
Ecological Niche Modeling of Aedes and Culex Mosquitoes: A Risk Map for Chikungunya and West Nile Viruses in Zambia
 , , ,
, , ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
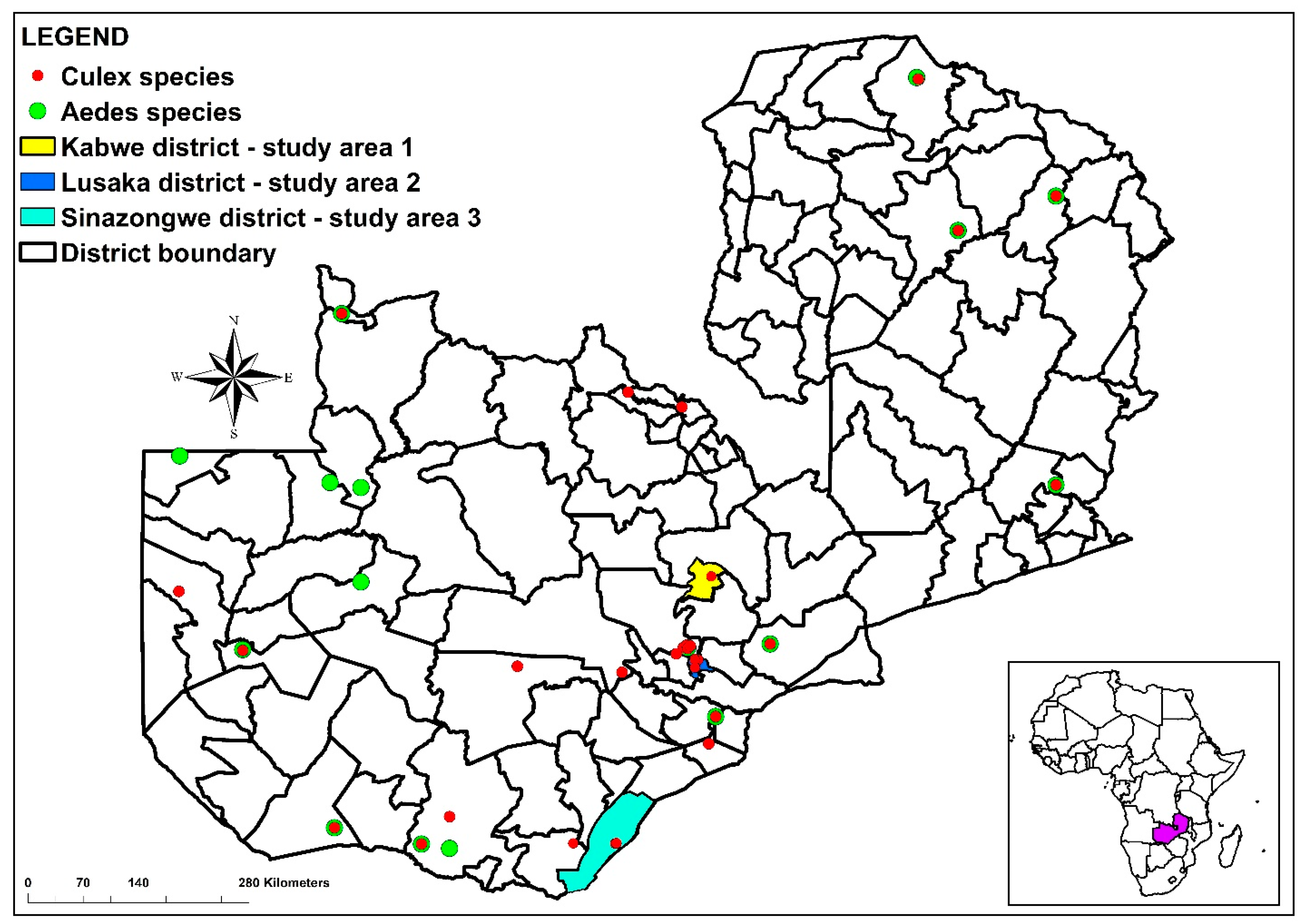
2.1. Study Areas
2.2. Mosquito Occurrence Data
2.3. Environmental Covariates
Modeling Procedure
3. Results
3.1. Distribution of Mosquitoes across the Sampling Areas
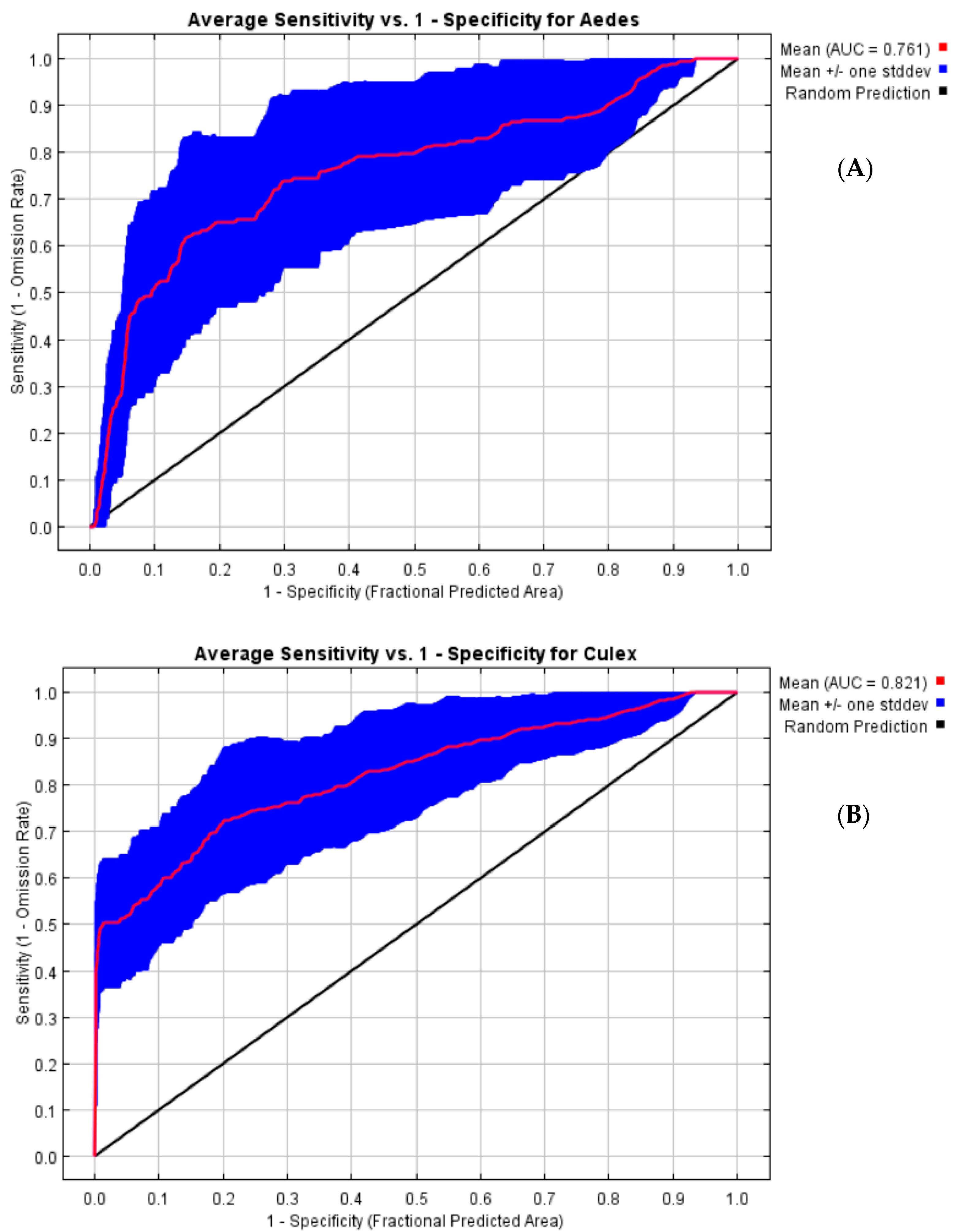
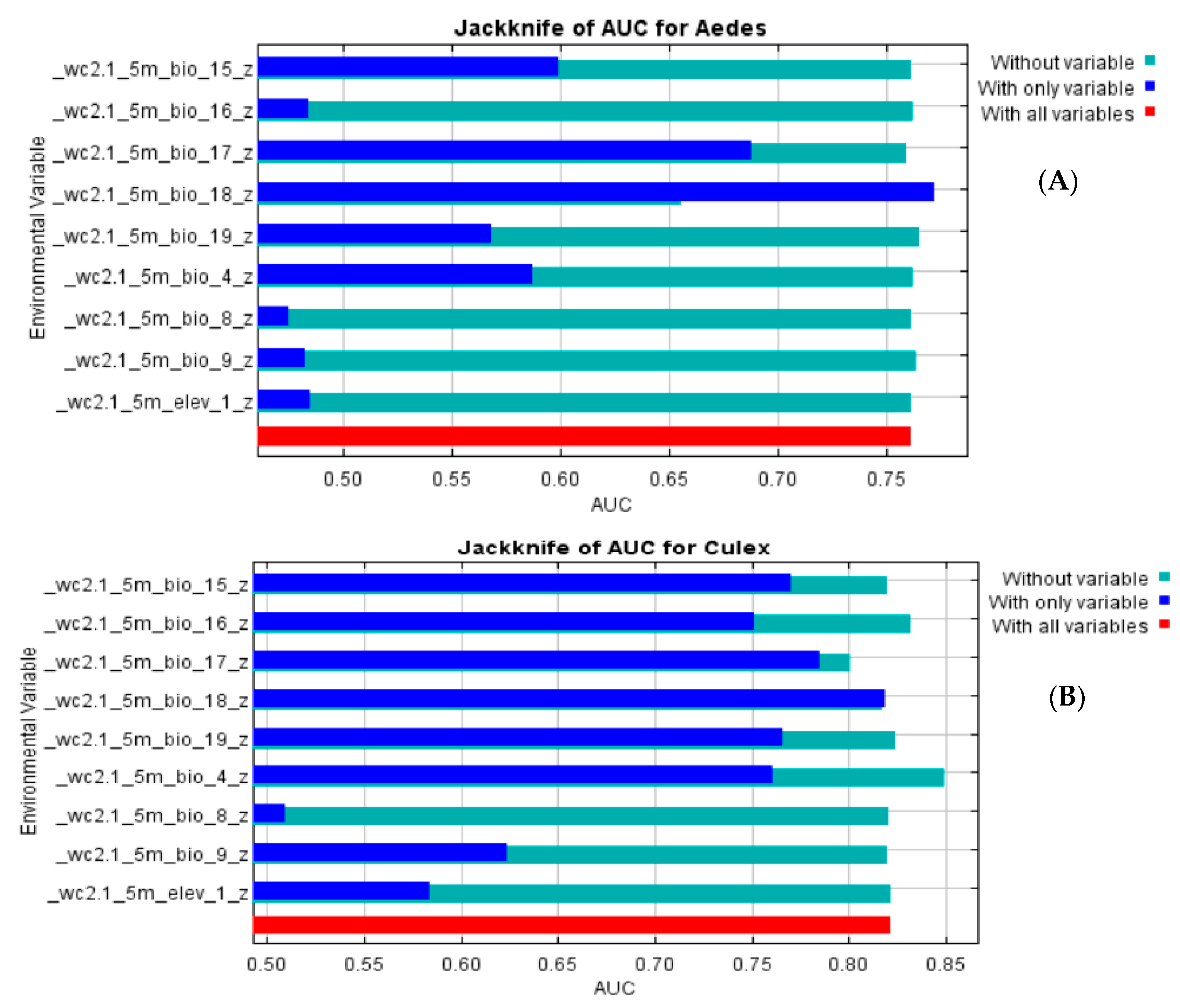
3.2. Model Performance
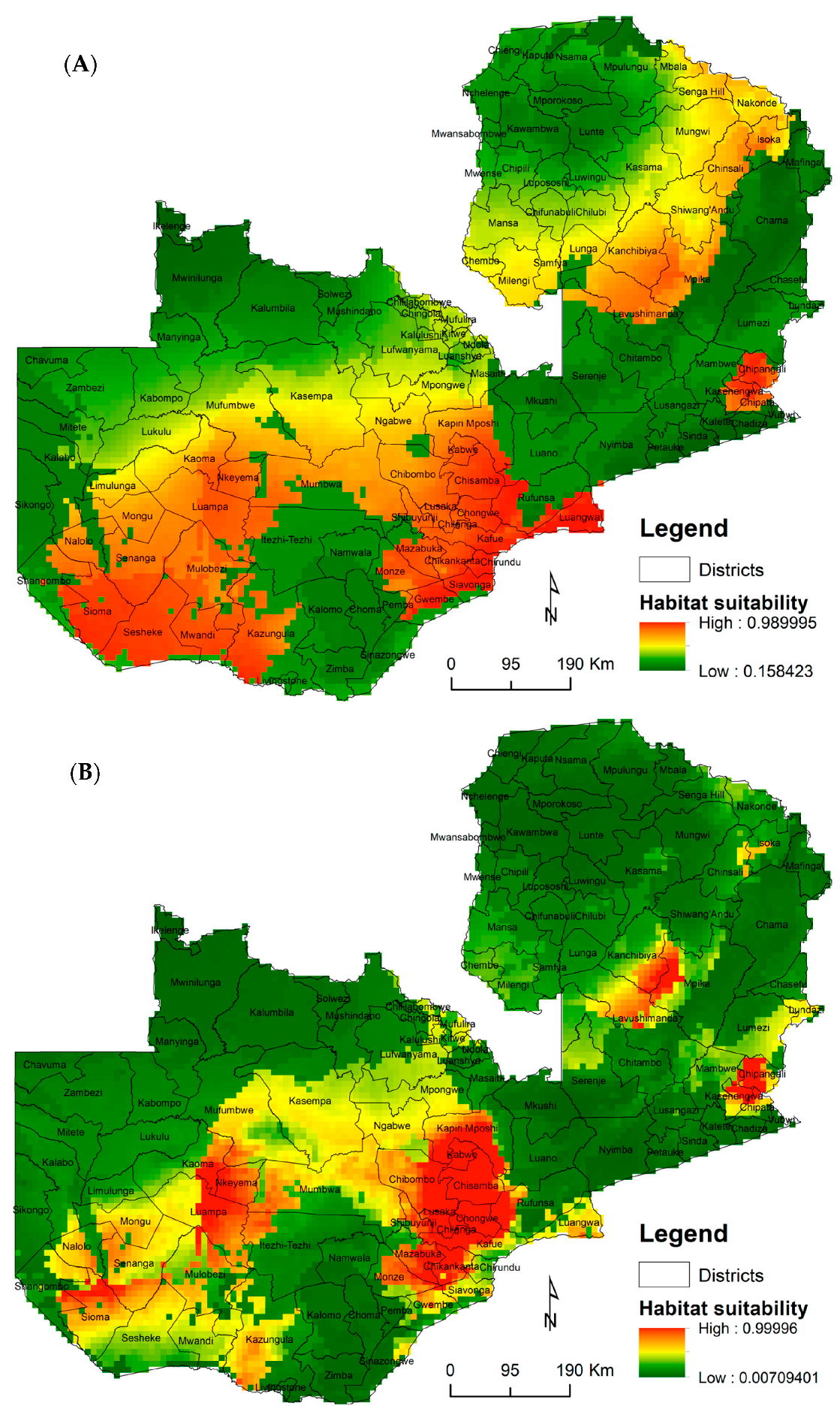
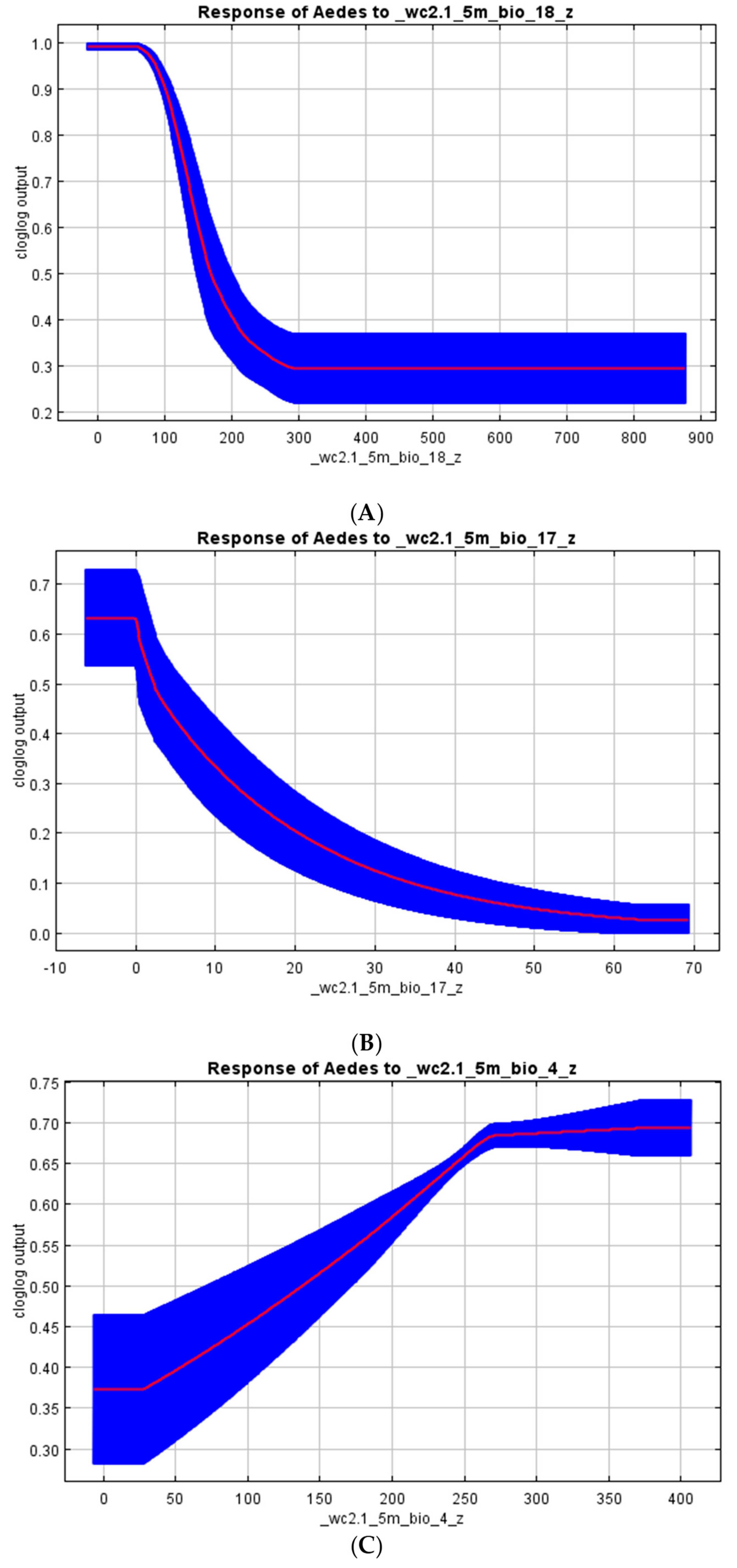
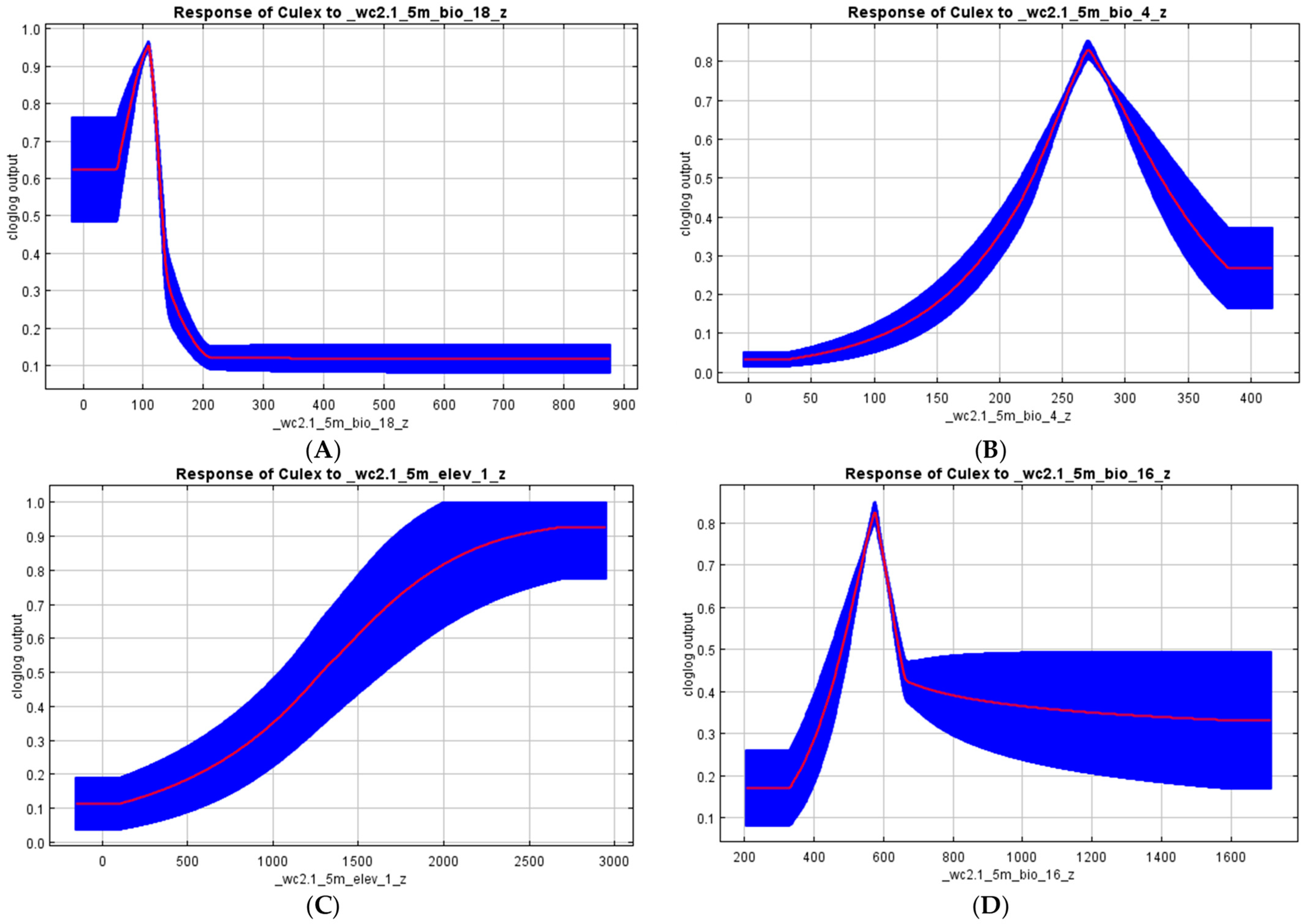
3.3. Vectors’ Habitat Suitability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zerbo, A.; Delgado, R.C.; González, P.A. Aedes-borne viral infections and risk of emergence/resurgence in Sub-Saharan African urban areas. J. Biosaf. Biosecurity 2020, 2, 58–63. [Google Scholar] [CrossRef]
- Girard, M.; Nelson, C.B.; Picot, V.; Gubler, D.J. Arboviruses: A global public health threat. Vaccine 2020, 38, 3989–3994. [Google Scholar] [CrossRef] [PubMed]
- Wahid, B.; Ali, A.; Rafique, S.; Idrees, M. Global expansion of chikungunya virus: Mapping the 64-year history. Int. J. Infect. Dis. 2017, 58, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Chu, J.J.H. Chikungunya virus: An update on antiviral development and challenges. Drug Discov. Today 2013, 18, 969–983. [Google Scholar] [CrossRef] [PubMed]
- Proesmans, S.; Katshongo, F.; Milambu, J.; Fungula, B.; Mavoko, H.M.; Ahuka-Mundeke, S.; Da Luz, R.I.; Van Esbroeck, M.; Ariën, K.K.; Cnops, L. Dengue and Chikungunya among Febrile Outpatients in Kinshasa, Democratic Republic of Congo: A cross-sectional study. PLoS Negl Trop Dis. 2019, 13, e0007047. [Google Scholar] [CrossRef]
- Bonn, D. How Did Chikungunya Reach Indian Ocean? Lancet Infect. Dis. 2006, 6, 543. [Google Scholar] [CrossRef]
- Hayes, E.B.; Sejvar, J.J.; Zaki, S.R.; Lanciotti, R.S.; Bode, A.V.; Campbell, G.L. Virology, pathology, and clinical manifestations of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1174. [Google Scholar] [CrossRef]
- Komar, N. West Nile viral encephalitis. Rev. Sci. Et Tech. Int. Off. Epizoot. 2000, 19, 166–176. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Campbell, S.R.; Alleyne, E.O.; Dobson, A.P.; Daszak, P. West Nile virus risk assessment and the bridge vector paradigm. Emerg. Infect. Dis. 2005, 11, 425. [Google Scholar] [CrossRef]
- Petersen, L.R.; Marfin, A.A. West Nile virus: A primer for the clinician. Ann. Intern. Med. 2002, 137, 173–179. [Google Scholar] [CrossRef]
- Chisenga, C.C.; Bosomprah, S.; Musukuma, K.; Mubanga, C.; Chilyabanyama, O.N.; Velu, R.M.; Kim, Y.C.; Reyes-Sandoval, A.; Chilengi, R. Sero-prevalence of arthropod-borne viral infections among Lukanga swamp residents in Zambia. PLoS ONE 2020, 15, e0235322. [Google Scholar] [CrossRef] [PubMed]
- Mweene-Ndumba, I.; Siziya, S.; Monze, M.; Mazaba, M.L.; Masaninga, F.; Songolo, P.; Mwaba, P.; Babaniyi, O.A. Seroprevalence of West Nile virus specific IgG and IgM antibodies in North-Western and Western provinces of Zambia. Afr. Health Sci. 2015, 15, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Rodger, L. An outbreak of suspected chikungunya fever in Nothern Rhodesia. S. Afr. Med. J. 1961, 35, 126–128. [Google Scholar] [PubMed]
- Mazaba-Liwewe, M.L.; Babaniyi, O.; Monza, M.; Mweene-Ndumba, I.; Mulenga, D.; Masaninga, F.; Songolo, P.; Kasolo, F.; Siziya, S. Dengue fever and factors associated with it in Western provinces of Zambia. Int. Public Health J. 2016, 8, 65. [Google Scholar]
- Babaniyi, O.; Mazaba-Liwewe, M.L.; Masaninga, F.; Mwaba, P.; Mulenga, D.; Songolo, P.; Mweene-Ndumba, I.; Rudatsikira, E.; Siziya, S. Prevalence of yellow fever in North-Western Province of Zambia. Int. J. Public Health Epidemiol. 2016, 8, 29–32. [Google Scholar]
- Orba, Y.; Hang’ombe, B.M.; Mweene, A.S.; Wada, Y.; Anindita, P.D.; Phongphaew, W.; Qiu, Y.; Kajihara, M.; Mori-Kajihara, A.; Eto, Y.; et al. First isolation of West Nile virus in Zambia from mosquitoes. Transbound. Emerg. Dis. 2018, 65, 434. [Google Scholar] [CrossRef]
- Simulundu, E.; Ndashe, K.; Chambaro, H.M.; Squarre, D.; Reilly, P.M.; Chitanga, S.; Changula, K.; Mukubesa, A.N.; Ndebe, J.; Tembo, J. West Nile Virus in Farmed Crocodiles, Zambia, 2019. Emerg. Infect. Dis. 2020, 26, 811. [Google Scholar] [CrossRef]
- Takaya, S.; Kutsuna, S.; Nakayama, E.; Taniguchi, S.; Tajima, S.; Katanami, Y.; Yamamoto, K.; Takeshita, N.; Hayakawa, K.; Kato, Y. Chikungunya fever in traveler from Angola to Japan, 2016. Emerg. Infect. Dis. 2017, 23, 156. [Google Scholar] [CrossRef]
- Hertz, J.T.; Munishi, O.M.; Ooi, E.E.; Howe, S.; Lim, W.Y.; Chow, A.; Morrissey, A.B.; Bartlett, J.A.; Onyango, J.J.; Maro, V.P. Chikungunya and dengue fever among hospitalized febrile patients in northern Tanzania. Am. J. Trop. Med. Hyg. 2012, 86, 171. [Google Scholar] [CrossRef]
- Townsend Peterson, A.; Papeş, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of GARP and Maxent. Ecography 2007, 30, 550–560. [Google Scholar] [CrossRef]
- Abdelkrim, O.; Samia, B.; Said, Z.; Souad, L. Modeling and mapping the habitat suitability and the potential distribution of Arboviruses vectors in Morocco. Parasite 2021, 28, 37. [Google Scholar] [CrossRef] [PubMed]
- Sofia, M.; Giannakopoulos, A.; Giantsis, I.A.; Touloudi, A.; Birtsas, P.; Papageorgiou, K.; Athanasakopoulou, Z.; Chatzopoulos, D.C.; Vrioni, G.; Galamatis, D. West Nile Virus Occurrence and Ecological Niche Modeling in Wild Bird Species and Mosquito Vectors: An Active Surveillance Program in the Peloponnese Region of Greece. Microorganisms 2022, 10, 1328. [Google Scholar] [CrossRef] [PubMed]
- León, B.; Jiménez-Sánchez, C.; Retamosa-Izaguirre, M. An environmental niche model to estimate the potential presence of Venezuelan equine encephalitis virus in Costa Rica. Int. J. Environ. Res. Public Health 2021, 18, 227. [Google Scholar] [CrossRef] [PubMed]
- Burch, C.; Loraamm, R.; Unnasch, T.; Downs, J. Utilizing ecological niche modelling to predict habitat suitability of eastern equine encephalitis in Florida. Ann. GIS 2020, 26, 133–147. [Google Scholar] [CrossRef]
- Libanda, B.; Bwalya, K.; Nkolola, N.B.; Chilekana, N. Quantifying long-term variability of precipitation and temperature over Zambia. J. Atmos. Sol. Terr. Phys. 2020, 198, 105201. [Google Scholar] [CrossRef]
- Dautu, G.; Mweene, A.S.; Samui, K.L.; Sindato, C.; Roy, P.; Noad, R.; Paweska, J.; Majiwa, P.A.; Musoke, A.J. Rift Valley fever: Real or perceived threat for Zambia? Onderstepoort J. Vet. Res. 2012, 79, 1–6. [Google Scholar] [CrossRef]
- Velu, R.M.; Kwenda, G.; Libonda, L.; Chisenga, C.C.; Flavien, B.N.; Chilyabanyama, O.N.; Simunyandi, M.; Bosomprah, S.; Sande, N.C.; Changula, K. Mosquito-Borne Viral Pathogens Detected in Zambia: A Systematic Review. Pathogens 2021, 10, 1007. [Google Scholar] [CrossRef]
- Kent, R.J. The Mosquitoes of Macha, Zambia; Johns Hopkins Malaria Research Institute: Baltimore, MD, USA, 2006. [Google Scholar]
- Chambaro, H.M.; Hirose, K.; Sasaki, M.; Libanda, B.; Sinkala, Y.; Fandamu, P.; Muleya, W.; Banda, F.; Chizimu, J.; Squarre, D. An unusually long Rift valley fever inter-epizootic period in Zambia: Evidence for enzootic virus circulation and risk for disease outbreak. PLoS Neglected Trop. Dis. 2022, 16, e0010420. [Google Scholar] [CrossRef]
- Masaninga, F.; Muleba, M.; Masendu, H.; Songolo, P.; Mweene-Ndumba, I.; Mazaba-Liwewe, M.L.; Kamuliwo, M.; Ameneshewa, B.; Siziya, S.; Babaniyi, O.A. Distribution of yellow fever vectors in Northwestern and Western Provinces, Zambia. Asian Pac. J. Trop. Med. 2014, 7, S88–S92. [Google Scholar] [CrossRef]
- Data, W.-G.C. Free Climate Data for Ecological Modeling and GIS. 2016. Available online: http://www.worldclim.org (accessed on 3 March 2023).
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Richman, R.; Diallo, D.; Diallo, M.; Sall, A.A.; Faye, O.; Diagne, C.T.; Dia, I.; Weaver, S.C.; Hanley, K.A.; Buenemann, M. Ecological niche modeling of Aedes mosquito vectors of chikungunya virus in southeastern Senegal. Parasites Vectors 2018, 11, 255. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, B.; Mondet, B.; Touzeau, S. Using a climate-dependent model to predict mosquito abundance: Application to Aedes (Stegomyia) africanus and Aedes (Diceromyia) furcifer (Diptera: Culicidae). Infect. Genet. Evol. 2008, 8, 422–432. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The effect of temperature on life history traits of Culex mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Brault, A.C. Changing patterns of West Nile virus transmission: Altered vector competence and host susceptibility. Veter. Res. 2009, 40, 43–49. [Google Scholar] [CrossRef]
- Nayar, J. Effects of constant and fluctuating temperatures on life span of Aedes taeniorhynchus adults. J. Insect Physiol. 1972, 18, 1303–1313. [Google Scholar] [CrossRef]
- Waldock, J.; Chandra, N.L.; Lelieveld, J.; Proestos, Y.; Michael, E.; Christophides, G.; Parham, P.E. The role of environmental variables on Aedes albopictus biology and chikungunya epidemiology. Pathog. Glob. Health 2013, 107, 224–241. [Google Scholar] [CrossRef]
- Gangoso, L.; Aragonés, D.; Martínez-de la Puente, J.; Lucientes, J.; Delacour-Estrella, S.; Peña, R.E.; Montalvo, T.; Bueno-Marí, R.; Bravo-Barriga, D.; Frontera, E. Determinants of the current and future distribution of the West Nile virus mosquito vector Culex pipiens in Spain. Environ. Res. 2020, 188, 109837. [Google Scholar] [CrossRef]
- Bravo-Barriga, D.; Gomes, B.; Almeida, A.P.; Serrano-Aguilera, F.J.; Pérez-Martín, J.E.; Calero-Bernal, R.; Reina, D.; Frontera, E.; Pinto, J. The mosquito fauna of the western region of Spain with emphasis on ecological factors and the characterization of Culex pipiens forms. J. Vector Ecol. 2017, 42, 136–147. [Google Scholar] [CrossRef]
- Minakawa, N.; Sonye, G.; Mogi, M.; Githeko, A.; Yan, G. The effects of climatic factors on the distribution and abundance of malaria vectors in Kenya. J. Med. Entomol. 2002, 39, 833–841. [Google Scholar] [CrossRef]
- Linton, Y.-M.; Dao, A.; Diallo, M.; Sanogo, Z.L.; Samake, D.; Ousmane, Y.; Kouam, C.; Krajacich, B.J.; Faiman, R.; Bamou, R. Diversity, composition, altitude, and seasonality of high-altitude windborne migrating mosquitoes in the Sahel: Implications for disease transmission. Front. Epidemiol. 2022, 2, 40. [Google Scholar]
- Afrane, Y.A.; Zhou, G.; Lawson, B.W.; Githeko, A.K.; Yan, G. Life-table analysis of Anopheles arabiensis in western Kenya highlands: Effects of land covers on larval and adult survivorship. Am. J. Trop. Med. Hyg. 2007, 77, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Brady, O.J.; Golding, N.; Pigott, D.M.; Kraemer, M.U.; Messina, J.P.; Reiner, R.C., Jr.; Scott, T.W.; Smith, D.L.; Gething, P.W.; Hay, S.I. Global temperature constraints on Aedes aegypti and Ae. albopictus persistence and competence for dengue virus transmission. Parasites Vectors 2014, 7, 338. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Escovar, J.; Moncada, L. Altitude and life tables of Culex quinquefasciatus (Diptera cucilidae) populations. Rev. Colomb. Entomol. 2010, 36, 62–67. [Google Scholar] [CrossRef]
- Pascual, M.; Ahumada, J.A.; Chaves, L.F.; Rodo, X.; Bouma, M. Malaria resurgence in the East African highlands: Temperature trends revisited. Proc. Natl. Acad. Sci. USA 2006, 103, 5829–5834. [Google Scholar] [CrossRef]
- Mubemba, B.; Mburu, M.M.; Changula, K.; Muleya, W.; Moonga, L.C.; Chambaro, H.M.; Kajihara, M.; Qiu, Y.; Orba, Y.; Hayashida, K. Current knowledge of vector-borne zoonotic pathogens in Zambia: A clarion call to scaling-up “One Health” research in the wake of emerging and re-emerging infectious diseases. PLoS Neglected Trop. Dis. 2022, 16, e0010193. [Google Scholar] [CrossRef]
- Pearson, R.; Dawson, T.; Liu, C.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species’ distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Description | Abbreviation | Unit |
|---|---|---|
| Temperature seasonality (standard deviation × 100) | BIO4 | °C |
| Mean temperature of wettest quarter | BIO8 | °C |
| Mean temperature of driest quarter | BIO9 | °C |
| Precipitation seasonality (coefficient of variation) | BIO15 | Mm |
| Precipitation of wettest quarter | BIO16 | Mm |
| Precipitation of driest quarter | BIO17 | Mm |
| Precipitation of warmest quarter | BIO18 | Mm |
| Precipitation of coldest quarter | BIO19 | Mm |
| Elevation | N/A | M |
| Culex Model | Aedes Model | ||
|---|---|---|---|
| Environmental Variable | Contribution (%) | Environmental Variable | Contribution (%) |
| Precipitation of warmest quarter (BIO18) | 41.2 | Precipitation of warmest quarter (BIO18) | 85 |
| Precipitation of driest quarter (BIO17) | 37.7 | Precipitation of driest quarter (BIO17) | 14.3 |
| Temperature seasonality (standard deviation × 100; BIO4) | 11.7 | Precipitation of coldest quarter (BIO19) | 0.4 |
| Precipitation of coldest quarter (BIO19) | 3.7 | Temperature seasonality (standard deviation × 100; BIO4) | 0.2 |
| Precipitation of wettest quarter (BIO16) | 3 | Mean temperature of driest quarter (BIO9) | 0 |
| Elevation | 1.2 | Precipitation of wettest quarter (BIO16) | 0 |
| Precipitation seasonality (coefficient of variation; BIO15) | 1 | Elevation | 0 |
| Mean temperature of driest quarter (BIO9) | 0.4 | Mean temperature of driest quarter (BIO8) | 0 |
| Mean temperature of driest quarter (BIO8) | 0.1 | Precipitation seasonality (coefficient of variation; BIO15) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velu, R.M.; Kwenda, G.; Bosomprah, S.; Chisola, M.N.; Simunyandi, M.; Chisenga, C.C.; Bumbangi, F.N.; Sande, N.C.; Simubali, L.; Mburu, M.M.; et al. Ecological Niche Modeling of Aedes and Culex Mosquitoes: A Risk Map for Chikungunya and West Nile Viruses in Zambia. Viruses 2023, 15, 1900. https://doi.org/10.3390/v15091900
Velu RM, Kwenda G, Bosomprah S, Chisola MN, Simunyandi M, Chisenga CC, Bumbangi FN, Sande NC, Simubali L, Mburu MM, et al. Ecological Niche Modeling of Aedes and Culex Mosquitoes: A Risk Map for Chikungunya and West Nile Viruses in Zambia. Viruses. 2023; 15(9):1900. https://doi.org/10.3390/v15091900
Chicago/Turabian StyleVelu, Rachel Milomba, Geoffrey Kwenda, Samuel Bosomprah, Moses Ngongo Chisola, Michelo Simunyandi, Caroline Cleopatra Chisenga, Flavien Nsoni Bumbangi, Nicholus Chintu Sande, Limonty Simubali, Monicah Mirai Mburu, and et al. 2023. "Ecological Niche Modeling of Aedes and Culex Mosquitoes: A Risk Map for Chikungunya and West Nile Viruses in Zambia" Viruses 15, no. 9: 1900. https://doi.org/10.3390/v15091900