
Neglected Spleen Transcriptional Profile Reveals Inflammatory Disorder Conferred by Rabbit Hemorrhagic Disease Virus 2 Infection
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Animals
2.2. Infection and Collection of Biological Samples
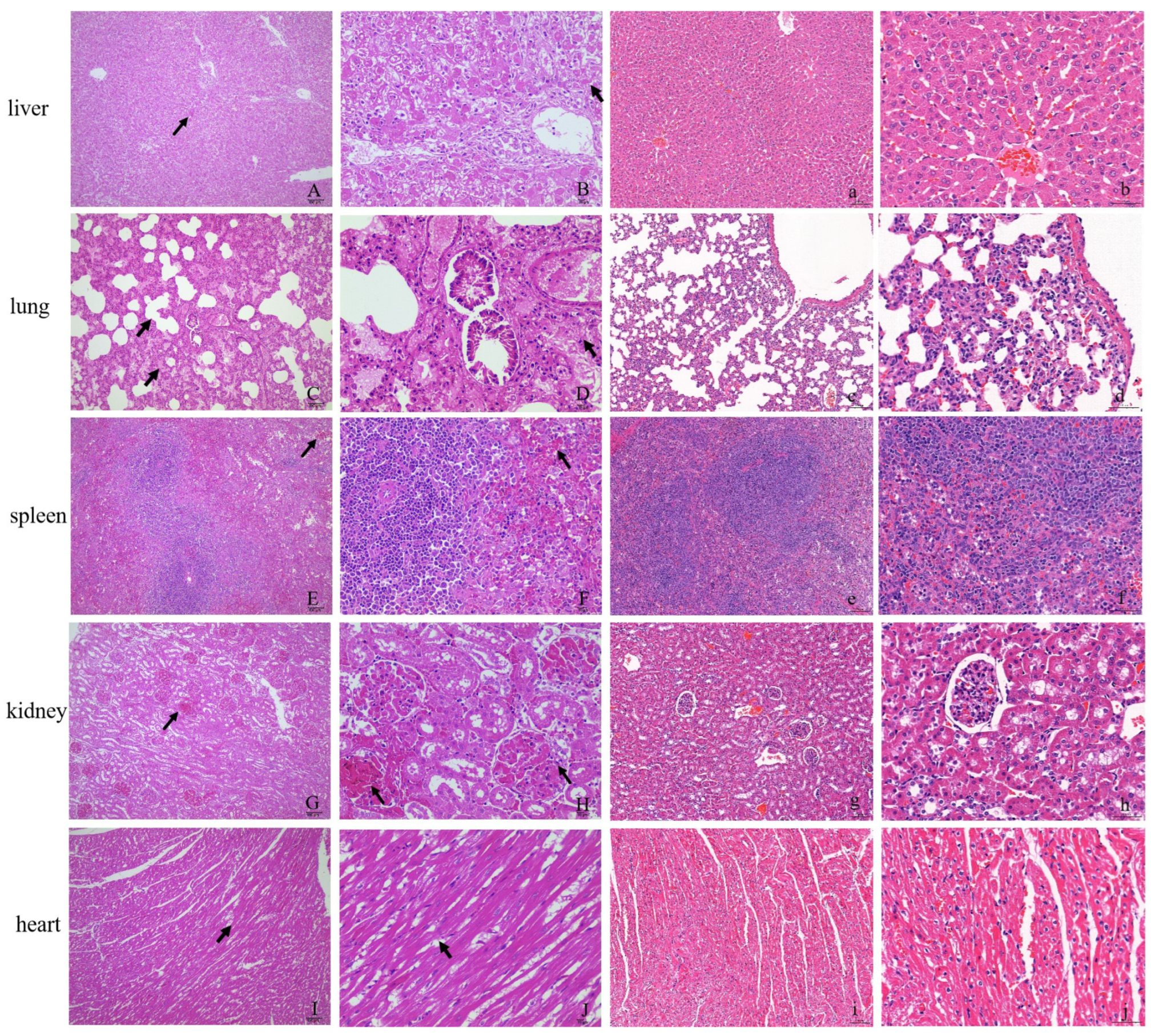
2.3. Histopathological Observation
2.4. Virus Quantification
2.5. cDNA Library Construction and Sequencing
2.6. Pre-Processing of Sequencing Reads and Gene Expression and Differential Gene Analyses
2.7. Gene Ontology (GO) and Pathway Enrichment Analysis
2.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Verification of the Illumina NovaSeq 6000 Sequencing Data
2.9. Statistical Analysis
3. Results
3.1. Clinical Signs, Case Fatality Rates, and Histopathological Changes in Experimentally Infected Rabbits
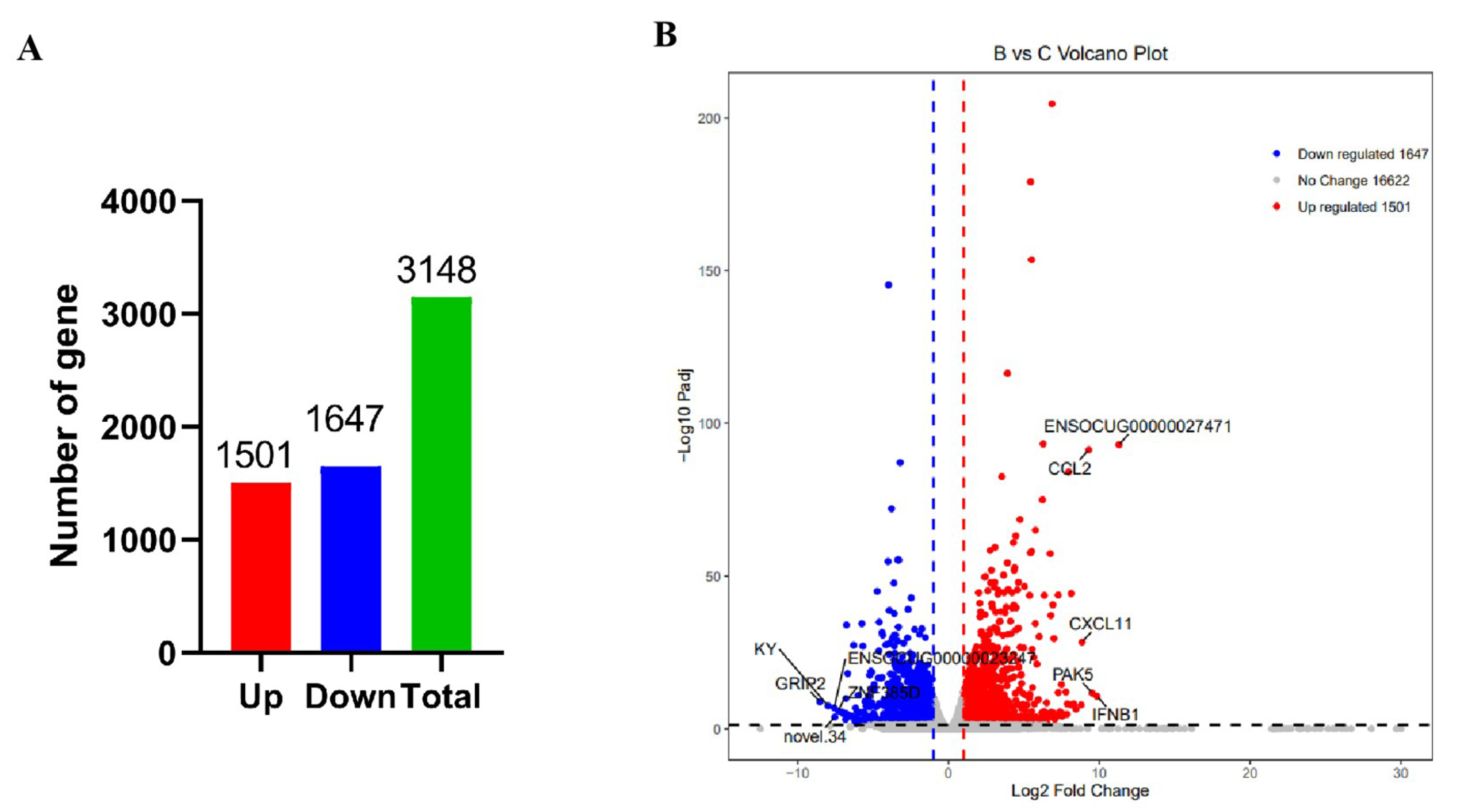
3.2. Viral RNA Burden and Transcriptomic Pattern of Spleen
3.3. Differentially Expressed Gene (DEG) Screening in the Spleen
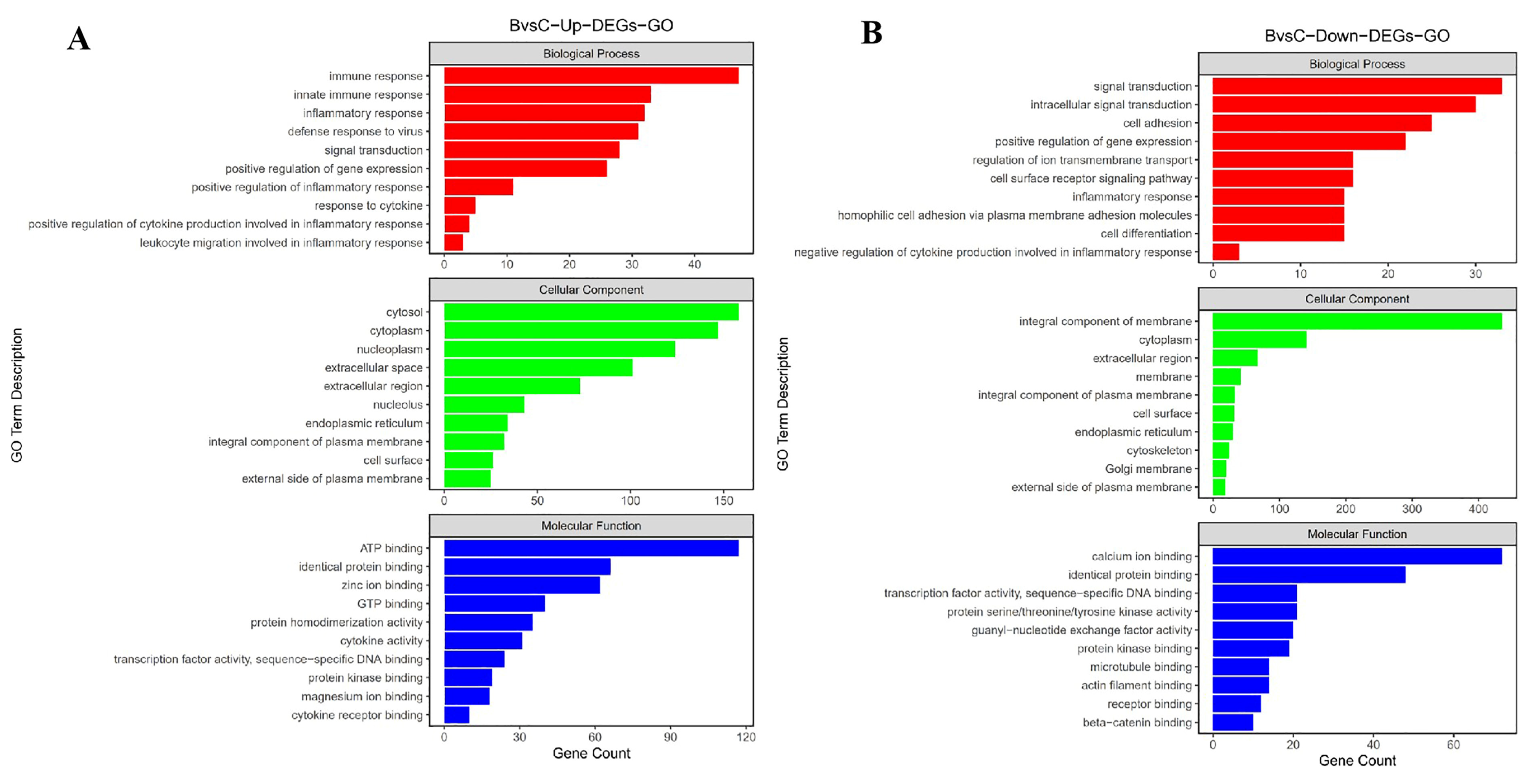
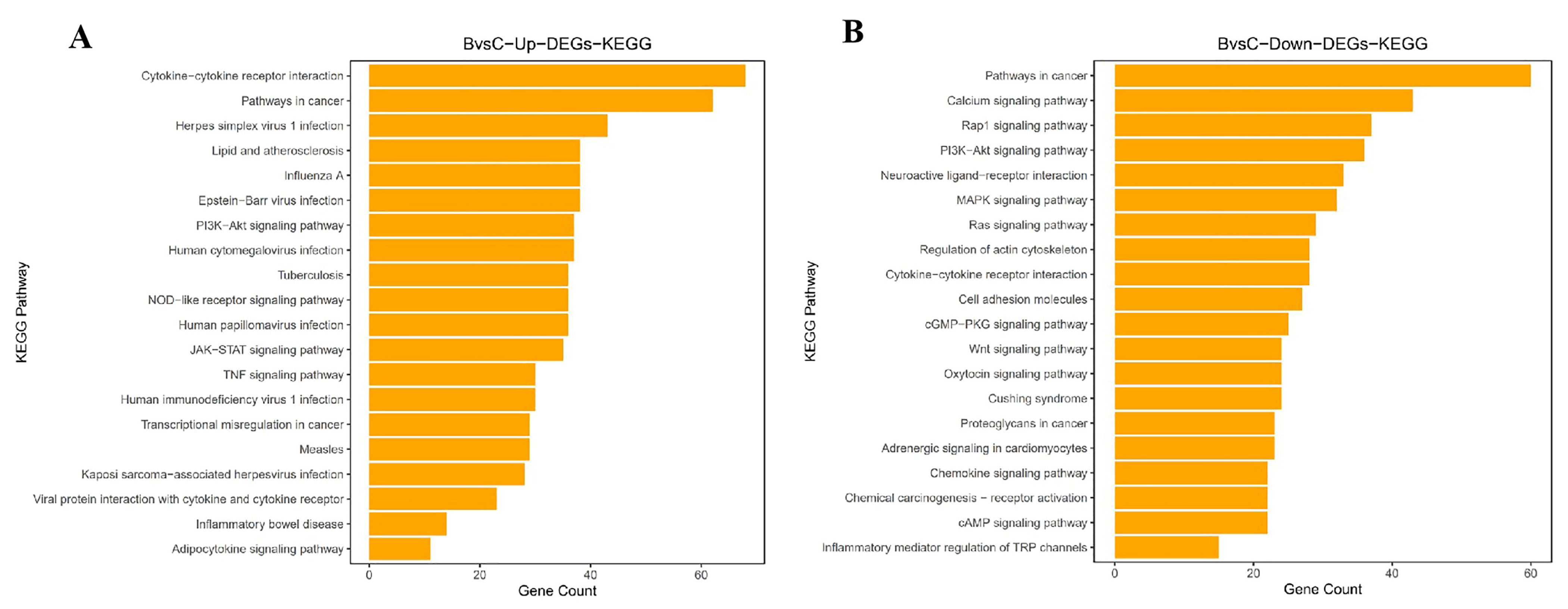
3.4. Functional Analysis and Biological Enrichment of DEGs
3.5. Transcriptome Data Verification by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abrantes, J.; Van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Xue, H.P.; Pu, B.Q.; Qian, N.H. A new viral disease in rabbit. Anim. Husb. Vet. Med. 1984, 16, 253–255. (In Chinese) [Google Scholar]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Reculé, G.; Lopes, A.M.; Esteves, P. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Rabbit hemorrhagic disease virus—molecular cloning and nucleotide sequencing of a calicivirus genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.J.; Thumfart, J.O. Rabbit hemorrhagic disease virus: Genome organization and polyprotein processing of a calicivirus studied after transient expression of cDNA constructs. Virology 2000, 276, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.T.; Li, B.Y.; Zhang, L.; Jiao, W.Q.; Liu, J.X. Bioinformatics analysis of rabbit haemorrhagic disease virus genome. Virol. J. 2011, 8, 494. [Google Scholar] [CrossRef] [PubMed]
- Le Gall-Recule, G.; Zwingelstein, F.; Laurent, S.; de Boisseson, C.; Portejoie, Y.; Rasschaert, D. Phylogenetic analysis of rabbit haemorrhagic disease virus in France between 1993 and 2000, and the characterisation of RHDV antigenic variants. Arch. Virol. 2003, 148, 65–81. [Google Scholar] [CrossRef]
- Capucci, L.; Fallacara, F.; Grazioli, S.; Lavazza, A.; Pacciarini, M.L.; Brocchi, E. A further step in the evolution of rabbit hemorrhagic disease virus: The appearance of the first consistent antigenic variant. Virus Res. 1998, 58, 115–126. [Google Scholar] [CrossRef]
- Le Gall-Recule, G.; Zwingelstein, F.; Boucher, S.; Le Normand, B.; Plassiart, G.; Portejoie, Y.; Decors, A.; Bertagnoli, S.; Guerin, J.-L.; Marchandeau, S. Detection of a new variant of rabbit haemorrhagic disease virus in France. Vet. Rec. 2011, 168, 137–138. [Google Scholar] [CrossRef]
- Dalton, K.P.; Nicieza, I.; Balseiro, A.; Muguerza, M.A.; Rosell, J.M.; Casais, R.; Álvarez, L.; Parra, F. Variant rabbit hemorrhagic disease virus in young rabbits, Spain. Emerg. Infect. Dis. 2012, 18, 2009–2012. [Google Scholar] [CrossRef]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Melo, P.; Correia, J.J.; Ramada, M.; Alves, P.C.; Parra, F.; Esteves, P.J. New variant of rabbit hemorrhagic disease virus, Portugal, 2012–2013. Emerg. Infect. Dis. 2013, 19, 1900–1902. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guérin, J.-L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef]
- Hu, B.; Wei, H.; Fan, Z.; Song, Y.; Chen, M.; Qiu, R.; Wang, F. Emergence of rabbit haemorrhagic disease virus 2 in China in 2020. Vet. Med. Sci. 2021, 7, 236–239. [Google Scholar] [CrossRef]
- Li, Y.; Du, D.; Zhou, L.; Su, L.; You, C.; Zhang, H.; Yu, J.; Xiao, L.; Huang, J. First report of GI.1aP-GI.2 recombinants of rabbit hemorrhagic disease virus in domestic rabbits in China. Front. Microbiol. 2023, 14, 1188380. [Google Scholar] [CrossRef]
- Qi, R.; Meng, C.; Zhu, J.; Li, H.; Miao, Q.; Tang, J.; Tang, A.; Guo, H.; Liu, C.; Li, C.; et al. The outbreak of rabbit hemorrhagic virus type 2 in the interior of China may be related to imported semen. Virol. Sin. 2022, 37, 623–626. [Google Scholar] [CrossRef]
- Yu, J.F.; Xiao, L.; Zhou, L.; Xie, J.; Cao, Y.; Ye, Y.G.; Li, X.Y.; Pan, M.; Ye, J.Q.; Wei, Y.; et al. Complete genome sequencing and genetic evolution analysis of rabbit hemorrhagic disease virus type 2 in China. Heilongjiang Anim. Sci. Vet. Med. 2021, 8, 73–77. (In Chinese) [Google Scholar]
- Neimanis, A.S.; Pettersson, U.L.; Huang, N.; Gavier-Widén, D.; Strive, T. Elucidation of the pathology and tissue distribution of Lagovirus europaeus GI.2/RHDV2 (rabbit haemorrhagic disease virus 2) in young and adult rabbits (Oryctolagus cuniculus). Vet. Res. 2018, 49, 46. [Google Scholar] [CrossRef] [PubMed]
- Neave, M.J.; Hall, R.N.; Huang, N.; McColl, K.A.; Kerr, P.; Hoehn, M.; Taylor, J.; Strive, T. Robust innate immunity of young rabbits mediates resistance to rabbit hemorrhagic disease caused by Lagovirus europaeus GI.1 but not GI.2. Viruses 2018, 19, 512. [Google Scholar] [CrossRef] [PubMed]
- Semerjyan, A.B.; Sargsyan, M.A.; Arzumanyan, H.H.; Hakobyan, L.H.; Abroyan, L.O.; Avetisyan, A.S.; Karalova, E.M.; Manukyan, D.M.; Matevosyan, H.S.; Krasnikov, N.F.; et al. Immune cell pathology in rabbit hemorrhagic disease. Vet. World 2019, 12, 1332–1340. [Google Scholar] [CrossRef]
- Rystwej, N.; Tokarz-Deptuła, B.; Deptuła, W. Reactivity of selected markers of innate and adaptive immunity in rabbits experimentally infected with antigenic variants of RHD (Lagovirus europaeus/GI.1a). Vet. Res. Commun. 2022, 46, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Hrynkiewicz, R.; Bębnowska, D.; Maldonado, J.; Baratelli, M.; Köllner, B.; Niedźwiedzka-Rystwej, P. Immunity against Lagovirus europaeus and the Impact of the Immunological Studies on Vaccination. Vaccines 2021, 9, 255. [Google Scholar] [CrossRef]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. The importance of liver lesions and changes to biochemical and coagulation factors in the pathogenesis of RHD. Acta Biochim. Pol. 2015, 62, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Jenckel, M.; Hall, R.N.; Strive, T. Pathogen profiling of Australian rabbits by metatranscriptomic sequencing. Transbound Emerg Dis. 2022, 69, e2629–e2640. [Google Scholar] [CrossRef]
- Smertina, E.; Carroll, A.J.; Boileau, J.; Emmott, E.; Jenckel, M.; Vohra, H.; Rolland, V.; Hands, P.; Hayashi, J.; Neave, M.J.; et al. Lagovirus non-structural protein p23: A putative viroporin that interacts with heat shock proteins and uses a disulfide bond for dimerization. Front. Microbiol. 2022, 13, 923256. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Yu, J.F.; Lin, Y.; Zhou, L.; Guo, Z.Q.; Xie, J.; Yue, J.G.; Ye, Y.G.; Cao, Y.; Li, X.Y.; et al. First detection of a type 2 rbbbit hemrrhagic disease virus (RHDV2/b/GI.2) in China and its pathological observation. China Anim. Husb. Vet. Med. 2021, 48, 348–355. (In Chinese) [Google Scholar]
- Velarde, R.; Cavadini, P.; Neimanis, A.; Cabezón, O.; Chiari, M.; Gaffuri, A.; Lavín, S.; Grilli, G.; Gavier-Widén, D.; Lavazza, A.; et al. Spillover events of infection of brown hares (Lepus europaeus) with rabbit haemorrhagic disease type 2 Virus (RHDV2) caused sporadic cases of an european brown hare syndrome-like disease in Italy and Spain. Transbound. Emerg. Dis. 2017, 64, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.; Abad-Guamán, R.; Nicodemus, N.; Diaz-Perales, A.; García, J.; Carabaño, R.; Menoyo, D. Effect of pre- and post-weaning dietary supplementation with arginine and glutamine on rabbit performance and intestinal health. BMC Vet. Res. 2019, 13, 199. [Google Scholar] [CrossRef] [PubMed]
- Schnupf, P.; Sansonetti, P.J. Quantitative RT-PCR profiling of the rabbit immune response: Assessment of acute shigella flexneri infection. PLoS ONE 2012, 7, e36446. [Google Scholar] [CrossRef]
- Li, H.C.; Yan, L.; Wang, D.L.; Chu, F.F.; Peng, P.; Xia, W. Imiquimod inhibit scar formation by regulating the expression of Th1/Th2 cell related chemokines in rabbit ear hypertrophic scar model. Prog. Mod. Biomed. 2014, 14. (In Chinese) [Google Scholar]
- Chen, W.; Tu, T.; Luo, Y.; Yang, Z.X.; Yao, X.P.; Wu, X.L.; Wang, Y.J. Detection of a new emerging strain of rabbit haemorrhagic disease virus 2 (GI.2) in China. J. Vet. Res. 2022, 66, 289–295. [Google Scholar] [CrossRef]
- Chen, M.; Fan, Z.; Hu, B.; Song, Y.; Wei, H.; Qiu, R.; Zhu, W.; Xu, W.; Wang, F. Pathogenicity of the newly emerged Lagovirus europaeus GI.2 strain in China in experimentally infected rabbits. Vet. Microbiol. 2022, 265, 109311. [Google Scholar] [CrossRef]
- Dalton, K.; Balseiro, A.; Juste, R.; Podadera, A.; Nicieza, I.; del Llano, D.; González, R.; Alonso, J.M.; Prieto, J.; Parra, F.; et al. Clinical course and pathogenicity of variant rabbit haemorrhagic disease virus in experimentally infected adult and kit rabbits: Significance towards control and spread. Vet. Microbiol. 2018, 220, 24–32. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef]
- Sokol, C.L.; Luster, A.D. The chemokine system in innate immunity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016303. [Google Scholar] [CrossRef]
- Franzoni, G.; Pedrera, M.; Sánchez-Cordón, P.J. African swine fever virus infection and cytokine response in vivo: An update. Viruses 2023, 15, 233. [Google Scholar] [CrossRef]
- Song, P.P.; Li, W.; Xie, J.Q.; Hou, Y.L.; You, C.G. Cytokine storm induced by SARS-CoV-2. Clin. Chim. Acta. 2020, 509, 280–287. [Google Scholar] [CrossRef]
- Rose-John, S. Interleukin-6 family cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak-Ryczek, A.; Tokarz-Deptula, B.; Deptula, W. Expression of IL-1Ra, IL-6, IL-8, IL-18, TNF-alpha and IFN-gamma genes in peripheral blood leukocytes of rabbits infected with RHDV (Rabbit Haemorrhagic Disease Virus). Dev. Comp. Immunol. 2017, 76, 310–315. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, A.D.; Mohamed, F.M.; Zhang, J.; Brown, C.C. Early pathogenesis in rabbit hemorrhagic disease virus 2. Microb. Pathog. 2022, 173, 105814. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Jiang, S.C.; Zhang, Z.W.; Fu, Y.F.; Hu, J.; Li, Z.L. Quantification of cytokine storms during virus infections. Front. Immunol. 2021, 17, 659419. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Liang, Z.A.; Xue, L.Z. Neuromedin U: Potential roles in immunity and inflammation. Immunology 2021, 162, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.Y.; Song, D.; Li, H.C.; He, M.L. Stress proteins: The biological functions in virus infection, present and challenges for target-based antiviral drug development. Signal Transduct. Target. Ther. 2020, 5, 125. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Time of Death Postinfection (PI) | Clinical Symptoms | Number of Deaths | Mortality (%) |
|---|---|---|---|---|
| Infection | between 12/16 h | anorexia, lethargy | 1 | 100% |
| between 17/20 h | anorexia, lethargy | 1 | ||
| between 21/24 h | anorexia, lethargy, bleeding from the nose | 3 | ||
| Control | - | - | none | 0% |
| Sample Group | Sample Name | Clean Data (bp) | Clean Reads Number | Clean Rate (%) | Genome Mapping (%) | Detected Gene NO. |
|---|---|---|---|---|---|---|
| Infection group | B | 6,791,407,800 | 45276052 | 96.48% | 87.74% | 17,552 |
| 7,982,321,400 | 53215476 | 97.79% | 86.72% | 17,532 | ||
| 6,785,876,400 | 45239176 | 97.14% | 86.42% | 17,392 | ||
| Control group | C | 6,863,618,100 | 45757454 | 96.97% | 86.96% | 17,963 |
| 6,705,405,900 | 44702706 | 97.82% | 86.31% | 17,829 | ||
| 8,044,070,100 | 53627134 | 97.39% | 87.52% | 17,993 |
| Differentially Expressed Genes | IL1α | IL-6 | IL-8 | IL-22 | CCL2 | CXCL9 | NMUR1 | HSPB7 | KCNIP2 |
|---|---|---|---|---|---|---|---|---|---|
| qRT-PCR (2−ΔΔct) | 4.85 ± 1.08 | 27.15 ± 3.11 | 19.72 ± 16.44 | 6.24 ± 1.88 | 122.42 ± 28.51 | 7.75 ± 5.07 | 0.28 ± 0.07 | 0.19 ± 0.12 | 0.27 ± 0.07 |
| RNA-seq (log2) | 4.97 | 11.31 | 6.35 | 6.58 | 9.30 | 6.91 | −5.81 | −6.18 | −6.08 |
| expression pattern | up | up | up | up | up | up | down | down | down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Li, Y.; Xiao, L.; Xie, J.; Guo, Z.; Ye, Y.; Lin, Y.; Cao, Y.; Wu, X.; Mao, C.; et al. Neglected Spleen Transcriptional Profile Reveals Inflammatory Disorder Conferred by Rabbit Hemorrhagic Disease Virus 2 Infection. Viruses 2024, 16, 495. https://doi.org/10.3390/v16040495
Yu J, Li Y, Xiao L, Xie J, Guo Z, Ye Y, Lin Y, Cao Y, Wu X, Mao C, et al. Neglected Spleen Transcriptional Profile Reveals Inflammatory Disorder Conferred by Rabbit Hemorrhagic Disease Virus 2 Infection. Viruses. 2024; 16(4):495. https://doi.org/10.3390/v16040495
Chicago/Turabian StyleYu, Jifeng, Yan Li, Lu Xiao, Jing Xie, Zhiqiang Guo, Yonggang Ye, Yi Lin, Ye Cao, Xuejing Wu, Congjian Mao, and et al. 2024. "Neglected Spleen Transcriptional Profile Reveals Inflammatory Disorder Conferred by Rabbit Hemorrhagic Disease Virus 2 Infection" Viruses 16, no. 4: 495. https://doi.org/10.3390/v16040495