
Seroprevalence of IgG Antibodies Directed against Dengue, Chikungunya and West Nile Viruses and Associated Risk Factors in Madagascar, 2011 to 2013
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Laboratory Analysis
2.3. Environmental Data Collection
2.4. Multiple Factors Analysis (MFA)
2.5. Logistical Regression Analysis
3. Results
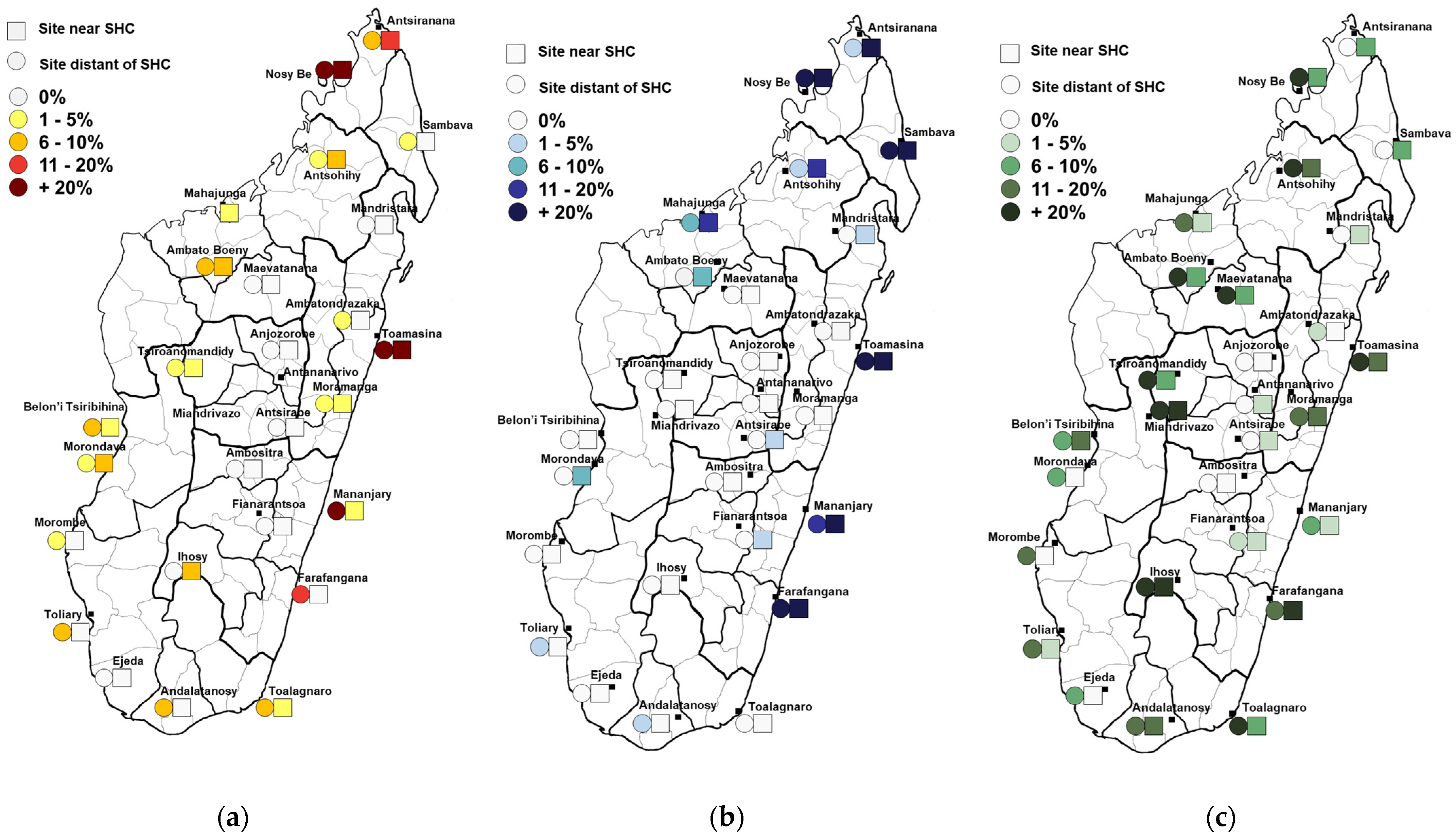
3.1. Seroprevalence Results
3.1.1. Dengue Virus
3.1.2. Chikungunya Virus
3.1.3. West Nile Virus
3.2. Logistical Regression Results
3.2.1. Multiple Factor Analysis for Environmental Variables
- Factor 1 positivity represents ecosystems with high day and night temperatures, with low precipitation and NDVI, associated with the absence of wooded vegetation.
- Factor 2 positivity represents ecosystems with high NDVI and relatively high night temperatures, without grassland and moderate levels of cultivated areas, but with the important presence of humid forests and wetlands.
- Factor 3 positivity represents ecosystems with high day and night temperatures, with moderate precipitation dominated by the presence of cultivated areas but no bare soil or water bodies.
- Factor 4 positivity represents areas with moderate temperatures and NDVI but with a low surface of water, moderate levels of forests, and the important presence of bare soil.
3.2.2. Multivariate Regression Models
4. Discussion
4.1. Dengue and Chikungunya Viruses
4.2. West Nile Virus
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fontenille, D.; Mathiot, C.; Rodhain, F.; Maleyran, D.; Rakotoarivony, I.; Digoutte, J.P.; Coulanges, P. Arbovirus diseases in the region of Tsiroanomandidy, Madagascar. Entomological, virological and serological studies. Ann. Soc. Belg. Med. Trop 1988, 68, 43–52. [Google Scholar] [PubMed]
- Ratsitorahina, M.; Harisoa, J.; Ratovonjato, J.; Biacabe, S.; Reynes, J.-M.; Zeller, H.; Raoelina, Y.; Talarmin, A.; Richard, V.; Louis Soares, J. Outbreak of Dengue and Chikungunya Fevers, Toamasina, Madagascar, 2006. Emerg. Infect. Dis. 2008, 14, 1135–1137. [Google Scholar] [CrossRef]
- Guzman, M.G.; Halstead, S.B.; Artsob, H.; Buchy, P.; Farrar, J.; Gubler, D.J.; Hunsperger, E.; Kroeger, A.; Margolis, H.S.; Martínez, E.; et al. Dengue: A Continuing Global Threat. Nat. Rev. Microbiol. 2010, 8, S7–S16. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The Global Distribution and Burden of Dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Gray, T.J.; Webb, C.E. A Review of the Epidemiological and Clinical Aspects of West Nile Virus. Int. J. Gen. Med. 2014, 7, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Renault, P.; Balleydier, E.; D’Ortenzio, E.; Bâville, M.; Filleul, L. Epidemiology of Chikungunya Infection on Reunion Island, Mayotte, and Neighboring Countries. Médecine Mal. Infect. 2012, 42, 93–101. [Google Scholar] [CrossRef]
- Sissoko, D.; Ezzedine, K.; Moendandzé, A.; Giry, C.; Renault, P.; Malvy, D. Field Evaluation of Clinical Features during Chikungunya Outbreak in Mayotte, 2005-2006. Trop. Med. Int. Health 2010, 15, 600–607. [Google Scholar] [CrossRef]
- Furuya-Kanamori, L.; Liang, S.; Milinovich, G.; Soares Magalhaes, R.J.; Clements, A.C.A.; Hu, W.; Brasil, P.; Frentiu, F.D.; Dunning, R.; Yakob, L. Co-Distribution and Co-Infection of Chikungunya and Dengue Viruses. BMC Infect. Dis. 2016, 16, 84. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, N.G.; Girmann, M.; Randriamampionona, N.; Bialonski, A.; Maus, D.; Krefis, A.C.; Njarasoa, C.; Rajanalison, J.F.; Ramandrisoa, H.D.; Randriarison, M.L.; et al. Seroprevalence of Antibodies against Chikungunya, Dengue, and Rift Valley Fever Viruses after Febrile Illness Outbreak, Madagascar. Emerg. Infect. Dis. 2012, 18, 1780–1786. [Google Scholar] [CrossRef] [Green Version]
- Randrianasolo, L.; Raoelina, Y.; Ratsitorahina, M.; Ravolomanana, L.; Andriamandimby, S.; Heraud, J.-M.; Rakotomanana, F.; Ramanjato, R.; Randrianarivo-Solofoniaina, A.E.; Richard, V. Sentinel Surveillance System for Early Outbreak Detection in Madagascar. BMC Public Health 2010, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Tantely, M.L.; Goodman, S.M.; Rakotondranaivo, T.; Boyer, S. Review of West Nile Virus Circulation and Outbreak Risk in Madagascar: Entomological and Ornithological Perspectives. Parasite 2016, 23, 49. [Google Scholar] [CrossRef] [Green Version]
- Tantely, L.M.; Cêtre-Sossah, C.; Rakotondranaivo, T.; Cardinale, E.; Boyer, S. Population Dynamics of Mosquito Species in a West Nile Virus Endemic Area in Madagascar. Parasite 2017, 24, 3. [Google Scholar] [CrossRef] [Green Version]
- Morvan, J.; Chin, L.H.; Fontenille, D.; Rakotoarivony, I.; Coulanges, P. Prevalence of antibodies to West Nile virus in youngsters from 5 to 20 years old in Madagascar. Bull. Soc. Pathol. Exot. 1991, 84, 225–234. [Google Scholar]
- Lonchampt, C.; Migliani, R.; Ratsitorahina, M.; Rabarijaona, L.P.; Ramarokoto, C.E.; Rakoto Andrianarivelo, M.; Rousset, D. Persistence of an endemic circulation of the West Nile virus in Madagascar. Arch. Inst. Pasteur Madag. 2003, 69, 33–36. [Google Scholar]
- Olive, M.-M.; Chevalier, V.; Grosbois, V.; Tran, A.; Andriamandimby, S.-F.; Durand, B.; Ravalohery, J.-P.; Andriamamonjy, S.; Rakotomanana, F.; Rogier, C.; et al. Integrated Analysis of Environment, Cattle and Human Serological Data: Risks and Mechanisms of Transmission of Rift Valley Fever in Madagascar. PLoS Negl. Trop. Dis. 2016, 10, e0004827. [Google Scholar] [CrossRef] [Green Version]
- Westgard, J.O.; Barry, P.L.; Hunt, M.R.; Groth, T. A Multi-Rule Shewhart Chart for Quality Control in Clinical Chemistry. Clin. Chem. 1981, 27, 493–501. [Google Scholar] [CrossRef]
- Clarke, D.H.; Casals, J. Techniques for Hemagglutination and Hemagglutination-Inhibition with Arthropod-Borne Viruses. Am. J. Trop. Med. Hyg. 1958, 7, 561–573. [Google Scholar] [CrossRef] [Green Version]
- Moat, J.; Smith, P. Atlas of the vegetation of Madagascar. In Royal Botanic Gardens, Kew; Kew Publishing: London, UK, 2007. [Google Scholar]
- Cornet, A. Essai de cartographie bioclimatique à Madagascar. Not. Explic. De L’orstom 1974, 55, 1–28. [Google Scholar]
- Campbell, K.M.; Haldeman, K.; Lehnig, C.; Munayco, C.V.; Halsey, E.S.; Laguna-Torres, V.A.; Yagui, M.; Morrison, A.C.; Lin, C.-D.; Scott, T.W. Weather Regulates Location, Timing, and Intensity of Dengue Virus Transmission between Humans and Mosquitoes. PLoS Negl. Trop. Dis. 2015, 9, e0003957. [Google Scholar] [CrossRef]
- Fontenille, D. Arbovirus transmission cycles in Madagascar. Arch. Inst. Pasteur Madag. 1989, 55, 1–317. [Google Scholar]
- Raharimalala, F.N.; Ravaomanarivo, L.H.; Ravelonandro, P.; Rafarasoa, L.S.; Zouache, K.; Tran-Van, V.; Mousson, L.; Failloux, A.-B.; Hellard, E.; Moro, C.V.; et al. Biogeography of the Two Major Arbovirus Mosquito Vectors, Aedes aegypti and Aedes albopictus (Diptera, Culicidae), in Madagascar. Parasit Vectors 2012, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Mbanzulu, K.M.; Wumba, R.; Mboera, L.E.G.; Kayembe, J.-M.N.; Engbu, D.; Bojabwa, M.M.; Zanga, J.K.; Misinzo, G.; Kimera, S.I. Pattern of Aedes aegypti and Aedes albopictus Associated with Human Exposure to Dengue Virus in Kinshasa, the Democratic Republic of the Congo. Trop. Med. 2022, 7, 392. [Google Scholar] [CrossRef]
- Ravaonjanahary, C. Les Aedes de Madagascar (Diptera-Culicidae). Étude monographique du genre. 2. Biologie d’Aedes (Diceromyia) tiptoni. Trav. Doc. L’orstom 1978, 87, 1–210. [Google Scholar]
- Crepeau, T.N.; Healy, S.P.; Bartlett-Healy, K.; Unlu, I.; Farajollahi, A.; Fonseca, D.M. Effects of Biogents Sentinel Trap Field Placement on Capture Rates of Adult Asian Tiger Mosquitoes, Aedes Albopictus. PLoS ONE 2013, 8, e60524. [Google Scholar] [CrossRef]
- Samuel, M.; Maoz, D.; Manrique, P.; Ward, T.; Runge-Ranzinger, S.; Toledo, J.; Boyce, R.; Horstick, O. Community Effectiveness of Indoor Spraying as a Dengue Vector Control Method: A Systematic Review. PLoS Negl. Trop. Dis. 2017, 11, e0005837. [Google Scholar] [CrossRef]
- Boonyuan, W.; Kongmee, M.; Bangs, M.J.; Prabaripai, A.; Chareonviriyaphap, T. Host Feeding Responses of Aedes aegypti (L.) Exposed to Deltamethrin. J. Vector Ecol. 2011, 36, 361–372. [Google Scholar] [CrossRef]
- Obembe, A.; Popoola, K.O.K.; Oduola, A.O.; Awolola, S.T. Differential Behaviour of Endophilic Anopheles Mosquitoes in Rooms Occupied by Tobacco Smokers and Non-Smokers in Two Nigerian Villages. J. Appl. Sci. Environ. Manag. 2018, 22, 981. [Google Scholar] [CrossRef] [Green Version]
- Steppuhn, A.; Gase, K.; Krock, B.; Halitschke, R.; Baldwin, I.T. Nicotine’s Defensive Function in Nature. PLoS Biol. 2004, 2, E217. [Google Scholar] [CrossRef]
- Wong, L.P.; AbuBakar, S.; Chinna, K. Community Knowledge, Health Beliefs, Practices and Experiences Related to Dengue Fever and Its Association with IgG Seropositivity. PLoS Negl. Trop. Dis. 2014, 8, e2789. [Google Scholar] [CrossRef] [Green Version]
- Manore, C.A.; Hickmann, K.S.; Xu, S.; Wearing, H.J.; Hyman, J.M. Comparing Dengue and Chikungunya Emergence and Endemic Transmission in A. Aegypti and A. Albopictus. J. Theor. Biol. 2014, 356, 174–191. [Google Scholar] [CrossRef] [Green Version]
- Institut Pasteur de Madagascar. Rapport Annuel 2021, Rapport Annuel de l’Institut Pasteur de Madagascar, 2022. Available online: https://www.pasteur.mg/wp-content/uploads/2022/09/Rapport-dactivites-IPM_2021.pdf (accessed on 20 June 2023).
- Murray, K.O.; Garcia, M.N.; Yan, C.; Gorchakov, R. Persistence of Detectable Immunoglobulin M Antibodies up to 8 Years after Infection with West Nile Virus. Am. J. Trop. Med. Hyg. 2013, 89, 996–1000. [Google Scholar] [CrossRef] [Green Version]
- Tantely, M.L.; Guis, H.; Randriananjantenaina, I.; Raharinirina, M.R.; Velonirina, H.J.; Cardinale, E.; Raveloarijaona, N.; Cêtre-Sossah, C.; Garros, C.; Girod, R. Mosquito Species Associated with Horses in Madagasacr: A Review of Their Vector Status with Regard to the Epidemiology of West Nile Fever. Med. Vet. Entomol. 2022, 36, 1–13. [Google Scholar] [CrossRef]
- Abdullahi, I.N.; Emeribe, A.U.; Ghamba, P.E.; Omosigho, P.O.; Bello, Z.M.; Oderinde, B.S.; Fasogbon, S.A.; Olayemi, L.; Daneji, I.M.; Musa, M.H.; et al. Distribution Pattern and Prevalence of West Nile Virus Infection in Nigeria from 1950 to 2020: A Systematic Review. Epidemiol. Health 2020, 42, e2020071. [Google Scholar] [CrossRef]
- DeGroote, J.P.; Sugumaran, R. National and Regional Associations Between Human West Nile Virus Incidence and Demographic, Landscape, and Land Use Conditions in the Coterminous United States. Vector Borne Zoonotic Dis. 2012, 12, 657–665. [Google Scholar] [CrossRef]


{kind=link}
| City | Dengue Virus | Chikungunya Virus | West Nile Virus | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Nb Positive | (%) | CI * | Nb Positive | (%) | CI * | Nb Positive | (%) | CI * | |
| Ambato Boeny | 4 | 6.7 | 1.9–11.4 | 1 | 1.7 | 0.0–4.0 | 10 | 16.7 | 7.1–26.2 |
| Ambatondrazaka | 1 | 1.7 | 0.0–4.0 | 0 | 0.0 | 0.0–0.1 | 1 | 1.7 | 0.0–4.0 |
| Ambositra | 0 | 0.0 | 0.0–0.1 | 0 | 0.0 | 0.0–0.1 | 0 | 0.0 | 0.0–0.1 |
| Andalatanosy | 2 | 3.3 | 0.0–8.1 | 1 | 1.7 | 0.0–4.0 | 11 | 18.3 | 16.0–20.7 |
| Anjozorobe | 0 | 0.0 | 0.0–0.1 | 0 | 0.0 | 0.0–0.1 | 0 | 0.0 | 0.0–0.1 |
| Antsohihy | 4 | 6.7 | 1.9–11.4 | 7 | 11.7 | 0.0–23.6 | 18 | 30.0 | 6.2–53.8 |
| Antananarivo | 1 | 1.7 | 0.0–4.0 | 0 | 0.0 | 0.0–0.1 | 1 | 1.7 | 0.0–4.0 |
| Antsirabe | 0 | 0.0 | 0.0–0.1 | 1 | 1.7 | 0.0–4.0 | 1 | 1.7 | 0.0–4.0 |
| Antsiranana | 7 | 11.7 | 4.5–18.8 | 8 | 13.3 | 0.0–27.6 | 3 | 5.0 | 0.0–12.1 |
| Belon’i Tsiribihina | 3 | 5.0 | 0.0–12.1 | 0 | 0.0 | 0.0–0.1 | 6 | 10.0 | 5.2–14.8 |
| Ejeda | 0 | 0.0 | 0.0–0.1 | 0 | 0.0 | 0.0–0.1 | 2 | 3.3 | 0.0–8.1 |
| Farafangana | 4 | 6.7 | 0.0–16.2 | 37 | 61.7 | 59.3–64.0 | 15 | 25.0 | 17.9–32.1 |
| Fianarantsoa | 0 | 0.0 | 0.0–0.1 | 1 | 1.7 | 0.0–4.0 | 2 | 3.3 | 0.0–8.1 |
| Ihosy | 3 | 5.0 | 0.0–12.1 | 0 | 0.0 | 0.0–0.1 | 20 | 33.3 | 23.8–42.9 |
| Maevatanana | 0 | 0.0 | 0.0–0.1 | 0 | 0.0 | 0.0–0.1 | 11 | 18.3 | 6.4–30.2 |
| Mahajanga | 1 | 1.7 | 0.0–4.0 | 8 | 13.3 | 3.8–22.9 | 6 | 10.0 | 5.2–14.8 |
| Mananjary | 9 | 15.0 | 0.0–31.7 | 22 | 36.7 | 8.1–65.3 | 4 | 6.7 | 1.9–11.4 |
| Mandritsara | 0 | 0.0 | 0.0–0.1 | 1 | 1.7 | 0.0–4.0 | 2 | 3.3 | 0.0–8.1 |
| Miandrivazo | 1 | 1.7 | 0.0–4.0 | 0 | 0.0 | 0.0–0.1 | 15 | 25.0 | 22.6–27.4 |
| Moramanga | 2 | 3.3 | 0.0–8.1 | 0 | 0.0 | 0.0–0.1 | 9 | 15.0 | 12.6–17.4 |
| Morombe | 1 | 1.7 | 0.0–4.0 | 0 | 0.0 | 0.0–0.1 | 5 | 8.3 | 0.0–20.2 |
| Morondava | 3 | 5.0 | 2.6–7.4 | 2 | 3.3 | 0.0–8.1 | 3 | 5.0 | 0.0–12.1 |
| Nosy Be | 29 | 48.3 | 26.9–69.8 | 52 | 86.7 | 81.9–91.4 | 14 | 23.3 | 4.2–42.4 |
| Sambava | 1 | 1.7 | 0.0–4.0 | 46 | 76.7 | 52.8–100.0 | 2 | 3.3 | 0.0–8.1 |
| Taolagnaro | 4 | 6.7 | 1.9–11.4 | 0 | 0.0 | 0.0–0.1 | 11 | 18.3 | 1.6–35.0 |
| Toamasina | 26 | 43.3 | 30.4–56.2 | 42 | 70.0 | 0.0–0.1 | 27 | 45.0 | 0.5–19.5 |
| Toliary | 2 | 3.3 | 0.0–8.1 | 1 | 1.7 | 0.0–4.0 | 6 | 10.0 | 5.2–14.8 |
| Tsiroanomandidy | 2 | 3.3 | 0.0–8.1 | 0 | 0.0 | 0.0–0.1 | 9 | 15.0 | 3.1–26.9 |
| TOTAL | 110 | 6.5 | 3.2–9.9 | 230 | 13.7 | 6.5–20.9 | 214 | 12.7 | 9.0–16.5 |
| Covariates | Factor 1 | Factor 2 | Factor 3 | Factor 4 |
|---|---|---|---|---|
| Land Surface Temperature Night (LSTN) | 0.6359734 | 0.4834523 | 0.3790634 | 0.1336330 |
| Land Surface Temperature Day (LSTD) | 0.6743499 | / | 0.5535304 | / |
| Normalized Difference Vegetation Index (NDVI) | −0.6806744 | 0.4492561 | / | 0.1418775 |
| Precipitations | −0.2008070 | −0.3733040 | 0.2891146 | 0.2345488 |
| Land Cover Grassland | / | −1.2587948 | / | / |
| Land Cover Water/wetlands | 0.6192618 | 0.8190209 | −0.4854862 | −0.7919986 |
| Land Cover Bare soil | 0.9902471 | 0.4585246 | −0.5887855 | 0.8189414 |
| Land Cover Cultivated areas | −0.3448483 | 0.3614344 | 0.6748149 | −0.1676591 |
| Land cover Humid Forest | −1.3089895 | 0.9065925 | / | 0.2996699 |
| Virus | Tested Variable | N Tested 1 | Nb Pos (%) | IC Pos | Adjusted OR | IC 95% OR | p-Value |
|---|---|---|---|---|---|---|---|
| Dengue | MFA Factor 2 | 1678 | 1.66 | 1.24–2.18 | 0.001 | ||
| MFA Factor 4 | 1678 | 1.88 | 1.30–2.72 | 0.001 | |||
| Smoker | 0.031 | ||||||
| No | 1334 | 93 (7.0) | 4.8–9.1 | 1 | |||
| Yes | 344 | 17 (4.9) | 2.5–7.4 | 0.48 | 0.26–0.89 | ||
| Frequent work in rice fields | <0.001 | ||||||
| No | 909 | 77 (8.5) | 5.5–11.4 | 1 | |||
| Yes | 769 | 33 (4.3) | 2.6–6.0 | 0.36 | 0.21–0.63 | ||
| Frequent activities in the forest | 0.048 | ||||||
| No | 1083 | 58 (5.4) | 3.4–7.3 | 1 | |||
| Yes | 595 | 52 (8.7) | 5.4–12.1 | 1.72 | 1.00–2.94 | ||
| House targeted for anti-mosquito spraying program (last 12 months) | 0.002 | ||||||
| No | 1190 | 103 (8.7) | 6.1–11.3 | 1 | |||
| Yes | 488 | 7 (1.4) | 0.3–2.6 | 0.24 | 0.10–0.59 | ||
| Chikungunya | MFA Factor 2 | 1678 | 2.48 | 1.37–4.46 | 0.003 | ||
| General environment | 0.031 | ||||||
| Urban | 516 | 105 (20.3) | 13.2–27.5 | 1 | 0.008 | ||
| Peri-urban | 450 | 21 (4.7) | 1.4–7.9 | 0.14 | 0.04–0.51 | 0.003 | |
| Rural | 714 | 104 (14.6) | 6.8–22.3 | 0.25 | 0.06–0.99 | 0.048 | |
| Frequent work in rice fields | 0.018 | ||||||
| No | 911 | 162 (17.8) | 12.8–22.7 | 1 | |||
| Yes | 769 | 68 (8.8) | 3.6–14.1 | 0.42 | 0.21–0.86 | ||
| Frequent activities in the forest | 0.015 | ||||||
| No | 1084 | 133 (12.2) | 8.5–16.1 | 1 | |||
| Yes | 596 | 97 (16.3) | 8.9–23.7 | 2.27 | 1.17–4.39 | ||
| Non-biological wastes next to the house | 0.041 | ||||||
| No | 457 | 25 (5.5) | 2.8–8.1 | 1 | |||
| Yes | 1221 | 205 (16.8) | 11.6–21.9 | 2.04 | 1.03–4.03 | ||
| House targeted for anti-mosquito spraying program (last 12 months) | <0.001 | ||||||
| No | 1190 | 228 (19.2) | 13.7–24.6 | 1 | |||
| Yes | 488 | 2 (0.01) | 0.00–0.01 | 0.02 | 0.00–0.18 | ||
| West Nile | MFA Factor 2 | 1678 | 1.37 | 1.08–1.75 | 0.010 | ||
| MFA Factor 3 | 1678 | 1.48 | 1.11–1.97 | 0.007 | |||
| Frequent contacts to water bodies (fishing, swimming) | 0.033 | ||||||
| No | 1564 | 195 (12.5) | 10.0–14.9 | 1 | |||
| Yes | 116 | 19 (16.4) | 8.1–24.6 | 2.07 | 1.06–3.84 | ||
| Electricity in the house | 0.001 | ||||||
| No | 1114 | 180 (16.2) | 12.8–19.5 | 1 | |||
| Yes | 564 | 34 (6.0) | 3.7–8.3 | 0.42 | 0.25–0.70 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broban, A.; Olive, M.-M.; Tantely, M.L.; Dorsemans, A.-C.; Rakotomanana, F.; Ravalohery, J.-P.; Rogier, C.; Heraud, J.-M.; Andriamandimby, S.F. Seroprevalence of IgG Antibodies Directed against Dengue, Chikungunya and West Nile Viruses and Associated Risk Factors in Madagascar, 2011 to 2013. Viruses 2023, 15, 1707. https://doi.org/10.3390/v15081707
Broban A, Olive M-M, Tantely ML, Dorsemans A-C, Rakotomanana F, Ravalohery J-P, Rogier C, Heraud J-M, Andriamandimby SF. Seroprevalence of IgG Antibodies Directed against Dengue, Chikungunya and West Nile Viruses and Associated Risk Factors in Madagascar, 2011 to 2013. Viruses. 2023; 15(8):1707. https://doi.org/10.3390/v15081707
Chicago/Turabian StyleBroban, Anaïs, Marie-Marie Olive, Michael Luciano Tantely, Anne-Claire Dorsemans, Fanjasoa Rakotomanana, Jean-Pierre Ravalohery, Christophe Rogier, Jean-Michel Heraud, and Soa Fy Andriamandimby. 2023. "Seroprevalence of IgG Antibodies Directed against Dengue, Chikungunya and West Nile Viruses and Associated Risk Factors in Madagascar, 2011 to 2013" Viruses 15, no. 8: 1707. https://doi.org/10.3390/v15081707