
First Detection and Circulation of RHDV2 in New Zealand
 , , ,
, , ,
Abstract
:1. Introduction

2. Materials and Methods
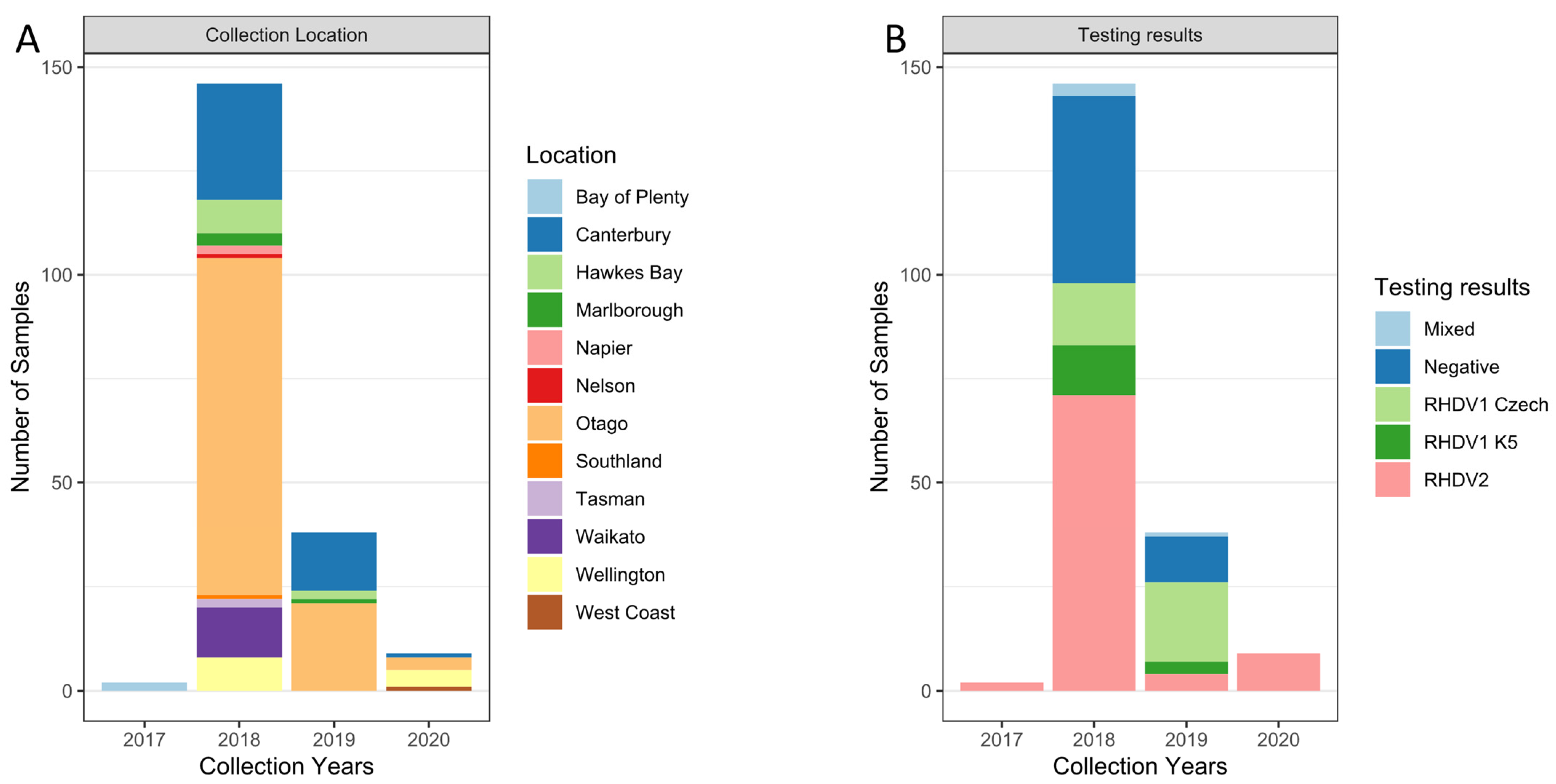
2.1. Tissue Sample Collection
2.2. Virus Detection
2.3. Virus Sequencing
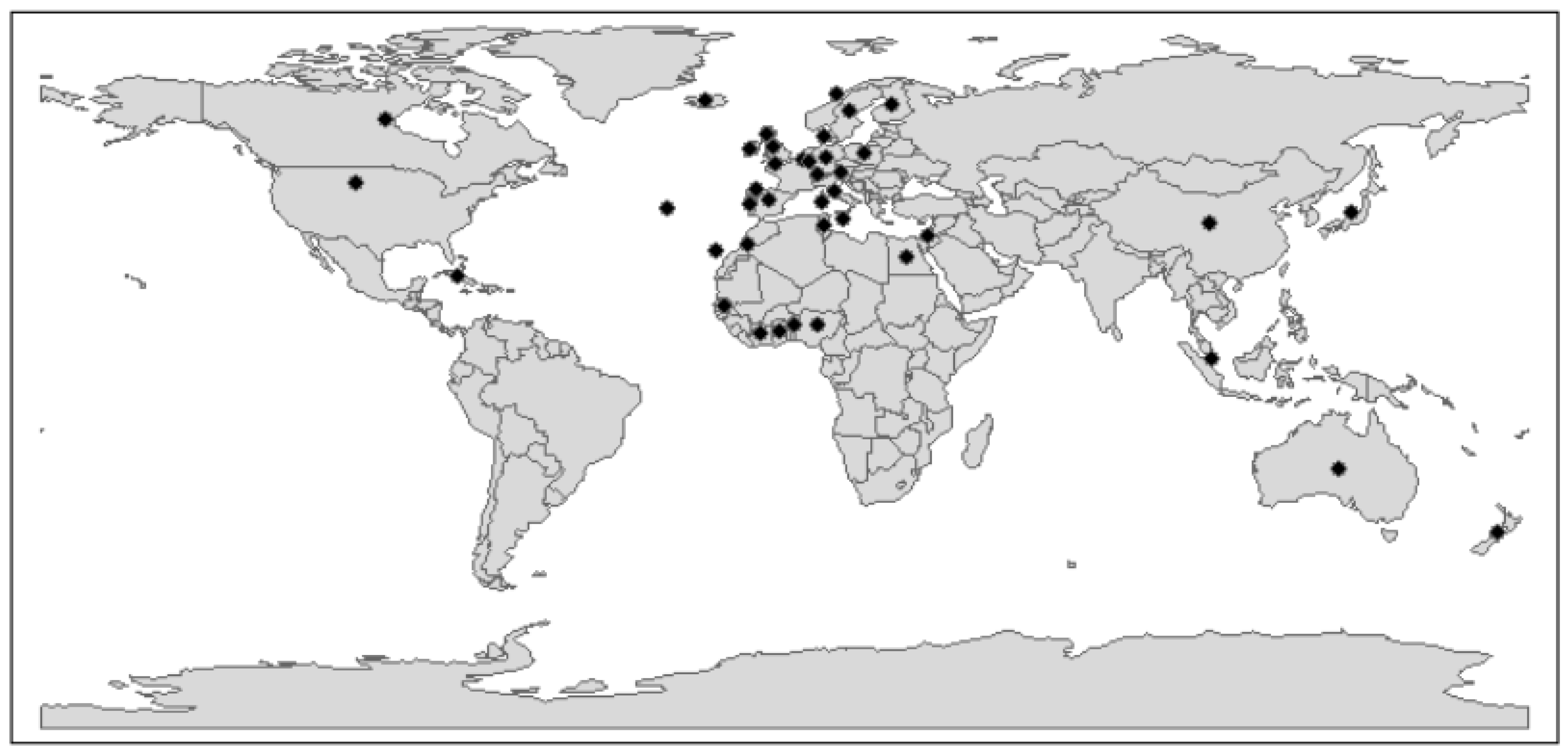
2.4. Phylogenetic and Phylogeographic Analyses
2.5. Bayesian Evolutionary Analysis
2.6. Figures
3. Results
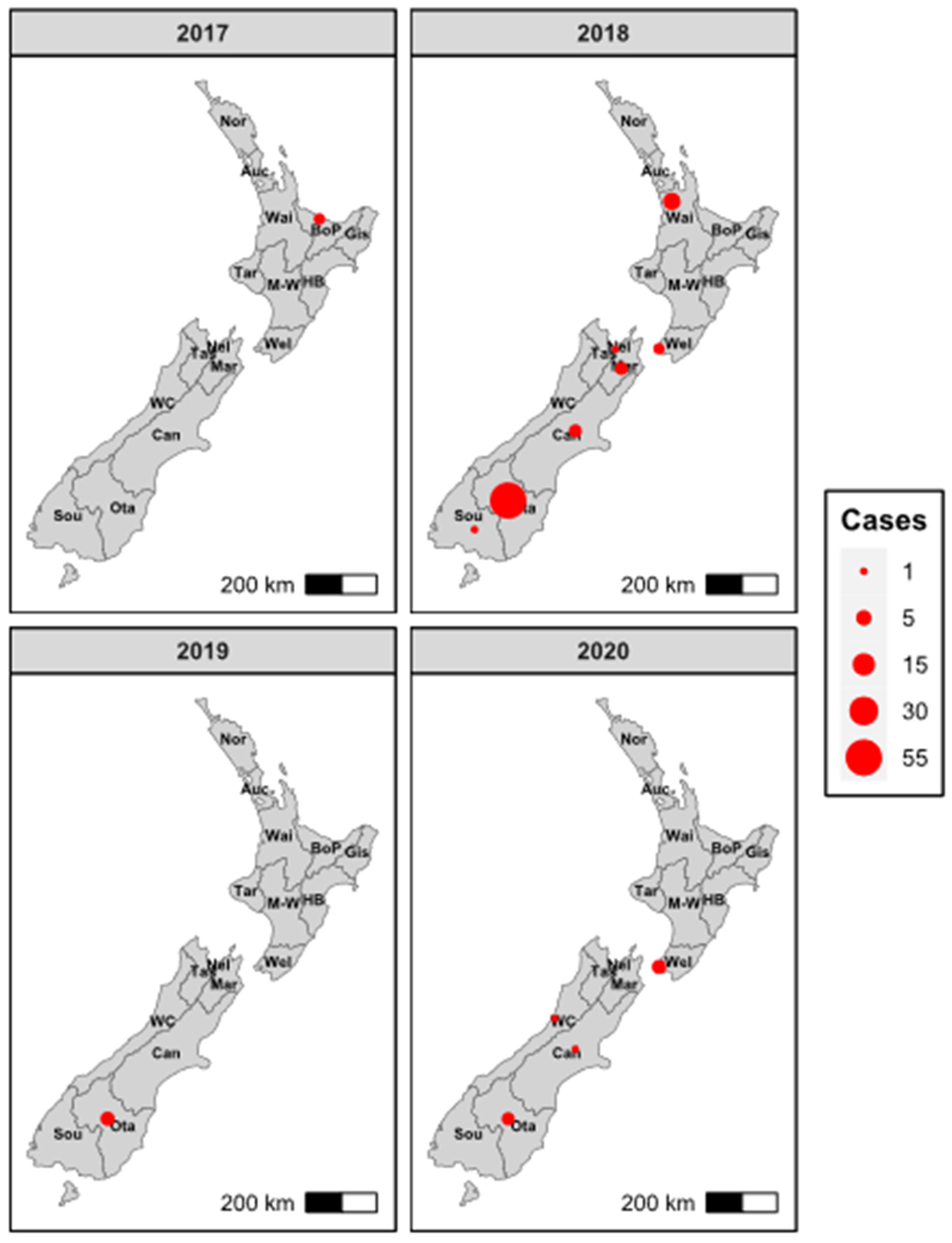
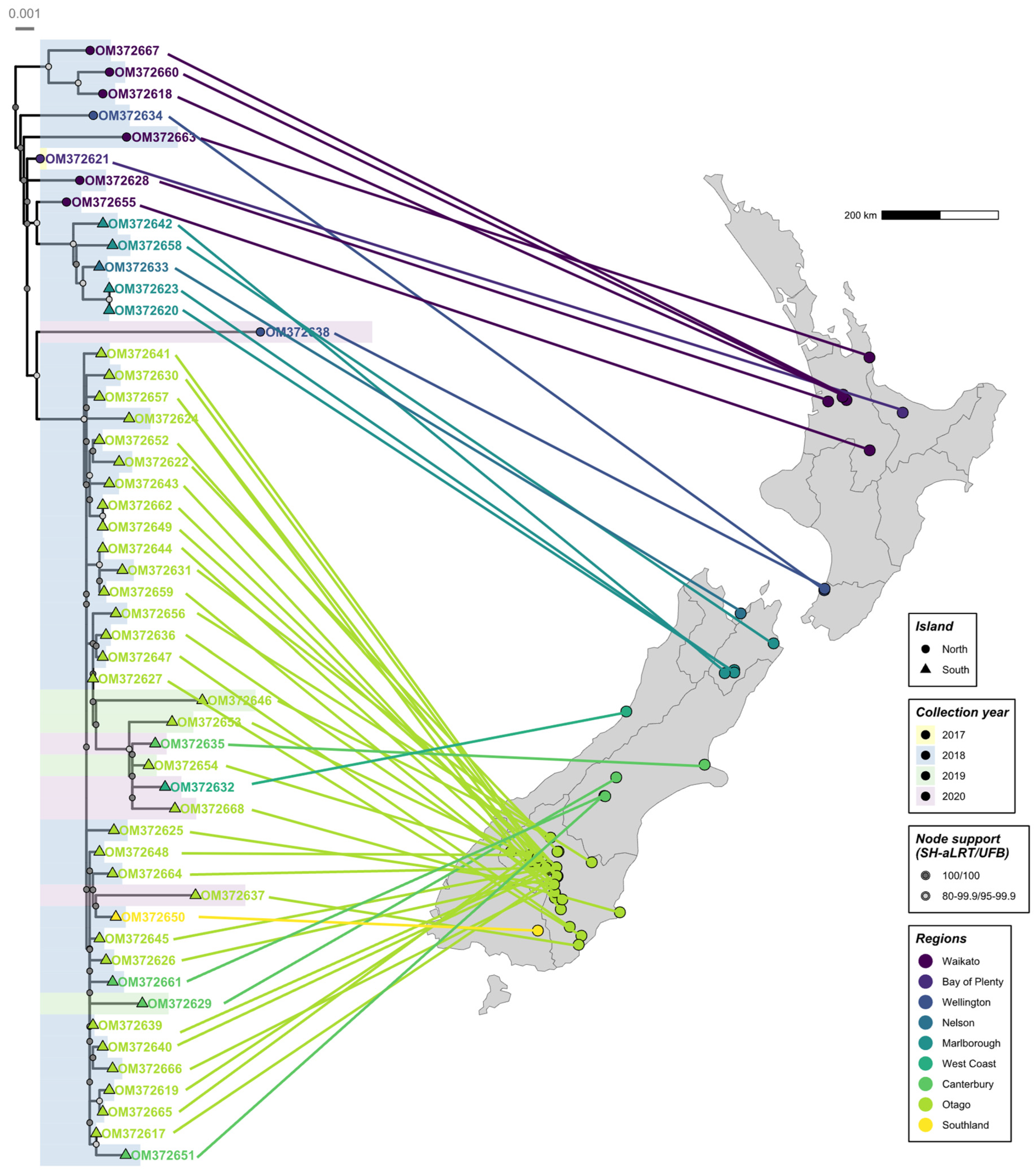
3.1. RHDV2 Was First Detected in New Zealand in 2017
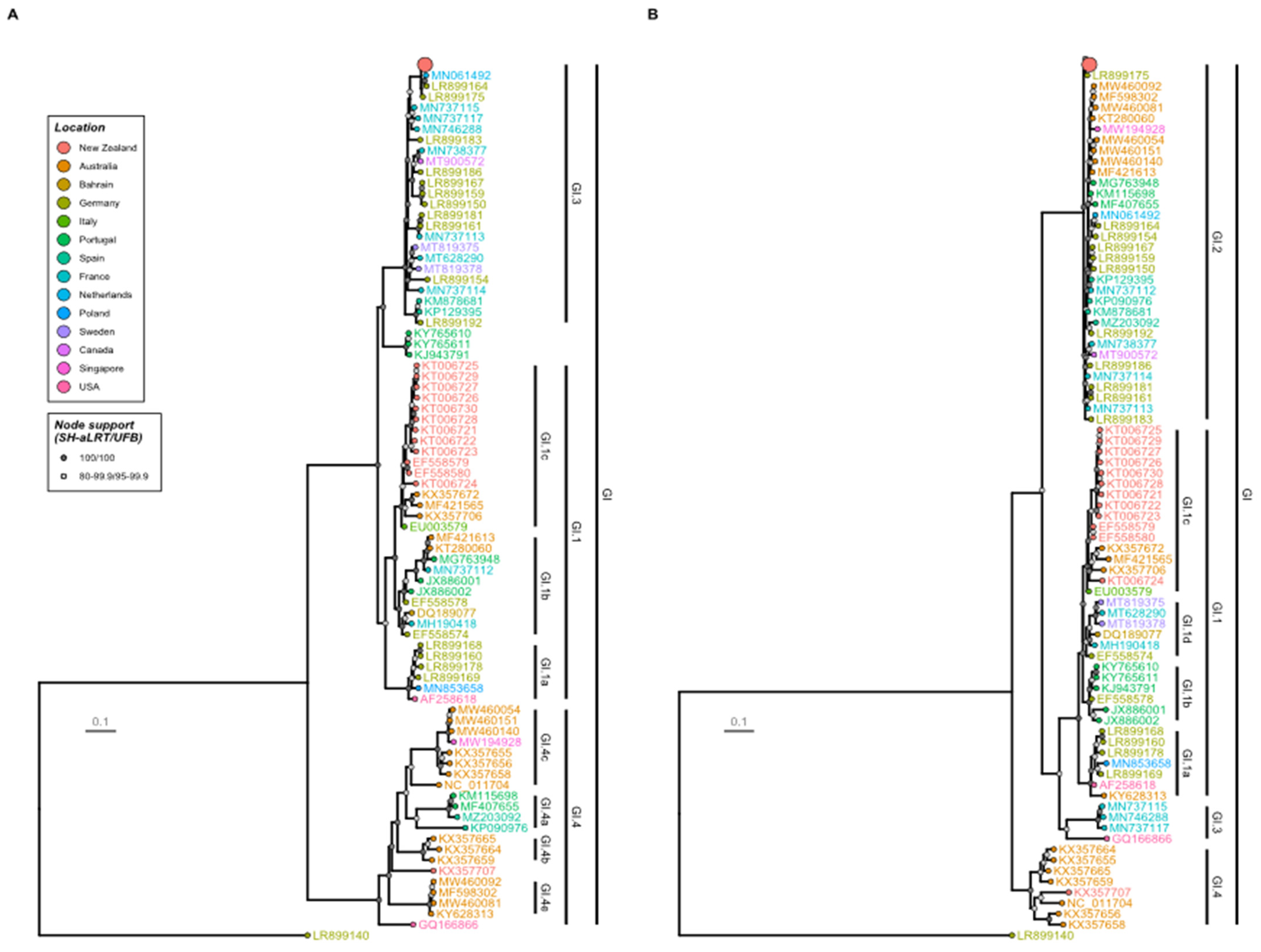
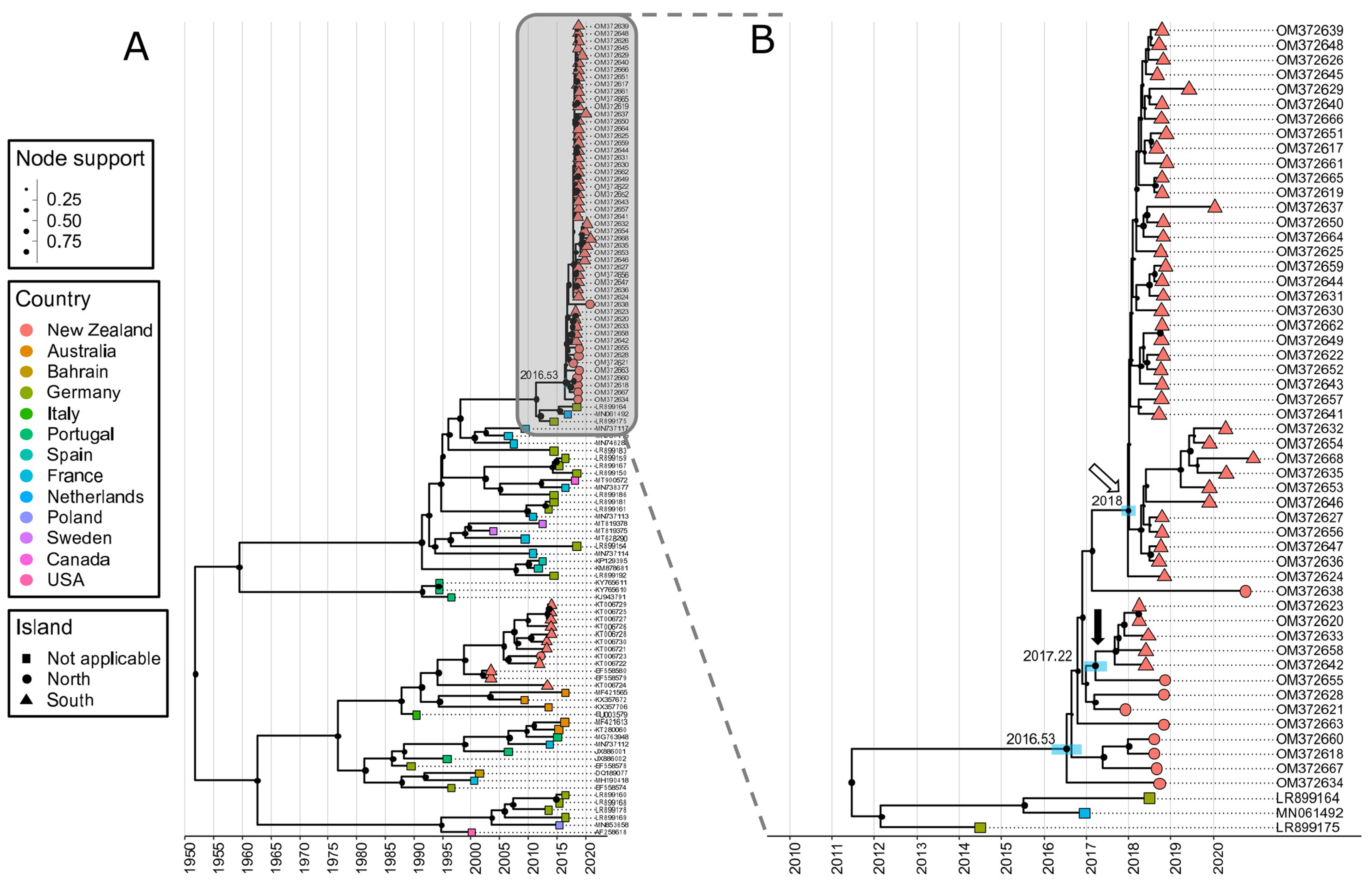
3.2. Phylogenetic Analysis Reveals the New Zealand RHDV2 Incursion as Genotype GI.3P-GI.2
3.3. First Introduction to New Zealand in Mid-2016
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Recule, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Celio, A.P.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Le Gall-Recule, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guerin, J.L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit haemorrhagic disease virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Nicieza, I.; Balseiro, A.; Muguerza, M.A.; Rosell, J.M.; Casais, R.; Alvarez, A.L.; Parra, F. Variant Rabbit hemorrhagic disease virus in young rabbits, Spain. Emerg. Infect. Dis. 2012, 18, 2009–2012. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.N.; King, T.; O’Connor, T.; Read, A.J.; Arrow, J.; Trought, K.; Duckworth, J.; Piper, M.; Strive, T. Age and Infectious Dose Significantly Affect Disease Progression after RHDV2 Infection in Naive Domestic Rabbits. Viruses 2021, 13, 1184. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Nicieza, I.; Abrantes, J.; Esteves, P.J.; Parra, F. Spread of new variant RHDV in domestic rabbits on the Iberian Peninsula. Vet. Microbiol. 2014, 169, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Buehler, M.; Jesse, S.T.; Kueck, H.; Lange, B.; Koenig, P.; Jo, W.K.; Osterhaus, A.; Beineke, A. Lagovirus europeus GI.2 (Rabbit hemorrhagic disease virus 2) infection in captive mountain hares (Lepus timidus) in Germany. Vet. Res. 2020, 16, 166. [Google Scholar] [CrossRef]
- Camarda, A.; Pugliese, N.; Cavadini, P.; Circella, E.; Capucci, L.; Caroli, A.; Legretto, M.; Mallia, E.; Lavazza, A. Detection of the new emerging Rabbit haemorrhagic disease type 2 virus (RHDV2) in Sicily from rabbit (Oryctolagus cuniculus) and Italian hare (Lepus corsicanus). Res. Vet. Sci. 2014, 97, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Cavadini, P.; Molinari, S.; Merzoni, F.; Vismarra, A.; Posautz, A.; Alzaga Gil, V.; Chiari, M.; Giannini, F.; Capucci, L.; Lavazza, A. Widespread occurrence of the non-pathogenic hare calicivirus (HaCV Lagovirus GII.2) in captive-reared and free-living wild hares in Europe. Transbound. Emerg. Dis. 2021, 68, 509–518. [Google Scholar] [CrossRef]
- Lankton, J.S.; Knowles, S.; Keller, S.; Shearn-Bochsler, V.I.; Ip, H.S. Pathology of Lagovirus europaeus GI.2/RHDV2/b (Rabbit hemorrhagic disease virus 2) in native North American lagomorphs. J. Wildl. Dis. 2021, 57, 694–700. [Google Scholar] [CrossRef]
- Le Gall-Recule, G.; Lemaitre, E.; Bertagnoli, S.; Hubert, C.; Top, S.; Decors, A.; Marchandeau, S.; Guitton, J.S. Large-scale lagovirus disease outbreaks in European brown hares (Lepus europaeus) in France caused by RHDV2 strains spatially shared with rabbits (Oryctolagus cuniculus). Vet. Res. 2017, 48, 70. [Google Scholar] [CrossRef]
- Neimanis, A.S.; Ahola, H.; Larsson Pettersson, U.; Lopes, A.M.; Abrantes, J.; Zohari, S.; Esteves, P.J.; Gavier-Widen, D. Overcoming species barriers: An outbreak of Lagovirus europaeus GI.2/RHDV2 in an isolated population of mountain hares (Lepus timidus). Vet. Res. 2018, 14, 367. [Google Scholar] [CrossRef] [PubMed]
- Puggioni, G.; Cavadini, P.; Maestrale, C.; Scivoli, R.; Botti, G.; Ligios, C.; Le Gall-Recule, G.; Lavazza, A.; Capucci, L. The new French 2010 Rabbit hemorrhagic disease virus causes an RHD-like disease in the Sardinian Cape hare (Lepus capensis mediterraneus). Vet. Res. 2013, 44, 96. [Google Scholar] [CrossRef] [PubMed]
- Velarde, R.; Cavadini, P.; Neimanis, A.; Cabezon, O.; Chiari, M.; Gaffuri, A.; Lavin, S.; Grilli, G.; Gavier-Widen, D.; Lavazza, A.; et al. Spillover events of infection of brown hares (Lepus europaeus) with Rabbit haemorrhagic disease type 2 virus (RHDV2) caused sporadic cases of an European brown hare syndrome-like disease in Italy and Spain. Transbound. Emerg. Dis. 2017, 64, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.N.; Peacock, D.E.; Kovaliski, J.; Mahar, J.E.; Mourant, R.; Piper, M.; Strive, T. Detection of RHDV2 in European brown hares (Lepus europaeus) in Australia. Vet. Rec. 2017, 180, 121. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, T.W.; Read, A.J.; Hall, R.N.; Strive, T.; Kirkland, P.D. Immunological Cross-Protection between Different Rabbit Hemorrhagic Disease Viruses-Implications for Rabbit Biocontrol and Vaccine Development. Vaccines 2022, 10, 666. [Google Scholar] [CrossRef] [PubMed]
- Taggart, P.L.; Hall, R.N.; Cox, T.E.; Kovaliski, J.; McLeod, S.R.; Strive, T. Changes in virus transmission dynamics following the emergence of RHDV2 shed light on its competitive advantage over previously circulating variants. Transbound. Emerg. Dis. 2022, 69, 1118–1130. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.V.; Matson, D.O.; Smith, A.W. Three-dimensional structure of calicivirus. J. Mol. Biol. 1994, 240, 256–264. [Google Scholar] [CrossRef]
- Prasad, B.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray crystallographic structure of the Norwalk virus capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Parra, F.; Prieto, M. Purification and characterization of a calicivirus as the causative agent of a lethal hemorrhagic disease in rabbits. J. Virol. 1990, 64, 4013–4015. [Google Scholar] [CrossRef]
- Ehresmann, D.W.; Schaffer, F.L. RNA synthesized in calicivirus-infected cells is atypical of picornaviruses. J. Virol. 1977, 22, 572–576. [Google Scholar] [CrossRef]
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Genomic and subgenomic RNAs of rabbit hemorrhagic disease virus are both protein-linked and packaged into particles. Virology 1991, 184, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Rabbit hemorrhagic disease virus—Molecular cloning and nucleotide sequencing of a calicivirus genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.J.; Thumfart, J.O. Rabbit hemorrhagic disease virus: Genome organization and polyprotein processing of a calicivirus studied after transient expression of cDNA constructs. Virology 2000, 276, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Boga, J.A.; Marín, M.S.; Casais, R.; Prieto, M.; Parra, F. In vitro translation of a subgenomic mRNA from purified virions of the Spanish field isolate AST/89 of rabbit hemorrhagic disease virus (RHDV). Virus Res. 1992, 26, 33–40. [Google Scholar] [CrossRef]
- Black, D.N.; Burroughs, J.N.; Harris, T.J.; Brown, F. The structure and replication of calicivirus RNA. Nature 1978, 274, 614–615. [Google Scholar] [CrossRef]
- Burroughs, J.N.; Brown, F. Presence of a covalently linked protein on calicivirus RNA. J. Gen. Virol. 1978, 41, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Mahar, J.E.; Jenckel, M.; Huang, N.; Smertina, E.; Holmes, E.C.; Strive, T.; Hall, R.N. Frequent intergenotypic recombination between the non-structural and structural genes is a major driver of epidemiological fitness in caliciviruses. Virus Evol. 2021, 7, veab080. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Recule, G. Recombination at the emergence of the pathogenic rabbit haemorrhagic disease virus Lagovirus europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Hall, R.N.; Mahar, J.E.; Haboury, S.; Stevens, V.; Holmes, E.C.; Strive, T. Emerging Rabbit Hemorrhagic Disease Virus 2 (RHDVb), Australia. Emerg. Infect. Dis. 2015, 21, 2276–2278. [Google Scholar] [CrossRef]
- Almeida, T.; Lopes, A.M.; Magalhaes, M.J.; Neves, F.; Pinheiro, A.; Goncalves, D.; Leitao, M.; Esteves, P.J.; Abrantes, J. Tracking the evolution of the G1/RHDVb recombinant strains introduced from the Iberian Peninsula to the Azores islands, Portugal. Infect. Genet. Evol. 2015, 34, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Correia, J.; Abrantes, J.; Melo, P.; Ramada, M.; Magalhaes, M.J.; Alves, P.C.; Esteves, P.J. Is the new variant RHDV replacing genogroup 1 in Portuguese wild rabbit populations? Viruses 2014, 7, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.N.; Mahar, J.E.; Read, A.J.; Mourant, R.; Piper, M.; Huang, N.; Strive, T. A strain-specific multiplex RT-PCR for Australian rabbit haemorrhagic disease viruses uncovers a new recombinant virus variant in rabbits and hares. Transbound. Emerg. Dis. 2018, 65, e444–e456. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Abrantes, J.; Lopes, A.M.; Nicieza, I.; Alvarez, A.L.; Esteves, P.J.; Parra, F. Complete genome sequence of two rabbit hemorrhagic disease virus variant b isolates detected on the Iberian Peninsula. Arch. Virol. 2015, 160, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Dalton, K.P.; Magalhaes, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full genomic analysis of new variant rabbit hemorrhagic disease virus revealed multiple recombination events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef]
- Lopes, A.M.; Blanco-Aguiar, J.; Martin-Alonso, A.; Leitao, M.; Foronda, P.; Mendes, M.; Goncalves, D.; Abrantes, J.; Esteves, P.J. Full genome sequences are key to disclose RHDV2 emergence in the Macaronesian islands. Virus Genes 2018, 54, 1–4. [Google Scholar] [CrossRef]
- Silverio, D.; Lopes, A.M.; Melo-Ferreira, J.; Magalhaes, M.J.; Monterroso, P.; Serronha, A.; Maio, E.; Alves, P.C.; Esteves, P.J.; Abrantes, J. Insights into the evolution of the new variant rabbit haemorrhagic disease virus (GI.2) and the identification of novel recombinant strains. Transbound. Emerg. Dis. 2018, 65, 983–992. [Google Scholar] [CrossRef]
- Miao, Q.; Qi, R.; Veldkamp, L.; Ijzer, J.; Kik, M.L.; Zhu, J.; Tang, A.; Dong, D.; Shi, Y.; van Oers, M.M.; et al. Immunogenicity in Rabbits of Virus-Like Particles from a Contemporary Rabbit Haemorrhagic Disease Virus Type 2 (GI.2/RHDV2/b) Isolated in The Netherlands. Viruses 2019, 11, 553. [Google Scholar] [CrossRef]
- Szillat, K.P.; Höper, D.; Beer, M.; Konig, P. Full-genome sequencing of German rabbit haemorrhagic disease virus uncovers recombination between RHDV (GI.2) and EBHSV (GII.1). Virus Evol. 2020, 6, veaa080. [Google Scholar] [CrossRef]
- Fitzner, A.; Kesy, A.; Bulenger, K.; Niedbalski, W. Evidence of independent introductions of RHDV2 strains in Poland based on the genome analysis of viral isolates from 2016–2018. Acta Biochim. Pol. 2021, 68, 255–263. [Google Scholar] [CrossRef]
- Hu, B.; Wei, H.; Fan, Z.; Song, Y.; Chen, M.; Qiu, R.; Zhu, W.; Xu, W.; Xue, J.; Wang, F. Emergence of rabbit haemorrhagic disease virus 2 in China in 2020. Vet. Med. Sci. 2021, 7, 236–239. [Google Scholar] [CrossRef]
- Ambagala, A.; Schwantje, H.; Laurendeau, S.; Snyman, H.; Joseph, T.; Pickering, B.; Hooper-McGrevy, K.; Babiuk, S.; Moffat, E.; Lamboo, L.; et al. Incursions of rabbit haemorrhagic disease virus 2 in Canada-Clinical, molecular and epidemiological investigation. Transbound. Emerg. Dis. 2021, 68, 1711–1720. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, V.K.; Xu, L.; Moran, K.; Mohamed, F.; Boston, T.; Pauszek, S.J.; Vierra, D.A.; Faburay, B.; Dodd, K.A.; Barrette, R.W. Coding-Complete Genome Sequences of Emerging Rabbit Hemorrhagic Disease Virus Type 2 Isolates Detected in 2020 in the United States. Microbiol. Resour. Announc. 2021, 10, e01064-20. [Google Scholar] [CrossRef] [PubMed]
- Rouco, C.; Aguayo-Adan, J.A.; Santoro, S.; Abrantes, J.; Delibes-Mateos, M. Worldwide rapid spread of the novel rabbit haemorrhagic disease virus (GI.2/RHDV2/b). Transbound. Emerg. Dis. 2019, 66, 1762–1764. [Google Scholar] [CrossRef] [PubMed]
- Cooke, B.; Chudleigh, P.; Simpson, S.; Saunders, G. The Economic Benefits of the Biological Control of Rabbits in Australia, 1950–2011. Aust. Econ. Hist. Rev. 2013, 53, 91–107. [Google Scholar] [CrossRef]
- NZERMA. Evaluation and Review Report: Reassessment of 1080 (HRE05002). Appendix G: Prevalence, Transmission and Control of Bovine Tuberculosis. 2007. Available online: https://www.epa.govt.nz/assets/FileAPI/hsno-ar/HRE05002/83dfaf6d4d/HRE05002-050.pdf (accessed on 27 February 2024).
- Cruz, J.; Glen, A.S.; Pech, R.P. Modelling Landscape-Level Numerical Responses of Predators to Prey: The Case of Cats and Rabbits. PLoS ONE 2013, 8, e73544. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.S.; Tempero, J.E.; Motha, M.X.; Hansen, M.F.; Atkinsona, P.H. Serology of rabbit haemorrhagic disease virus in wild rabbits before and after release of the virus in New Zealand. Vet. Microbiol. 1999, 66, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Sanson, R.L.; Brooks, H.V.; Horner, G.W. An epidemiological study of the spread of rabbit haemorrhagic disease virus amongst previously non-exposed rabbit populations in the North Island of New Zealand. N. Z. Vet. J. 2000, 48, 105–110. [Google Scholar] [CrossRef] [PubMed]
- SPCA. Rabbit Haemorrhagic Disease Virus and Rabbit Vaccination in New Zealand. 2018. Available online: https://www.spca.nz/advice-and-welfare/article/rabbit-haemorrhagic-disease-virus-and-rabbit-vaccination-in-new-zealand (accessed on 24 September 2021).
- Nicholson, L.J.; Mahar, J.E.; Strive, T.; Zheng, T.; Holmes, E.C.; Ward, V.K.; Duckworth, J.A. Benign Rabbit Calicivirus in New Zealand. Appl. Environ. Microbiol. 2017, 83, e00090-17. [Google Scholar] [CrossRef]
- National Animal Ethics Advisory Committee. Regulation of Animal Use in Research, Testing and Teaching in New Zealand—The Black, the White and the Grey. 2009. Available online: https://www.naeac.org.nz/assets/NAEAC-documents/Occasional-paper-2.pdf (accessed on 12 February 2024).
- Eden, J.S.; Kovaliski, J.; Duckworth, J.A.; Swain, G.; Mahar, J.E.; Strive, T.; Holmes, E.C. Comparative Phylodynamics of Rabbit Hemorrhagic Disease Virus in Australia and New Zealand. J. Virol. 2015, 89, 9548–9558. [Google Scholar] [CrossRef]
- Mahar, J.E.; Nicholson, L.; Eden, J.S.; Duchene, S.; Kerr, P.J.; Duckworth, J.; Ward, V.K.; Holmes, E.C.; Strive, T. Benign Rabbit Caliciviruses Exhibit Evolutionary Dynamics Similar to Those of Their Virulent Relatives. J. Virol. 2016, 90, 9317–9329. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.N.; Capucci, L.; Matthaei, M.; Esposito, S.; Kerr, P.J.; Frese, M.; Strive, T. An in vivo system for directed experimental evolution of rabbit haemorrhagic disease virus. PLoS ONE 2017, 12, e0173727. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Rambaut, A.; Lam, T.T.; Max Carvalho, L.; Pybus, O.G. Exploring the temporal structure of heterochronous sequences using TempEst (formerly Path-O-Gen). Virus Evol. 2016, 2, vew007. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Wickham, H.; Seidel, D. Scales: Scale Functions for Visualization, R package version 1.1.1; RStudio, Inc.: Vienna, Austria, 2020.
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; D’Agostino McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Bivand, R.; Nowosad, J.; Lovelace, R. spData: Datasets for Spatial Analysis, R package version 2.0.1; RStudio, Inc.: Vienna, Austria, 2021. [Google Scholar]
- Dunnington, D. ggspatial: Spatial Data Framework for ggplot2, R package version 1.1.5; RStudio, Inc.: Vienna, Austria, 2021.
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Wilke, C. cowplot: Streamlined Plot Theme and Plot Annotations for ‘ggplot2’, R package version 1.1.1; RStudio, Inc.: Vienna, Austria, 2020.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. dplyr: A Grammar of Data Manipulation, R Package Version 1.1.2; The R Foundation for Statistical Computing: Vienna, Austria, 2023.
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes; RStudio, Inc.: Vienna, Austria, 2022. [Google Scholar]
- Arnold, J.B. ggthemes: Extra Themes, Scales and Geoms for ‘ggplot2’; RStudio, Inc.: Vienna, Austria, 2021. [Google Scholar]
- Wang, L.G.; Lam, T.T.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R Package for Phylogenetic Tree Input and Output with Richly Annotated and Associated Data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Henning, J.; Meers, J.; Davies, P.R.; Morris, R.S. Survival of rabbit haemorrhagic disease virus (RHDV) in the environment. Epidemiol. Infect. 2005, 133, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.N.; Huang, N.; Roberts, J.; Strive, T. Carrion flies as sentinels for monitoring lagovirus activity in Australia. Transbound. Emerg. Dis. 2019, 66, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Simpson, V.; Everest, D.; Westcott, D. RHDV variant 2 and Capillaria hepatica infection in rabbits. Vet. Rec. 2014, 174, 486. [Google Scholar] [CrossRef]
- Martin-Alonso, A.; Martin-Carrillo, N.; Garcia-Livia, K.; Valladares, B.; Foronda, P. Emerging rabbit haemorrhagic disease virus 2 (RHDV2) at the gates of the African continent. Infect. Genet. Evol. 2016, 44, 46–50. [Google Scholar] [CrossRef]
- Carvalho, C.L.; Silva, S.; Gouveia, P.; Costa, M.; Duarte, E.L.; Henriques, A.M.; Barros, S.S.; Luis, T.; Ramos, F.; Fagulha, T.; et al. Emergence of rabbit haemorrhagic disease virus 2 in the archipelago of Madeira, Portugal (2016–2017). Virus Genes 2017, 53, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Peng, N.Y.G.; Hall, R.N.; Huang, N.; West, P.; Cox, T.E.; Mahar, J.E.; Mason, H.; Campbell, S.; O’Connor, T.; Read, A.J.; et al. Utilizing Molecular Epidemiology and Citizen Science for the Surveillance of Lagoviruses in Australia. Viruses 2023, 15, 2348. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, D.S.L.; Cox, T.; Strive, T.; Forsyth, D.M.; Stuart, I.; Hall, R.; Elsworth, P.; Campbell, S. Emerging RHDV2 suppresses the impact of endemic and novel strains of RHDV on wild rabbit populations. J. Appl. Ecol. 2020, 57, 630–641. [Google Scholar] [CrossRef]
- Mahar, J.E.; Hall, R.N.; Peacock, D.; Kovaliski, J.; Piper, M.; Mourant, R.; Huang, N.; Campbell, S.; Gu, X.; Read, A.; et al. Rabbit Hemorrhagic Disease Virus 2 (RHDV2; GI.2) Is Replacing Endemic Strains of RHDV in the Australian Landscape within 18 Months of Its Arrival. J. Virol. 2018, 92, e01374-17. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Amplicon Size (bp) | Nucleotide Position a | Gene b |
|---|---|---|---|---|
| GI.1a-Aus_fwd | GCGTGGCATTGTGCGCAGCATC | 562 | 4349 | RdRp |
| GI.1a-Aus_rev | TGTTGGTGATAAGCCATAATCGCG | 4911 | ||
| GI.1c_fwd | AGCAAGACTGTTGACTCAATTTCG | 435 | 5995 | VP60 |
| GI.1c_rev | AGGCCTGCACAGTCGTAACGTT | 6430 | ||
| GI.2_fwd | TTTCCCTGGAAGCAGTTCGTCA | 336 | 6213 | VP60 |
| GI.2_rev | TGTTGTCTGGTTTATGCCATTTGC | 6549 | ||
| GI.1a-K5_fwd | TTTATAGATGTATGCCCGCTCAAC | 263 | 4930 | RdRp |
| GI.1a-K5_rev | CCGTTCGAGTTCCTTGCGGACG | 5193 |
| Primer | Sequence (5′-3′) | Fragment | Ref |
|---|---|---|---|
| RHDV-1 | GTGAAARTTATGSCGGCTATGTCGCGC | 1 | [52] |
| GI.3PGI.2_1667R | TGGTCAAGGCCAAAGTTRATCG | This study | |
| RHDV1_GI.3 | GTGAAAGTTATGGCGGCTATGT | 1 alternate | This study |
| GI.3PGI.2_1698R | AGCCACTTCCTCTCCTGTGTAT | This study | |
| GI.3PGI.2_1451F | ACAACCAACACCTYGGAATCTTGAATG | 2 | This study |
| GI.3PGI.2_3064R | CTTGCCACTCRTCGTACTCRTC | This study | |
| GI.3PGI.2_2918F | TGTGYGCAAACCACCTTGTYAA | 3 | This study |
| GI.3PGI.2_4607R | CTCTTGGTGAGTTCCGTCTGYTCA | This study | |
| GI.3PGI.2_4399F | AAGATGGTCGCACGGTTTG | 4 | This study |
| GI.3PGI.2_6030R | GAGATCTGCGGGCGAGAT | This study | |
| GI.3PGI.2_5867F | CCACGTTGGTCCTGAGCGTYTA | 5 | This study |
| RHDV-end | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTATAATTTACTCTAAATTATAAACCAATTAAATTAATTAAC | [54] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, R.N.; Trought, K.; Strive, T.; Duckworth, J.A.; Jenckel, M. First Detection and Circulation of RHDV2 in New Zealand. Viruses 2024, 16, 519. https://doi.org/10.3390/v16040519
Hall RN, Trought K, Strive T, Duckworth JA, Jenckel M. First Detection and Circulation of RHDV2 in New Zealand. Viruses. 2024; 16(4):519. https://doi.org/10.3390/v16040519
Chicago/Turabian StyleHall, Robyn N., Katherine Trought, Tanja Strive, Janine A. Duckworth, and Maria Jenckel. 2024. "First Detection and Circulation of RHDV2 in New Zealand" Viruses 16, no. 4: 519. https://doi.org/10.3390/v16040519