Scholar] [CrossRef]
- Lathia, J.D.; Mack, S.C.; Mulkearns-Hubert, E.E.; Valentim, C.L.L.; Rich, J.N. Cancer Stem Cells in Glioblastoma. Genes Dev. 2015, 29, 1203–1217. [Google Scholar] [CrossRef]
- Zwernik, S.D.; Adams, B.H.; Raymond, D.A.; Warner, C.M.; Kassam, A.B.; Rovin, R.A.; Akhtar, P. AXL Receptor Is Required for Zika Virus Strain MR-766 Infection in Human Glioblastoma Cell Lines. Mol. Ther. Oncolytics 2021, 23, 447–457. [Google Scholar] [CrossRef]
- Zhu, Z.; Mesci, P.; Bernatchez, J.A.; Gimple, R.C.; Wang, X.; Schafer, S.T.; Wettersten, H.I.; Beck, S.; Clark, A.E.; Wu, Q.; et al. Zika Virus Targets Glioblastoma Stem Cells through a SOX2-Integrin Avβ5 Axis. Cell Stem Cell 2020, 26, 187–204.e10. [Google Scholar] [CrossRef]
- Chavali, P.L.; Stojic, L.; Meredith, L.W.; Joseph, N.; Nahorski, M.S.; Sanford, T.J.; Sweeney, T.R.; Krishna, B.A.; Hosmillo, M.; Firth, A.E.; et al. Neurodevelopmental Protein Musashi-1 Interacts with the Zika Genome and Promotes Viral Replication. Science 2017, 357, 83–88. [Google Scholar] [CrossRef]
- Kao, Y.T.; Wang, H.I.; Shie, C.T.; Lin, C.F.; Lai, M.M.C.; Yu, C.Y. Zika Virus Cleaves GSDMD to Disseminate Prognosticable and Controllable Oncolysis in a Human Glioblastoma Cell Model. Mol. Ther. Oncolytics 2023, 28, 104–117. [Google Scholar] [CrossRef]
- Nair, S.; Mazzoccoli, L.; Jash, A.; Govero, J.; Bais, S.S.; Hu, T.; Fontes-Garfias, C.R.; Shan, C.; Okada, H.; Shresta, S.; et al. Zika Virus Oncolytic Activity Requires CD8+ T Cells and Is Boosted by Immune Checkpoint Blockade. JCI Insight 2021, 6, e144619. [Google Scholar] [CrossRef] [PubMed]
- Crane, A.T.; Chrostek, M.R.; Krishna, V.D.; Shiao, M.; Toman, N.G.; Pearce, C.M.; Tran, S.K.; Sipe, C.J.; Guo, W.; Voth, J.P.; et al. Zika Virus-Based Immunotherapy Enhances Long-Term Survival of Rodents with Brain Tumors through Upregulation of Memory T-Cells. PLoS ONE 2020, 15, e0232858. [Google Scholar] [CrossRef] [PubMed]
- Iannolo, G.; Sciuto, M.R.; Cuscino, N.; Pallini, R.; Douradinha, B.; Vitiani, L.R.; De Maria, R.; Conaldi, P.G. Zika virus infection induces MiR34c expression in glioblastoma stem cells: New perspectives for brain tumor treatments. Cell Death Dis 2019, 10, 263. [Google Scholar] [CrossRef]
- Mazar, J.; Brooks, J.K.; Peloquin, M.; Rosario, R.; Sutton, E.; Longo, M.; Drehner, D.; Westmoreland, T.J. The Oncolytic Activity of Zika Viral Therapy in Human Neuroblastoma In Vivo Models Confers a Major Survival Advantage in a CD24-Dependent Manner. Cancer Res. Commun. 2024, 4, 65–80. [Google Scholar] [CrossRef]
- Garcez, P.P.; Guasti, A.; Ventura, N.; Higa, L.M.; Andreiuolo, F.; de Freitas, G.P.A.; Ribeiro, L.D.J.; Maia, R.A.; de Lima, S.M.B.; de Souza Azevedo, A.; et al. Case report: Regression of Glioblastoma after flavivirus infection. Front. Med. 2023, 10, 1192070. [Google Scholar] [CrossRef] [PubMed]
- Aid, M.; Abbink, P.; Larocca, R.A.; Boyd, M.; Nityanandam, R.; Nanayakkara, O.; Martinot, A.J.; Moseley, E.T.; Blass, E.; Borducchi, E.N.; et al. Zika Virus Persistence in the Central Nervous System and Lymph Nodes of Rhesus Monkeys. Cell 2017, 169, 610–620.e14. [Google Scholar] [CrossRef] [PubMed]
- Limonta, D.; Jovel, J.; Kumar, A.; Airo, A.M.; Hou, S.; Saito, L.; Branton, W.; Wong, G.K.S.; Mason, A.; Power, C.; et al. Human Fetal Astrocytes Infected with Zika Virus Exhibit Delayed Apoptosis and Resistance to Interferon: Implications for Persistence. Viruses 2018, 10, 646. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wu, J.; Ye, Q.; Ma, F.; Zhu, Q.; Wu, Y.; Shan, C.; Xie, X.; Li, D.; Zhan, X.; et al. Treatment of Human Glioblastoma with a Live Attenuated Zika Virus Vaccine Candidate. MBio 2018, 9, e01683-18. [Google Scholar] [CrossRef] [PubMed]
- Kaid, C.; dos Santos Madi, R.A.; Astray, R.; Goulart, E.; Caires-Junior, L.C.; Mitsugi, T.G.; Moreno, A.C.R.; Castro-Amarante, M.F.; Pereira, L.R.; Porchia, B.F.M.M.; et al. Safety, Tumor Reduction, and Clinical Impact of Zika Virus Injection in Dogs with Advanced-Stage Brain Tumors. Mol. Ther. 2020, 28, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.O.; Granha, I.; Ferreira, R.S.; Bueno, H.D.S.; Okamoto, O.K.; Kaid, C.; Zatz, M. Effect of Serial Systemic and Intratumoral Injections of Oncolytic ZIKVBR in Mice Bearing Embryonal CNS Tumors. Viruses 2021, 13, 2103. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Evaluation of Two Zika Viruses for Use in Controlled Human Infection Models (CHIM). Available online: https://classic.clinicaltrials.gov/ct2/show/NCT05123222 (accessed on 28 January 2024).



{kind=link}
{kind=link}
| OV | Type | Description | Phase | Type of CNS Tumor | CLT ID and/or References | |
|---|---|---|---|---|---|---|
| AdV | Ad-TD-nsIL12 | Eng | Deletion of E1ACR2, E1B19K and E3gp19K, WT E3B; armed with nsIL12, signal peptide deleted | I | Primary and progressive pediatric diffuse intrinsic pontine glioma | NCT05717712; NCT05717699 [40,41,42] |
| DNX-2401 (also known as tasadenoturev; Delta-24-RGD) | Eng | 24 bp deletion in E1A; RGD peptide insertion into the fiber knob that allows the virus to anchor directly to integrins | I, II | Naive diffuse intrinsic pontine gliomas | NCT03178032; NCT01956734; NCT02197169 [43] | |
| ADV-TK | Eng | Vector contains the TK gene | I | Recurrent high-grade glioma | NCT00870181 [44] | |
| DNX-2440 | Eng | 24 bp deletion in E1A; insertion of OX40L and RGD-4C genes | I | First or second recurrence of GBM | NCT03714334 | |
| HSV-1 | G207 | Eng | Deletion of γ134.5 gene, insertion of lacZ operon in UL39 | I, II | Recurrent or refractory cerebellar brain tumors; progressive or recurrent supratentorial brain tumors; recurrent high-grade glioma; malignant glioma | NCT03911388; NCT02457845; NCT04482933; NCT00157703; NCT00028158 [45] |
| MVR-C5252 | Eng | Active domain of human IL-12 and Fab fragment of anti-PD-1 antibody | I | Recurrent high-grade glioma | NCT06126744 [46] | |
| rQNestin (also known as rQNestin34.5v.2) | Eng | Restoration of one copy of ICP34.5 under transcriptional control of NP/EE | I | Malignant glioma | NCT03152318 [47] | |
| M032 | Eng | a γ134.5-deleted HSV-1 engineered to express murine IL-12 | I | Recurrent malignant glioma | NCT02062827 [48] | |
| Poliovirus | PVS-RIPO (also known as Lerapolturev) | Eng | Recombinant, non-pathogenic poliovirus:rhinovirus chimera; genome of PV1S with its cognate IRES element replaced with that of HRV2 | I, II | Malignant glioma (primary and recurrent); GBM | NCT02986178; NCT03043391; NCT01491893; NCT04479241 |
| REO | Reolysin | WT | Strain of a type 3 REO that selectively infects and lyses tumor cells via the Ras-activated pathway | I, II | Recurrent malignant gliomas; with high-grade relapsed or refractory brain tumors | NCT00528684; NCT02444546 |
| Retrovirus | Toca511 (also known as Vocimagene amiretrorepvec) | Eng | Non-lytic RRV that delivers a yeast cytosine deaminase to convert the prodrug Toca FC into the antimetabolite 5-fluorouracil | I, II, III | Recurrent brain tumors (anaplastic astrocytoma; anaplastic oligoastrocytoma; anaplastic oligodendroglioma; GBM) | NCT01985256; NCT01470794; NCT01156584 |
| MV | MV-CEA | Eng | CEA incorporated in the vector | I | Recurrent GBM | NCT00390299 |
| Vaccinia virus | TG6002 | Eng | Vaccinia virus strain Copenhagen with deletion of TK and RR genes, expresses the FCU1 gene | I | Recurrent GBM | NCT03294486 [49] |
| Parvovirus | ParvOryx | WT | WT rat parvovirus H1 (H-1PV) | I, II | Progressive primary or recurrent GBM | NCT01301430 [50] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderón-Peláez, M.-A.; Maradei Anaya, S.J.; Bedoya-Rodríguez, I.J.; González-Ipuz, K.G.; Vera-Palacios, D.; Buitrago, I.V.; Castellanos, J.E.; Velandia-Romero, M.L. Zika Virus: A Neurotropic Warrior against High-Grade Gliomas—Unveiling Its Potential for Oncolytic Virotherapy. Viruses 2024, 16, 561. https://doi.org/10.3390/v16040561
Calderón-Peláez M-A, Maradei Anaya SJ, Bedoya-Rodríguez IJ, González-Ipuz KG, Vera-Palacios D, Buitrago IV, Castellanos JE, Velandia-Romero ML. Zika Virus: A Neurotropic Warrior against High-Grade Gliomas—Unveiling Its Potential for Oncolytic Virotherapy. Viruses. 2024; 16(4):561. https://doi.org/10.3390/v16040561
Chicago/Turabian StyleCalderón-Peláez, María-Angélica, Silvia Juliana Maradei Anaya, Ingrid Juliana Bedoya-Rodríguez, Karol Gabriela González-Ipuz, Daniela Vera-Palacios, Isabella Victoria Buitrago, Jaime E. Castellanos, and Myriam L. Velandia-Romero. 2024. "Zika Virus: A Neurotropic Warrior against High-Grade Gliomas—Unveiling Its Potential for Oncolytic Virotherapy" Viruses 16, no. 4: 561. https://doi.org/10.3390/v16040561