
Herpes Simplex Virus 1 Glycoprotein B from a Hyperfusogenic Virus Mediates Enhanced Cell–Cell Fusion
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Effect of Reduced Temperature on HSV Entry into Vero Cells
2.3. Sequencing Analysis
2.4. Construction of Expression Vectors
2.5. Antibodies
2.6. SDS-PAGE and Western Blot
2.7. CELISA
2.8. Virus-Free Luciferase Reporter Assay for Cell–Cell Fusion
3. Results
3.1. HSV-1 ANG Enters Vero Cells at 4 °C
3.2. HSV-1 ANG and Its Derivative ANG Path Harbor Mutations in Entry and Fusion Glycoproteins
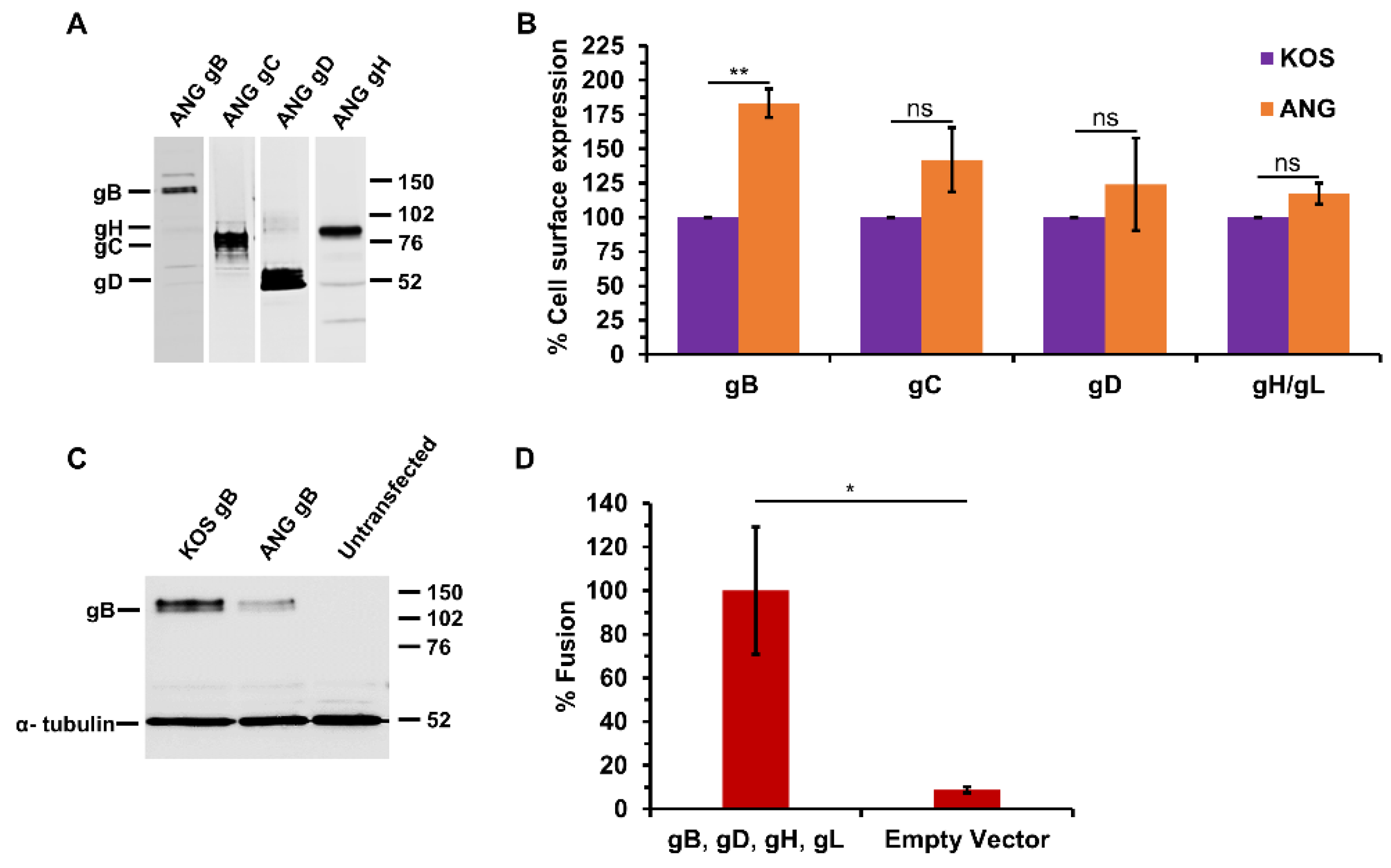
3.3. HSV-1 ANG gB, gD, gH, and gL Are Sufficient for Cell–Cell Fusion
3.4. ANG gB Exhibits Hyperfusogenic Activity for Cell–Cell Fusion When Co-Expressed with Wild Type gD and gH/gL
3.5. HSV-1 ANG Alleles of gD or gH/gL Do Not Alter Cell–Cell Fusion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Connolly, S.A.; Jardetzky, T.S.; Longnecker, R. The structural basis of herpesvirus entry. Nat. Rev. Microbiol. 2021, 19, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Krummenacher, C.; Carfi, A.; Eisenberg, R.J.; Cohen, G.H. Entry of herpesviruses into cells: The enigma variations. Adv. Exp. Med. Biol. 2013, 790, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Nicola, A.V. Herpesvirus Entry into Host Cells Mediated by Endosomal Low pH. Traffic 2016, 17, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Weed, D.J.; Nicola, A.V. Herpes simplex virus Membrane Fusion. Adv. Anat. Embryol. Cell Biol. 2017, 223, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.; Bruun, B.; Minson, T.; Browne, H. Glycoproteins gB, gD, and gHgL of herpes simplex virus type 1 are necessary and sufficient to mediate membrane fusion in a Cos cell transfection system. J. Virol. 1998, 72, 873–875. [Google Scholar] [CrossRef]
- Nicola, A.V.; Hou, J.; Major, E.O.; Straus, S.E. Herpes simplex virus type 1 enters human epidermal keratinocytes, but not neurons, via a pH-dependent endocytic pathway. J. Virol. 2005, 79, 7609–7616. [Google Scholar] [CrossRef]
- Nicola, A.V.; McEvoy, A.M.; Straus, S.E. Roles for endocytosis and low pH in herpes simplex virus entry into HeLa and Chinese hamster ovary cells. J. Virol. 2003, 77, 5324–5332. [Google Scholar] [CrossRef]
- Munk, K.; Donner, D. Cytopathischer effekt und plaque-morphologie verschiedener herpes-simplex-virus-stämme. Arch. Für Die Gesamte Virusforsch 1963, 13, 529–540. [Google Scholar] [CrossRef]
- Kaerner, H.C.; Schroder, C.H.; Ott-Hartmann, A.; Kumel, G.; Kirchner, H. Genetic variability of herpes simplex virus: Development of a pathogenic variant during passaging of a nonpathogenic herpes simplex virus type 1 virus strain in mouse brain. J. Virol. 1983, 46, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Falke, D.; Knoblich, A.; Muller, S. Fusion from without induced by herpes simplex virus type 1. Intervirology 1985, 24, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Lingen, M.; Seck, T.; Weise, K.; Falke, D. Single amino acid substitutions in the glycoprotein B carboxy terminus influence the fusion from without property of herpes simplex virus type 1. J. Gen. Virol. 1995, 76 Pt 7, 1843–1849. [Google Scholar] [CrossRef]
- Saharkhiz-Langroodi, A.; Holland, T.C. Identification of the fusion-from-without determinants of herpes simplex virus type 1 glycoprotein B. Virology 1997, 227, 153–159. [Google Scholar] [CrossRef]
- Dean, H.J.; Warner, M.S.; Terhune, S.S.; Johnson, R.M.; Spear, P.G. Viral determinants of the variable sensitivity of herpes simplex virus strains to gD-mediated interference. J. Virol. 1995, 69, 5171–5176. [Google Scholar] [CrossRef]
- Izumi, K.M.; Stevens, J.G. Molecular and biological characterization of a herpes simplex virus type 1 (HSV-1) neuroinvasiveness gene. J. Exp. Med. 1990, 172, 487–496. [Google Scholar] [CrossRef]
- Lopez, M.; Cocchi, F.; Menotti, L.; Avitabile, E.; Dubreuil, P.; Campadelli-Fiume, G. Nectin2alpha (PRR2alpha or HveB) and nectin2delta are low-efficiency mediators for entry of herpes simplex virus mutants carrying the Leu25Pro substitution in glycoprotein D. J. Virol. 2000, 74, 1267–1274. [Google Scholar] [CrossRef]
- Warner, M.S.; Geraghty, R.J.; Martinez, W.M.; Montgomery, R.I.; Whitbeck, J.C.; Xu, R.; Eisenberg, R.J.; Cohen, G.H.; Spear, P.G. A cell surface protein with herpesvirus entry activity (HveB) confers susceptibility to infection by mutants of herpes simplex virus type 1, herpes simplex virus type 2, and pseudorabies virus. Virology 1998, 246, 179–189. [Google Scholar] [CrossRef]
- Delboy, M.G.; Patterson, J.L.; Hollander, A.M.; Nicola, A.V. Nectin-2-mediated entry of a syncytial strain of herpes simplex virus via pH-independent fusion with the plasma membrane of Chinese hamster ovary cells. Virol. J. 2006, 3, 105. [Google Scholar] [CrossRef] [PubMed]
- Roller, D.G.; Dollery, S.J.; Doyle, J.L.; Nicola, A.V. Structure-function analysis of herpes simplex virus glycoprotein B with fusion-from-without activity. Virology 2008, 382, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Nicola, A.V.; De Leon, M.P.; Xu, R.; Hou, W.; Whitbeck, J.C.; Krummenacher, C.; Montgomery, R.I.; Spear, P.G.; Eisenberg, R.J.; Cohen, G.H. Monoclonal antibodies to distinct sites on herpes simplex virus (HSV) glycoprotein D block HSV binding to HVEM. J. Virol. 1998, 72, 3595–3601. [Google Scholar] [CrossRef] [PubMed]
- Dean, H.J.; Terhune, S.S.; Shieh, M.-T.; Susmarski, N.; Spear, P.G. Single Amino Acid Substitutions in gD of Herpes Simplex Virus I Confer Resistance to gD-Mediated Interference and Cause Cell-Type-Dependent Alterations in Infectivity. Virology 1994, 199, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Hitoshi, N.; Ken-ichi, Y.; Jun-ichi, M. Efficient selection for high-expression transfectants with a novel eukaryotic vector. Gene 1991, 108, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Kobasa, D.; Rodgers, M.E.; Wells, K.; Kawaoka, Y. Neuraminidase hemadsorption activity, conserved in avian influenza A viruses, does not influence viral replication in ducks. J. Virol. 1997, 71, 6706–6713. [Google Scholar] [CrossRef]
- Kousoulas, K.G.; Pellett, P.E.; Pereira, L.; Roizman, B. Mutations affecting conformation or sequence of neutralizing epitopes identified by reactivity of viable plaques segregate from syn and ts domains of HSV-1(F) gB gene. Virology 1984, 135, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Ali, M.; Kousoulas, K.; Huo, B.; Banks, T. Domain structure of herpes simplex virus 1 glycoprotein B: Neutralizing epitopes map in regions of continuous and discontinuous residues. Virology 1989, 172, 11–24. [Google Scholar] [CrossRef]
- Friedman, H.M.; Cohen, G.H.; Eisenberg, R.J.; Seidel, C.A.; Cines, D.B. Glycoprotein C of herpes simplex virus 1 acts as a receptor for the C3b complement component on infected cells. Nature 1984, 309, 633–635. [Google Scholar] [CrossRef]
- Eisenberg, R.J.; Ponce de Leon, M.; Friedman, H.M.; Fries, L.F.; Frank, M.M.; Hastings, J.C.; Cohen, G.H. Complement component C3b binds directly to purified glycoprotein C of herpes simplex virus types 1 and 2. Microb. Pathog. 1987, 3, 423–435. [Google Scholar] [CrossRef]
- Eisenberg, R.J.; Long, D.; Ponce de Leon, M.; Matthews, J.T.; Spear, P.G.; Gibson, M.G.; Lasky, L.A.; Berman, P.; Golub, E.; Cohen, G.H. Localization of epitopes of herpes simplex virus type 1 glycoprotein D. J. Virol. 1985, 53, 634–644. [Google Scholar] [CrossRef]
- Pertel, P.E.; Fridberg, A.; Parish, M.L.; Spear, P.G. Cell fusion induced by herpes simplex virus glycoproteins gB, gD, and gH-gL requires a gD receptor but not necessarily heparan sulfate. Virology 2001, 279, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Sodeik, B.; Ebersold, M.W.; Helenius, A. Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J. Cell Biol. 1997, 136, 1007–1021. [Google Scholar] [CrossRef]
- Nicola, A.V.; Straus, S.E. Cellular and viral requirements for rapid endocytic entry of herpes simplex virus. J. Virol. 2004, 78, 7508–7517. [Google Scholar] [CrossRef]
- Herold, B.C.; WuDUNN, D.; Soltys, N.; Spear, P.G. Glycoprotein C of herpes simplex virus type 1 plays a principal role in the adsorption of virus to cells and in infectivity. J. Virol. 1991, 65, 1090–1098. [Google Scholar] [CrossRef]
- Sari, T.K.; Gianopulos, K.A.; Weed, D.J.; Schneider, S.M.; Pritchard, S.M.; Nicola, A.V. Herpes Simplex Virus Glycoprotein C Regulates Low-pH Entry. Msphere 2020, 5, e00826-19. [Google Scholar] [CrossRef]
- Gianopulos, K.A.; Komala Sari, T.; Weed, D.J.; Pritchard, S.M.; Nicola, A.V. Conformational Changes in Herpes Simplex Virus Glycoprotein C. J. Virol. 2022, 96, e00163–22. [Google Scholar] [CrossRef] [PubMed]
- Dollery, S.J.; Lane, K.D.; Delboy, M.G.; Roller, D.G.; Nicola, A.V. Role of the UL45 protein in herpes simplex virus entry via low pH-dependent endocytosis and its relationship to the conformation and function of glycoprotein B. Virus Res. 2010, 149, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Siekavizza-Robles, C.R.; Dollery, S.J.; Nicola, A.V. Reversible conformational change in herpes simplex virus glycoprotein B with fusion-from-without activity is triggered by mildly acidic pH. Virol. J. 2010, 7, 352. [Google Scholar] [CrossRef]
- Sayers, C.L.; Elliott, G. Herpes Simplex Virus 1 Enters Human Keratinocytes by a Nectin-1-Dependent, Rapid Plasma Membrane Fusion Pathway That Functions at Low Temperature. J. Virol. 2016, 90, 10379–10389. [Google Scholar] [CrossRef] [PubMed]
- De La Cruz, N.; Knebel-Morsdorf, D. Endocytic Internalization of Herpes Simplex Virus 1 in Human Keratinocytes at Low Temperature. J. Virol. 2021, 95, e02195-20. [Google Scholar] [CrossRef] [PubMed]
- Bzik, D.J.; Fox, B.A.; DeLuca, N.A.; Person, S. Nucleotide sequence of a region of the herpes simplex virus type 1 gB glycoprotein gene: Mutations affecting rate of virus entry and cell fusion. Virology 1984, 137, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.H.; Gu, B.; Person, S. Role of glycoprotein B of herpes simplex virus type 1 in viral entry and cell fusion. J. Virol. 1988, 62, 2596–2604. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.Z.; Person, S.; DebRoy, C.; Gu, B.H. Functional regions and structural features of the gB glycoprotein of herpes simplex virus type 1. An analysis of linker insertion mutants. J. Mol. Biol. 1988, 201, 575–588. [Google Scholar] [CrossRef]
- Gage, P.J.; Levine, M.; Glorioso, J.C. Syncytium-inducing mutations localize to two discrete regions within the cytoplasmic domain of herpes simplex virus type 1 glycoprotein B. J. Virol. 1993, 67, 2191–2201. [Google Scholar] [CrossRef]
- Engel, J.P.; Boyer, E.P.; Goodman, J.L. Two novel single amino acid syncytial mutations in the carboxy terminus of glycoprotein B of herpes simplex virus type 1 confer a unique pathogenic phenotype. Virology 1993, 192, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Pataki, Z.; Sanders, E.K.; Heldwein, E.E. A surface pocket in the cytoplasmic domain of the herpes simplex virus fusogen gB controls membrane fusion. PLoS Pathog. 2022, 18, e1010435. [Google Scholar] [CrossRef] [PubMed]
- Ruel, N.; Zago, A.; Spear, P.G. Alanine substitution of conserved residues in the cytoplasmic tail of herpes simplex virus gB can enhance or abolish cell fusion activity and viral entry. Virology 2006, 346, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.L.; Greene, N.G.; King, D.S.; Heldwein, E.E. Membrane requirement for folding of the herpes simplex virus 1 gB cytodomain suggests a unique mechanism of fusion regulation. J. Virol. 2012, 86, 8171–8184. [Google Scholar] [CrossRef] [PubMed]
- Weed, D.J.; Dollery, S.J.; Komala Sari, T.; Nicola, A.V. Acidic pH Mediates Changes in Antigenic and Oligomeric Conformation of Herpes Simplex Virus gB and Is a Determinant of Cell-Specific Entry. J. Virol. 2018, 92, e01034-18. [Google Scholar] [CrossRef] [PubMed]
- Dollery, S.J.; Delboy, M.G.; Nicola, A.V. Low pH-induced conformational change in herpes simplex virus glycoprotein B. J. Virol. 2010, 84, 3759–3766. [Google Scholar] [CrossRef]
- Komala Sari, T.; Gianopulos, K.A.; Nicola, A.V. Glycoprotein C of herpes simplex virus 1 shields glycoprotein B from antibody neutralization. J. Virol. 2020, 94, e01852-19. [Google Scholar] [CrossRef] [PubMed]
- David, A.T.; Saied, A.; Charles, A.; Subramanian, R.; Chouljenko, V.N.; Kousoulas, K.G. A herpes simplex virus 1 (McKrae) mutant lacking the glycoprotein K gene is unable to infect via neuronal axons and egress from neuronal cell bodies. MBio 2012, 3, e00144-12. [Google Scholar] [CrossRef]
- Chowdhury, S.; Chouljenko, V.N.; Naderi, M.; Kousoulas, K.G. The amino terminus of herpes simplex virus 1 glycoprotein K is required for virion entry via the paired immunoglobulin-like type-2 receptor alpha. J. Virol. 2013, 87, 3305–3313. [Google Scholar] [CrossRef]
- Atanasiu, D.; Whitbeck, J.C.; Cairns, T.M.; Reilly, B.; Cohen, G.H.; Eisenberg, R.J. Bimolecular complementation reveals that glycoproteins gB and gH/gL of herpes simplex virus interact with each other during cell fusion. Proc. Natl. Acad. Sci. USA 2007, 104, 18718–18723. [Google Scholar] [CrossRef] [PubMed]
- Atanasiu, D.; Saw, W.T.; Cohen, G.H.; Eisenberg, R.J. Cascade of events governing cell-cell fusion induced by herpes simplex virus glycoproteins gD, gH/gL, and gB. J. Virol. 2010, 84, 12292–12299. [Google Scholar] [CrossRef] [PubMed]
- Avitabile, E.; Forghieri, C.; Campadelli-Fiume, G. Complexes between herpes simplex virus glycoproteins gD, gB, and gH detected in cells by complementation of split enhanced green fluorescent protein. J. Virol. 2007, 81, 11532–11537. [Google Scholar] [CrossRef] [PubMed]
- Avitabile, E.; Forghieri, C.; Campadelli-Fiume, G. Cross talk among the glycoproteins involved in herpes simplex virus entry and fusion: The interaction between gB and gH/gL does not necessarily require gD. J. Virol. 2009, 83, 10752–10760. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, F.; Fusco, D.; Menotti, L.; Gianni, T.; Eisenberg, R.J.; Cohen, G.H.; Campadelli-Fiume, G. The soluble ectodomain of herpes simplex virus gD contains a membrane-proximal pro-fusion domain and suffices to mediate virus entry. Proc. Natl. Acad. Sci. USA 2004, 101, 7445–7450. [Google Scholar] [CrossRef] [PubMed]
- Krummenacher, C.; Supekar, V.M.; Whitbeck, J.C.; Lazear, E.; Connolly, S.A.; Eisenberg, R.J.; Cohen, G.H.; Wiley, D.C.; Carfi, A. Structure of unliganded HSV gD reveals a mechanism for receptor-mediated activation of virus entry. EMBO J. 2005, 24, 4144–4153. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.R.; Saw, W.T.; Atanasiu, D.; Lou, H.; Eisenberg, R.J.; Cohen, G.H. Displacement of the C terminus of herpes simplex virus gD is sufficient to expose the fusion-activating interfaces on gD. J. Virol. 2013, 87, 12656–12666. [Google Scholar] [CrossRef] [PubMed]
- Atanasiu, D.; Cairns, T.M.; Whitbeck, J.C.; Saw, W.T.; Rao, S.; Eisenberg, R.J.; Cohen, G.H. Regulation of herpes simplex virus gB-induced cell-cell fusion by mutant forms of gH/gL in the absence of gD and cellular receptors. MBio 2013, 4, e00046-13. [Google Scholar] [CrossRef]
- Dollery, S.J.; Wright, C.C.; Johnson, D.C.; Nicola, A.V. Low-pH-dependent changes in the conformation and oligomeric state of the prefusion form of herpes simplex virus glycoprotein B are separable from fusion activity. J. Virol. 2011, 85, 9964–9973. [Google Scholar] [CrossRef]
- Weed, D.J.; Pritchard, S.M.; Gonzalez, F.; Aguilar, H.C.; Nicola, A.V. Mildly Acidic pH Triggers an Irreversible Conformational Change in the Fusion Domain of Herpes Simplex Virus 1 Glycoprotein B and Inactivation of Viral Entry. J. Virol. 2017, 91, e02123-16. [Google Scholar] [CrossRef]
- Fan, Q.; Hippler, D.P.; Yang, Y.; Longnecker, R.; Connolly, S.A. Multiple Sites on Glycoprotein H (gH) Functionally Interact with the gB Fusion Protein to Promote Fusion during Herpes Simplex Virus (HSV) Entry. MBio 2023, 14, e03368-22. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HSV-1 (ANG) Envelope Protein | Amino Acid Substitutions Relative to HSV-1 (WT) |
|---|---|
| gB | D285N, D329N, V553A, E830K, A855V |
| gD | L25P, Q27R, T230I, A346G |
| gH | A138L, S138L, T150V, A150V, E304K, E459K |
| gL | K90R, V100G, N115D, P168L, P196S |
| gC | R151H, M163T, I182V, T238I, V300D |
| UL45 | None |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gianopulos, K.A.; Makio, A.O.; Pritchard, S.M.; Cunha, C.W.; Hull, M.A.; Nicola, A.V. Herpes Simplex Virus 1 Glycoprotein B from a Hyperfusogenic Virus Mediates Enhanced Cell–Cell Fusion. Viruses 2024, 16, 251. https://doi.org/10.3390/v16020251
Gianopulos KA, Makio AO, Pritchard SM, Cunha CW, Hull MA, Nicola AV. Herpes Simplex Virus 1 Glycoprotein B from a Hyperfusogenic Virus Mediates Enhanced Cell–Cell Fusion. Viruses. 2024; 16(2):251. https://doi.org/10.3390/v16020251
Chicago/Turabian StyleGianopulos, Katrina A., Albina O. Makio, Suzanne M. Pritchard, Cristina W. Cunha, McKenna A. Hull, and Anthony V. Nicola. 2024. "Herpes Simplex Virus 1 Glycoprotein B from a Hyperfusogenic Virus Mediates Enhanced Cell–Cell Fusion" Viruses 16, no. 2: 251. https://doi.org/10.3390/v16020251