
Lichen or Associated Micro-Organism Compounds Are Active against Human Coronaviruses

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Natural Lichen Products
2.3. Cells and Culture Conditions
2.4. Viruses
2.5. Cell Toxicity Assay
2.6. HCoV-229E Infection Inhibition Assays
2.6.1. Luciferase Assay
2.6.2. Time-of-Addition Assay
2.7. SARS-CoV-2 Infection Inhibition Assays
2.8. Statistical Analysis
3. Results
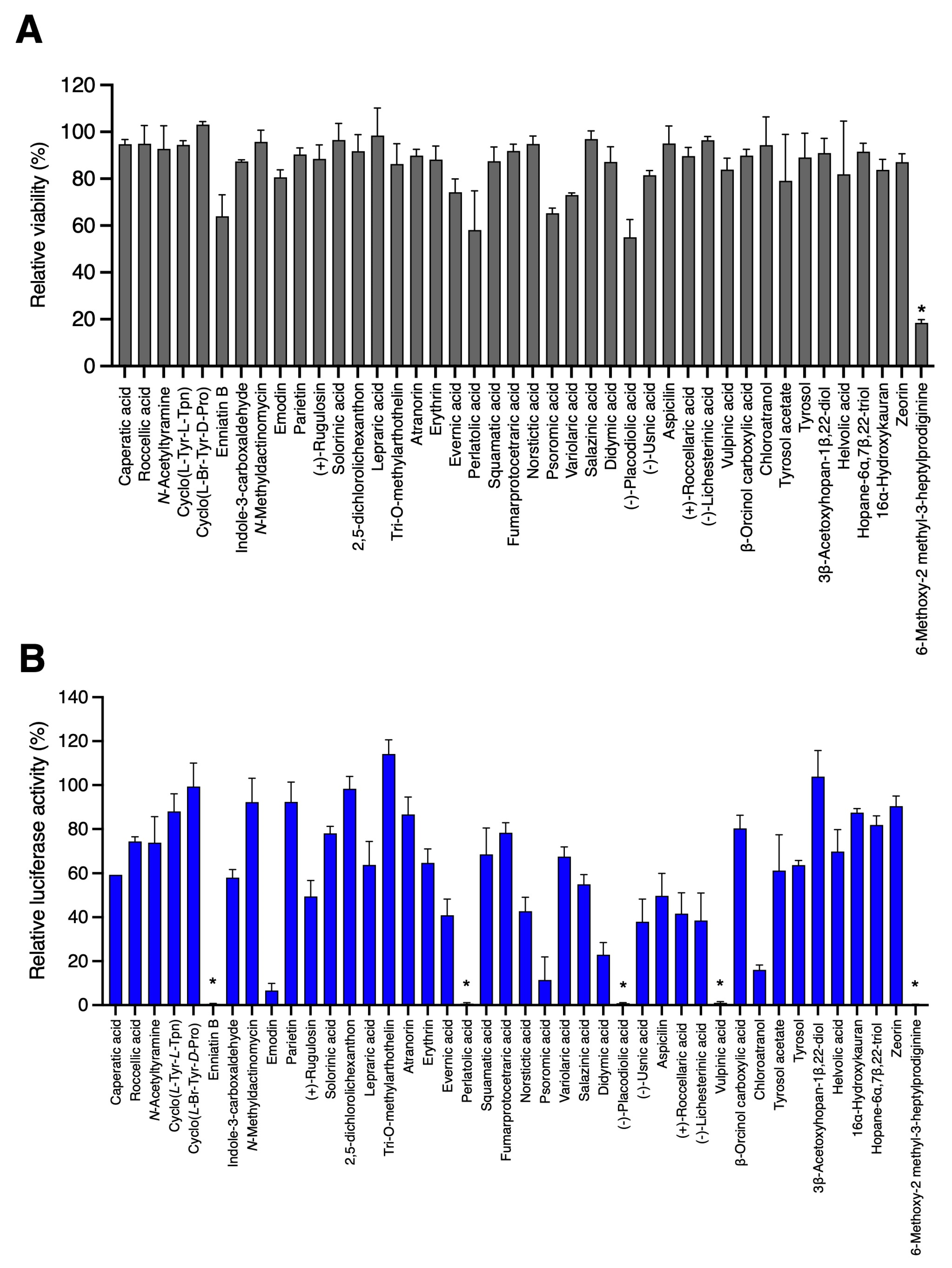
3.1. Screening of 42 Compounds Isolated from Lichens
3.2. Determination of IC50 of the Active Compounds
3.3. Kinetic Study
3.4. Anti-SARS-CoV-2 Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular evolution of Human coronavirus genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2020, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Malone, B.; Urakova, N.; Snijder, E.J.; Campbell, E.A. Structures and functions of coronavirus replication-transcription complexes and their relevance for SARS-CoV-2 drug design. Nat. Rev. Mol. Cell Biol. 2022, 23, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on Ace2 and TMPRSS2 and Is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zheng, Q.; Sun, L.; Ji, M.; Li, Y.; Deng, H.; Zhang, H. ORF3a of SARS-CoV-2 promotes lysosomal exocytosis-mediated viral egress. Dev. Cell 2021, 56, 3250–3263.e5. [Google Scholar] [CrossRef]
- Ghosh, S.; Dellibovi-Ragheb, T.A.; Pak, E.; Qiu, Q.; Fisher, M.; Takvorian, P.M.; Bleck, C.; Hsu, V.; Fehr, A.R.; Perlman, S.; et al. β-Coronaviruses use lysosomal organelles for cellular egress. bioRxiv 2020, 7, 192310. [Google Scholar] [CrossRef]
- Bruel, T.; Hadjadj, J.; Maes, P.; Planas, D.; Seve, A.; Staropoli, I.; Guivel-Benhassine, F.; Porrot, F.; Bolland, W.-H.; Nguyen, Y.; et al. Serum neutralization of SARS-CoV-2 Omicron sublineages BA.1 and BA.2 in patients receiving monoclonal antibodies. Nat. Med. 2022, 28, 1297–1302. [Google Scholar] [CrossRef]
- Takashita, E.; Yamayoshi, S.; Simon, V.; van Bakel, H.; Sordillo, E.M.; Pekosz, A.; Fukushi, S.; Suzuki, T.; Maeda, K.; Halfmann, P.; et al. Efficacy of Antibodies and Antiviral Drugs against Omicron BA.2.12.1, BA.4, and BA.5 Subvariants. N. Engl. J. Med. 2022, 387, 468–470. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M.; et al. Early Remdesivir to Prevent Progression to Severe COVID-19 in Outpatients. N. Engl. J. Med. 2022, 386, 305–315. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Leist, S.R.; Schäfer, A.; Dinnon, K.H.; Stevens, L.J.; et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Transl. Med. 2020, 12, eabb5883. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.; Hobbs, R.; Gbinigie, O.; Rahman, N.M.; Hayward, G.; Richards, D.; Dorward, J.; Lowe, D.; Standing, J.F.; Breuer, J.; et al. Molnupiravir Plus Usual Care Versus Usual Care Alone as Early Treatment for Adults with COVID-19 at Increased Risk of Adverse Outcomes (PANORAMIC): Preliminary Analysis from the United Kingdom Randomised, Controlled Open-Label, Platform Adaptive Trial. Lancet 2022, 401, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.R.; Allerton, C.M.N.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J.; et al. An oral SARS-CoV-2 Mpro inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 374, 1586–1593. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.; Leister-Tebbe, H.; Gardner, A.; Abreu, P.; Bao, W.; Wisemandle, W.; Baniecki, M.; Hendrick, V.M.; Damle, B.; Simón-Campos, A.; et al. Oral Nirmatrelvir for High-Risk, Nonhospitalized Adults with COVID-19. N. Engl. J. Med. 2022, 386, 1397–1408. [Google Scholar] [CrossRef]
- Ali, S.I.; Sheikh, W.M.; Rather, M.A.; Venkatesalu, V.; Muzamil Bashir, S.; Nabi, S.U. Medicinal plants: Treasure for antiviral drug discovery. Phytother. Res. 2021, 35, 3447–3483. [Google Scholar] [CrossRef]
- Vougogiannopoulou, K.; Corona, A.; Tramontano, E.; Alexis, M.N.; Skaltsounis, A.-L. Natural and Nature-Derived Products Targeting Human Coronaviruses. Molecules 2021, 26, 448. [Google Scholar] [CrossRef]
- Christy, M.P.; Uekusa, Y.; Gerwick, L.; Gerwick, W.H. Natural Products with Potential to Treat RNA Virus Pathogens Including SARS-CoV-2. J. Nat. Prod. 2021, 84, 161–182. [Google Scholar] [CrossRef]
- Müller, C.; Schulte, F.W.; Lange-Grünweller, K.; Obermann, W.; Madhugiri, R.; Pleschka, S.; Ziebuhr, J.; Hartmann, R.K.; Grünweller, A. Broad-spectrum antiviral activity of the eIF4A inhibitor silvestrol against corona- and picornaviruses. Antivir. Res. 2018, 150, 123–129. [Google Scholar] [CrossRef]
- Kim, D.E.; Min, J.S.; Jang, M.S.; Lee, J.Y.; Shin, Y.S.; Song, J.H.; Kim, H.R.; Kim, S.; Jin, Y.-H.; Kwon, S. Natural Bis-Benzylisoquinoline Alkaloids-Tetrandrine, Fangchinoline, and Cepharanthine, Inhibit Human Coronavirus OC43 Infection of MRC-5 Human Lung Cells. Biomolecules 2019, 9, 696. [Google Scholar] [CrossRef]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef]
- Meunier, T.; Desmarets, L.; Bordage, S.; Bamba, M.; Hervouet, K.; Rouillé, Y.; François, N.; Decossas, M.; Sencio, V.; Trottein, F.; et al. A Photoactivable Natural Product with Broad Antiviral Activity against Enveloped Viruses, Including Highly Pathogenic Coronaviruses. Antimicrob. Agents Chemother. 2022, 66, e01581-21. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, S.; Shin, D.H.; Kim, M.-S. Inhibition of SARS-CoV 3CL protease by flavonoids. J. Enzyme Inhib. Med. Chem. 2020, 35, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Sharma, P.; Joshi, T.; Pundir, H.; Mathpal, S.; Chandra, S. Structure-based screening of novel lichen compounds against SARS Coronavirus main protease (Mpro) as potentials inhibitors of COVID-19. Mol. Divers. 2021, 25, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Hamdoun, S.; Chen, R.; Yang, L.; Ip, C.K.; Qu, Y.; Li, R.; Jiang, H.; Yang, Z.; Chung, S.K.; et al. Identification of natural compounds as SARS-CoV-2 entry inhibitors by molecular docking-based virtual screening with bio-layer interferometry. Pharmacol. Res. 2021, 172, 105820. [Google Scholar] [CrossRef]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020, 10, 766–788. [Google Scholar] [CrossRef]
- Zambare, V.P.; Christopher, L.P. Biopharmaceutical potential of lichens. Pharm. Biol. 2012, 50, 778–798. [Google Scholar] [CrossRef]
- Honegger, R. Lichens and Their Allies Past and Present. Mycota 2022, 5, 133–183. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Grube, M. Lichens redefined as complex ecosystems. New Phytol. 2020, 227, 1281–1283. [Google Scholar] [CrossRef]
- Huneck, S.; Yoshimura, I. Identification of Lichen Substances. In Identification of Lichen Substances; Huneck, S., Yoshimura, I., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 11–123. ISBN 978-3-642-85243-5. [Google Scholar]
- Parrot, D.; Antony-Babu, S.; Intertaglia, L.; Grube, M.; Tomasi, S.; Suzuki, M.T. Littoral lichens as a novel source of potentially bioactive Actinobacteria. Sci. Rep. 2015, 5, 15839. [Google Scholar] [CrossRef]
- Lagarde, A.; Jargeat, P.; Roy, M.; Girardot, M.; Imbert, C.; Millot, M.; Mambu, L. Fungal communities associated with Evernia prunastri, Ramalina fastigiata and Pleurosticta acetabulum: Three epiphytic lichens potentially active against Candida biofilms. Microbiol. Res. 2018, 211, 1–12. [Google Scholar] [CrossRef]
- Desmarets, L.; Callens, N.; Hoffmann, E.; Danneels, A.; Lavie, M.; Couturier, C.; Dubuisson, J.; Belouzard, S.; Rouillé, Y. A reporter cell line for the automated quantification of SARS-CoV-2 infection in living cells. Front. Microbiol. 2022, 13, 1031204. [Google Scholar] [CrossRef] [PubMed]
- Belouzard, S.; Machelart, A.; Sencio, V.; Vausselin, T.; Hoffmann, E.; Deboosere, N.; Rouillé, Y.; Desmarets, L.; Séron, K.; Danneels, A.; et al. Clofoctol inhibits SARS-CoV-2 replication and reduces lung pathology in mice. PLoS Pathog. 2022, 18, e1010498. [Google Scholar] [CrossRef] [PubMed]
- Dieu, A.; Mambu, L.; Champavier, Y.; Chaleix, V.; Sol, V.; Gloaguen, V.; Millot, M. Antibacterial activity of the lichens Usnea Florida and Flavoparmelia caperata (Parmeliaceae). Nat. Prod. Res. 2020, 34, 3358–3362. [Google Scholar] [CrossRef]
- Le Pogam, P.; Boustie, J.; Richomme, P.; Denis, A.; Schinkovitz, A. The inherent matrix properties of lichen metabolites in matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2017, 31, 1993–2002. [Google Scholar] [CrossRef] [PubMed]
- Parrot, D.; Intertaglia, L.; Jehan, P.; Grube, M.; Suzuki, M.T.; Tomasi, S. Chemical analysis of the Alphaproteobacterium strain MOLA1416 associated with the marine lichen Lichina pygmaea. Phytochemistry 2018, 145, 57–67. [Google Scholar] [CrossRef]
- Noël, A.; Ferron, S.; Rouaud, I.; Gouault, N.; Hurvois, J.-P.; Tomasi, S. Isolation and Structure Identification of Novel Brominated Diketopiperazines from Nocardia ignorata—A Lichen-Associated Actinobacterium. Molecules 2017, 22, 371. [Google Scholar] [CrossRef]
- Parrot, D.; Legrave, N.; Intertaglia, L.; Rouaud, I.; Legembre, P.; Grube, M.; Suzuki, M.T.; Tomasi, S. Cyaneodimycin, a Bioactive Compound Isolated from the Culture of Streptomyces cyaneofuscatus Associated with Lichina confinis. Eur. J. Org. Chem. 2016, 2016, 3977–3982. [Google Scholar] [CrossRef]
- Lagarde, A.; Mambu, L.; Mai, P.-Y.; Champavier, Y.; Stigliani, J.-L.; Beniddir, M.A.; Millot, M. Chlorinated bianthrones from the cyanolichen Nephroma laevigatum. Fitoterapia 2021, 149, 104811. [Google Scholar] [CrossRef]
- Toure, S.; Millot, M.; Ory, L.; Roullier, C.; Khaldi, Z.; Pichon, V.; Girardot, M.; Imbert, C.; Mambu, L. Access to Anti-Biofilm Compounds from Endolichenic Fungi Using a Bioguided Networking Screening. J. Fungi 2022, 8, 1012. [Google Scholar] [CrossRef]
- Parrot, D.; Peresse, T.; Hitti, E.; Carrie, D.; Grube, M.; Tomasi, S. Qualitative and spatial metabolite profiling of lichens by a LC-MS approach combined with optimised extraction. Phytochem. Anal. 2015, 26, 23–33. [Google Scholar] [CrossRef]
- Bauer, J.; Waltenberger, B.; Noha, S.M.; Schuster, D.; Rollinger, J.M.; Boustie, J.; Chollet, M.; Stuppner, H.; Werz, O. Discovery of depsides and depsidones from lichen as potent inhibitors of microsomal prostaglandin E2 synthase-1 using pharmacophore models. ChemMedChem 2012, 7, 2077–2081. [Google Scholar] [CrossRef]
- Delebassée, S.; Mambu, L.; Pinault, E.; Champavier, Y.; Liagre, B.; Millot, M. Cytochalasin E in the lichen Pleurosticta acetabulum. Anti-proliferative activity against human HT-29 colorectal cancer cells and quantitative variability. Fitoterapia 2017, 121, 146–151. [Google Scholar] [CrossRef]
- Millot, M.; Tomasi, S.; Articus, K.; Rouaud, I.; Bernard, A.; Boustie, J. Metabolites from the Lichen Ochrolechia parella growing under two different heliotropic conditions. J. Nat. Prod. 2007, 70, 316–318. [Google Scholar] [CrossRef]
- Dieu, A.; Millot, M.; Champavier, Y.; Mambu, L.; Chaleix, V.; Sol, V.; Gloaguen, V. Uncommon Chlorinated Xanthone and Other Antibacterial Compounds from the Lichen Cladonia incrassata. Planta Med. 2014, 80, 931–935. [Google Scholar] [CrossRef]
- Millot, M.; Martin-de-Lassalle, M.; Chollet-Krugler, M.; Champavier, Y.; Mambu, L.; Chulia, J.-A.; Lacaille-Dubois, M.-A. Two New Retigerane-Type Sesterterpenoids from the Lichen Leprocaulon microscopicum. Helv. Chim. Acta 2016, 99, 169–173. [Google Scholar] [CrossRef]
- Horhant, D.; Lamer, A.-C.L.; Boustie, J.; Uriac, P.; Gouault, N. Separation of a mixture of paraconic acids from Cetraria islandica (L.) Ach. employing a fluorous tag—Catch and release strategy. Tetrahedron Lett. 2007, 48, 6031–6033. [Google Scholar] [CrossRef]
- Sweidan, A.; Chollet-Krugler, M.; van de Weghe, P.; Chokr, A.; Tomasi, S.; Bonnaure-Mallet, M.; Bousarghin, L. Design, synthesis and biological evaluation of potential antibacterial butyrolactones. Bioorg. Med. Chem. 2016, 24, 5823–5833. [Google Scholar] [CrossRef]
- Huneck, S.; Preiss, A.; Schmidt, J.; Mendez, A.M. 3β-Acetoxyhopan-1β,22-diol, a triterpene from the lichen Pseudoparmelia texana. Phytochemistry 1983, 22, 2027–2030. [Google Scholar] [CrossRef]
- Lagarde, A. Études Phytochimiques du Lichen Nephroma laevigatum et de ses Champignons Endolichéniques. Évaluation des Activités Antiprolifératives et Anti-Biofilms. Ph.D. Thesis, Université de Limoges, Limoges, France, 2017. [Google Scholar]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the Mycotoxin Enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef]
- Bendz, G.; Bohman, G.; Santesson, J. Chlorinated Anthraquinones from Nephroma laevigatum. Acta Chem. Scand. 1967, 21, 2889–2890. [Google Scholar] [CrossRef]
- Dwivedi, S.P.D.; Pandey, V.B.; Shah, A.H.; Rao, Y.B. Chemical constituents of Rhamnus procumbens and pharmacological actions of emodin. Phytother. Res. 1988, 2, 51–53. [Google Scholar] [CrossRef]
- Ho, T.-Y.; Wu, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-H.; Fang, M.-C.; Chen, Y.-Z.; Huang, S.-C.; Wang, D.-Y. Quantitative analysis of fragrance allergens in various matrixes of cosmetics by liquid-liquid extraction and GC-MS. J. Food Drug Anal. 2021, 29, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, D.N.; Zarubaev, V.V.; Shtro, A.A.; Polovinka, M.P.; Luzina, O.A.; Komarova, N.I.; Salakhutdinov, N.F.; Kiselev, O.I. Anti-viral activity of (−)- and (+)-usnic acids and their derivatives against influenza virus A(H1N1)2009. Bioorg. Med. Chem. Lett. 2012, 22, 7060–7064. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Sahu, N.; Singh, A.P.; Singh, V.K.; Singh, S.C.; Upadhye, V.J.; Mathew, A.T.; Kumar, R.; Sinha, R.P. Exploration of Novel Lichen Compounds as Inhibitors of SARS-CoV-2 Mpro: Ligand-Based Design, Molecular Dynamics, and ADMET Analyses. Appl. Biochem. Biotechnol. 2022, 194, 6386–6406. [Google Scholar] [CrossRef] [PubMed]
- Prateeksha, G.; Rana, T.S.; Ashthana, A.K.; Barik, S.K.; Singh, B.N. Screening of cryptogamic secondary metabolites as putative inhibitors of SARS-CoV-2 main protease and ribosomal binding domain of spike glycoprotein by molecular docking and molecular dynamics approaches. J. Mol. Struct. 2021, 1240, 130506. [Google Scholar] [CrossRef]
- Oettl, S.K.; Gerstmeier, J.; Khan, S.Y.; Wiechmann, K.; Bauer, J.; Atanasov, A.G.; Malainer, C.; Awad, E.M.; Uhrin, P.; Heiss, E.H.; et al. Imbricaric acid and perlatolic acid: Multi-targeting anti-inflammatory depsides from Cetrelia monachorum. PLoS ONE 2013, 8, e76929. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Name of Compound | Class of Compound | Source | Reference |
|---|---|---|---|---|
| Aliphatic acids | ||||
| 1 | Caperatic acid | Flavoparmelia caperata | [34] | |
| 2 | Roccellic acid | Lepraria membranacea | [35] | |
| Amino-acid derivatives | ||||
| 3 | N-Acetyltyramine | Endolichenic fungus Nemania aena var aureolatum | * | |
| 4 | Cyclo(L-Tyr-L-Tpn) | α-Proteobacterium MOLA 1416 (lichen-associated bacterium) | [36] | |
| 5 | Cyclo(L-Br-Tyr-D-Pro) | Nocardia ignorata (lichen-associated bacterium) | [37] | |
| 6 | Enniatin B | Fusarium avenaceum (endolichenic fungus) | * | |
| 7 | Indole-3-carboxaldehyde | Nocardia ignorata (lichen-associated bacterium) | [37] | |
| 8 | N-Methyldactinomycin | Streptomyces cyaneofuscatus (lichen-associated bacterium) | [38] | |
| Anthraquinones | ||||
| 9 | Emodin | Nephroma laevigatum | [39] | |
| 10 | Parietin | Xanthoria parietina | [29] | |
| 11 | (+)-Rugulosin | Coniochaeta lignicola (endolichenic fungi) | [40] | |
| 12 | Solorinic acid | Solorina crocea | [29] | |
| Chromones and Xanthones | ||||
| 13 | 2,5-dichlorolichexanthon | Pertusaria aleianta | [29] | |
| 14 | Lepraric acid | Roccella fuciformis | [41] | |
| 15 | Tri-O-methylarthothelin | Dimelaena cf. australiensis | [29] | |
| Depsides | ||||
| 16 | Atranorin | Parmotrema tinctorum | [29] | |
| 17 | Erythrin | Roccella phycopsis | [36] | |
| 18 | Evernic acid | Evernia prunastri | [29] | |
| 19 | Perlatolic acid | Cladonia portentosa | [42] | |
| 20 | Squamatic acid | Cladonia squamosa | * | |
| Depsidones | ||||
| 21 | Fumarprotocetraric acid | Cetraria islandica | [29] | |
| 22 | Norstictic acid | Pleurosticta acetabulum | [43] | |
| 23 | Psoromic acid | Squamarina cartilaginea | [29] | |
| 24 | Variolaric acid | Ochrolechia parella | [44] | |
| 25 | Salazinic acid | Parmelia saxatilis | * | |
| Dibenzofurans | ||||
| 26 | Didymic acid | Cladonia incrassata | [45] | |
| 27 | (−)-Placodiolic acid | Leprocaulum microscopicum | [46] | |
| 28 | (−)-Usnic acid | Leprocaulum microscopicum | [46] | |
| Lactones and macrolides | ||||
| 29 | Aspicilin | Aspicilia caesiocinerea | [29] | |
| 30 | (+)-Roccellaric acid | Cetraria islandica | [47] | |
| 31 | (−)-Lichesterinic acid | Cetraria komarovii Synthetic compound | [48] | |
| Pulvinic acid derivative | ||||
| 32 | Vulpinic acid | Letharia vulpina | [29] | |
| Phenolic acid and derivatives | ||||
| 33 | β-Orcinol carboxylic acid | Pseudevernia furfuracea | [29] | |
| 34 | Chloroatranol | Evernia prunastri | [29] | |
| 35 | Tyrosol acetate | Endolichenic fungi Nemania aena var aureolatum | * | |
| 36 | Tyrosol | Endolichenic fungi Nemania aena var aureolatum | * | |
| Terpenes | ||||
| 37 | 3β-Acetoxyhopan-1β,22-diol | Pseudoparmelia texana | [49] | |
| 38 | Helvolic acid | Endolichenic fungi Nemania aena var aureolatum | [50] | |
| 39 | 16α-Hydroxykauran | Ramalina tumidula | [29] | |
| 40 | Hopane-6α,7β,22-triol | Nephroma laevigatum | * | |
| 41 | Zeorin | Leprocaulum microscopicum | [46] | |
| Others derivatives | ||||
| 42 | 6-Methoxy-2-methyl-3-heptylprodiginine | α-Proteobacterium MOLA 1416 (lichen-associated bacterium) | [36] |
| Huh-7 | Huh-7/TMPRSS2 | ||||
|---|---|---|---|---|---|
| CC50 (μM) * | IC50 (μM) | SI | IC50 (μM) | SI | |
| Perlatolic acid | 48.9 | 20.81 ± 7.44 | 2.3 | 16.42 ± 1.66 | 3.0 |
| Vulpinic acid | 155 | 19.86 ± 9.46 | 7.8 | 14.58 ± 5.55 | 10.6 |
| Emodin | >185 | 59.58 ± 1.62 | >3.1 | 59.25 ± 2.47 | >3.1 |
| Chloroatranol | >268 | 65.81 ± 6.27 | >4 | 68.86 ± 11.52 | >3.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desmarets, L.; Millot, M.; Chollet-Krugler, M.; Boustie, J.; Camuzet, C.; François, N.; Rouillé, Y.; Belouzard, S.; Tomasi, S.; Mambu, L.; et al. Lichen or Associated Micro-Organism Compounds Are Active against Human Coronaviruses. Viruses 2023, 15, 1859. https://doi.org/10.3390/v15091859
Desmarets L, Millot M, Chollet-Krugler M, Boustie J, Camuzet C, François N, Rouillé Y, Belouzard S, Tomasi S, Mambu L, et al. Lichen or Associated Micro-Organism Compounds Are Active against Human Coronaviruses. Viruses. 2023; 15(9):1859. https://doi.org/10.3390/v15091859
Chicago/Turabian StyleDesmarets, Lowiese, Marion Millot, Marylène Chollet-Krugler, Joël Boustie, Charline Camuzet, Nathan François, Yves Rouillé, Sandrine Belouzard, Sophie Tomasi, Lengo Mambu, and et al. 2023. "Lichen or Associated Micro-Organism Compounds Are Active against Human Coronaviruses" Viruses 15, no. 9: 1859. https://doi.org/10.3390/v15091859