
Establishment and Application of a Triplex Real-Time RT-PCR Assay for Differentiation of PEDV, PoRV, and PDCoV


Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses, Primers, and Probes
2.2. Clinical Samples and Nucleic Acid Extraction
2.3. Development and Optimization of the Triplex Real-Time RT-PCR
2.4. Standard Curve Generation of the Triplex Real-Time RT-PCR
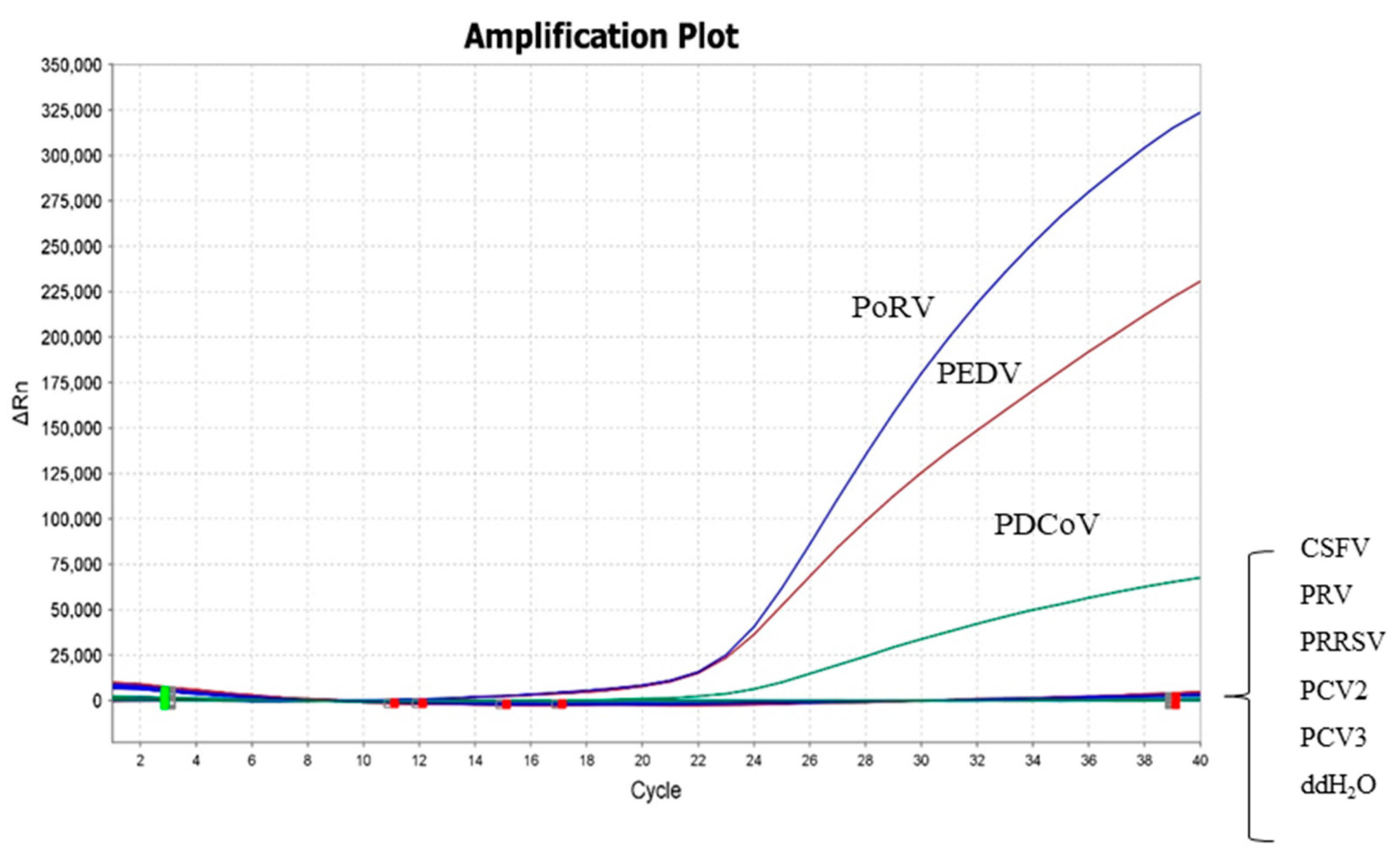
2.5. Specificity and Sensitivity of the Triplex Real-Time RT-PCR
2.6. Repeatability and Reproducibility of the Triplex Real-Time RT-PCR
2.7. Comparison of the Triplex Real-Time RT-PCR with the Commercial Single-Plex RT-PCR Kit
2.8. Clinical Application of the Triplex Real-Time RT-PCR
2.9. Sequencing and Phylogenetic Analysis
3. Results
3.1. Primers and Probes Design and Concentration Optimization
3.2. Standard Curve of the Triplex Real-Time RT-PCR
3.3. Standard Curve of the Triplex Real-Time RT-PCR
3.4. Repeatability and Reproducibility of the Triplex Real-Time RT-PCR
3.5. Comparison of the Triplex Real-Time RT-PCR with the Commercial Single-Plex Commercial RT-PCR Kit
3.6. Clinical Application of the Triplex Real-Time RT-PCR
3.7. Gene Sequencing and Phylogenetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jia, S.; Feng, B.; Wang, Z.; Ma, Y.; Gao, X.; Jiang, Y.; Cui, W.; Qiao, X.; Tang, L.; Li, Y.; et al. Dual priming oligonucleotide (DPO)-based real-time RT-PCR assay for accurate differentiation of four major viruses causing porcine viral diarrhea. Mol. Cell. Probes 2019, 47, 101435. [Google Scholar] [CrossRef] [PubMed]
- Theuns, S.; Desmarets, L.M.; Heylen, E.; Zeller, M.; Dedeurwaerder, A.; Roukaerts, I.D.; Van Ranst, M.; Matthijnssens, J.; Nauwynck, H.J. Porcine group A rotaviruses with heterogeneous VP7 and VP4 genotype combinations can be found together with enteric bacteria on Belgian swine farms. Vet. Microbiol. 2014, 172, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Hu, H.; Saif, L.J. Porcine deltacoronavirus infection: Etiology, cell culture for virus isolation and propagation, molecular epidemiology and pathogenesis. Virus Res. 2016, 226, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.Y.; Wang, Q.H. Prevention and Control of Porcine Epidemic Diarrhea: The Development of Recombination-Resistant Live Attenuated Vaccines. Viruses 2022, 14, 1317. [Google Scholar] [CrossRef]
- Wood, E.N. An apparently new syndrome of porcine epidemic diarrhoea. Vet. Rec. 1977, 100, 243–244. [Google Scholar] [CrossRef]
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef]
- Wang, D.; Fang, L.R.; Xiao, S.B. Porcine epidemic diarrhea in China. Virus Res. 2016, 226, 7–13. [Google Scholar] [CrossRef]
- Stevenson, G.W.; Hoang, H.; Schwartz, K.J.; Burrough, E.R.; Sun, D.; Madson, D.; Cooper, V.L.; Pillatzki, A.; Gauger, P.; Schmitt, B.J.; et al. Emergence of Porcine epidemic diarrhea virus in the United States: Clinical signs, lesions, and viral genomic sequences. J. Vet. Diagn. Investig. 2013, 25, 649–654. [Google Scholar] [CrossRef]
- Kumar, D.; Shepherd, F.K.; Springer, N.L.; Mwangi, W.; Marthaler, D.G. Rotavirus Infection in Swine: Genotypic Diversity, Immune Responses, and Role of Gut Microbiome in Rotavirus Immunity. Pathogens 2022, 11, 1078. [Google Scholar] [CrossRef]
- Park, G.N.; Kim, D.I.; Choe, S.; Shin, J.; An, B.H.; Kim, K.S.; Hyun, B.H.; Lee, J.S.; An, D.J. Genetic Diversity of Porcine Group A Rotavirus Strains from Pigs in South Korea. Viruses 2022, 14, 2522. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Amimo, J.O.; Saif, L.J. Porcine Rotaviruses: Epidemiology, Immune Responses and Control Strategies. Viruses 2017, 9, 48. [Google Scholar] [CrossRef]
- Theil, K.W.; Saif, L.J.; Moorhead, P.D.; Whitmoyer, R.E. Porcine rotavirus-like virus (group B rotavirus): Characterization and pathogenicity for gnotobiotic pigs. J. Clin. Microbiol. 1985, 21, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Turlewicz-Podbielska, H.; Pomorska-Mol, M. Porcine Coronaviruses: Overview of the State of the Art. Virol. Sin. 2021, 36, 833–851. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.Y.; Lau, S.K.P.; Lam, C.S.F.; Lau, C.C.Y.; Tsang, A.K.L.; Lau, J.H.N.; Bai, R.; Teng, J.L.L.; Tsang, C.C.C.; Wang, M.; et al. Discovery of Seven Novel Mammalian and Avian Coronaviruses in the Genus Deltacoronavirus Supports Bat Coronaviruses as the Gene Source of Alphacoronavirus and Betacoronavirus and Avian Coronaviruses as the Gene Source of Gammacoronavirus and Deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [PubMed]
- Marthaler, D.; Jiang, Y.; Collins, J.; Rossow, K. Complete Genome Sequence of Strain SDCV/USA/Illinois121/2014, a Porcine Deltacoronavirus from the United States. Genome Announc. 2014, 2, e00218-14. [Google Scholar] [CrossRef]
- Li, G.; Chen, Q.; Harmon, K.M.; Yoon, K.-J.; Schwartz, K.J.; Hoogland, M.J.; Gauger, P.C.; Main, R.G.; Zhang, J. Full-Length Genome Sequence of Porcine Deltacoronavirus Strain USA/IA/2014/8734. Genome Announc. 2014, 2, e00278-14. [Google Scholar] [CrossRef]
- Wang, L.Y.; Byrum, B.; Zhang, Y. Porcine Coronavirus HKU15 Detected in 9 US States, 2014. Emerg. Infect. Dis. 2014, 20, 1594–1595. [Google Scholar] [CrossRef]
- Zhang, J.Q. Porcine deltacoronavirus: Overview of infection dynamics, diagnostic methods, prevalence and genetic evolution. Virus Res. 2016, 226, 71–84. [Google Scholar] [CrossRef]
- Shu, X.; Han, F.; Hu, Y.; Hao, C.; Li, Z.; Wei, Z.; Zhang, H. Co-infection of porcine deltacoronavirus and porcine epidemic diarrhoea virus alters gut microbiota diversity and composition in the colon of piglets. Virus Res. 2022, 322, 198954. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, L.; Yang, C.; Zheng, Y.; Gauger, P.C.; Anderson, T.; Harmon, K.M.; Zhang, J.; Yoon, K.J.; Main, R.G.; et al. The emergence of novel sparrow deltacoronaviruses in the United States more closely related to porcine deltacoronaviruses than sparrow deltacoronavirus HKU17. Emerg. Microbes Infect. 2018, 7, 105. [Google Scholar] [CrossRef]
- Chen, N.; Xiao, Y.; Li, X.; Li, S.; Xie, N.; Yan, X.; Li, X.; Zhu, J. Development and application of a quadruplex real-time PCR assay for differential detection of porcine circoviruses (PCV1 to PCV4) in Jiangsu province of China from 2016 to 2020. Transbound. Emerg. Dis. 2021, 68, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Li, X.D.; Hu, Y.X.; Liu, P.G.; Zhu, Z.B.; Liu, P.R.; Chen, C.H.; Wu, X.D. Development and application of a duplex real-time PCR assay for differentiation of genotypes I and II African swine fever viruses. Transbound. Emerg. Dis. 2022, 69, 2971–2979. [Google Scholar] [CrossRef] [PubMed]
- Wittwer, C.T.; Vandesompele, J.; Shipley, G.L.; Pfaffl, M.W.; Nolan, T.; Mueller, R.; Kubista, M.; Huggett, J.; Hellemans, J.; Garson, J.A.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar]
- Zhang, Y.; Chen, Y.; Zhou, J.; Wang, X.; Ma, L.; Li, J.; Yang, L.; Yuan, H.; Pang, D.; Ouyang, H. Porcine Epidemic Diarrhea Virus: An Updated Overview of Virus Epidemiology, Virulence Variation Patterns and Virus-Host Interactions. Viruses 2022, 14, 2434. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, X.; Shi, D.; Shi, H.; Zhang, X.; Feng, L. Complete genome sequence of a porcine epidemic diarrhea virus variant. J. Virol. 2012, 86, 3408. [Google Scholar] [CrossRef]
- Ben Salem, A.N.; Sergei, A.C.; Olga, P.B.; Olga, G.A.; Mahjoub, A.; Larissa, B.P. Multiplex nested RT-PCR for the detection of porcine enteric viruses. J. Virol. Methods 2010, 165, 283–293. [Google Scholar] [CrossRef]
- Liu, G.P.; Jiang, Y.H.; Opriessnig, T.; Gu, K.D.; Zhang, H.L.; Yang, Z.Q. Detection and differentiation of five diarrhea related pig viruses utilizing a multiplex PCR assay. J. Virol. Methods 2019, 263, 32–37. [Google Scholar] [CrossRef]
- Ding, G.; Fu, Y.; Li, B.; Chen, J.; Wang, J.; Yin, B.; Sha, W.; Liu, G. Development of a multiplex RT-PCR for the detection of major diarrhoeal viruses in pig herds in China. Transbound. Emerg. Dis. 2020, 67, 678–685. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, Q.; Li, B.; Cui, X.; Wei, X.; Ding, Q.; Wang, Y.; Hu, H. Prevalence, phylogenetic and evolutionary analysis of porcine deltacoronavirus in Henan province, China. Prev. Vet. Med. 2019, 166, 8–15. [Google Scholar] [CrossRef]
- Li, W.; Lei, M.K.; Li, Z.F.; Li, H.M.; Liu, Z.; He, Q.G.; Luo, R. Development of a Genetically Engineered Bivalent Vaccine against Porcine Epidemic Diarrhea Virus and Porcine Rotavirus. Viruses 2022, 14, 1746. [Google Scholar] [CrossRef]
- Papp, H.; Laszlo, B.; Jakab, F.; Ganesh, B.; De Grazia, S.; Matthijnssens, J.; Ciarlet, M.; Martella, V.; Banyai, K. Review of group A rotavirus strains reported in swine and cattle. Vet. Microbiol. 2013, 165, 190–199. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses | Primers/Probes | Sequences (5′–3′) | Reference |
|---|---|---|---|
| PEDV | PEDV-M-F | CACTCCTTAGTGGTACATTGCTTGTAGA | This study |
| PEDV-M-R | CCTTGGCGACTGTGACGAA | This study | |
| PEDV-M-P | FAM-ACAGGTAAGTCAATTACC-MGB | This study | |
| PoRV | PoRV-NSP5-F | GAAGTCTCCAGAGGATATTGGACC | This study |
| PoRV-NSP5-R | TCTTAACTGCATTCGATCTAATCGA | This study | |
| PoRV-NSP5-P | ROX-CTGATTCTGCTTCAAACG-MGB | This study | |
| PDCoV | PDCoV-N-F | CCTACTACTGACGCGTCTTGGTT | Adapted from [20] |
| PDCoV-N-R | TGCCACGAAACTGAGGATGA | Adapted from [20] | |
| PDCoV-N-P | TAMRA-TGCTCAAAGCTCAAAAC-MGB | Adapted from [20] |
| Standard Sample | Target | Intra-Reproductivity (Ct Value) | SD | Coefficients of Variation (%) | Inter-Reproductivity (Ct Value) | SD | Coefficients of Variation (%) |
|---|---|---|---|---|---|---|---|
| 6.0 × 106 copies/μL | PEDV | 14.15 | 0.36 | 2.56 | 14.35 | 0.49 | 3.41 |
| PoRV | 15.08 | 0.05 | 0.35 | 15.13 | 0.11 | 0.74 | |
| PDCoV | 13.40 | 0.34 | 2.52 | 13.46 | 0.30 | 2.24 | |
| 6.0 × 104 copies/μL | PEDV | 20.18 | 0.52 | 2.60 | 20.24 | 0.44 | 2.19 |
| PoRV | 20.99 | 0.10 | 0.48 | 21.08 | 0.20 | 0.93 | |
| PDCoV | 20.99 | 0.59 | 2.66 | 20.73 | 0.68 | 3.32 | |
| 6.0 × 102 copies/μL | PEDV | 26.00 | 1.12 | 4.32 | 25.23 | 1.02 | 4.04 |
| PoRV | 27.83 | 1.22 | 4.39 | 27.71 | 1.04 | 3.75 | |
| PDCoV | 25.40 | 1.21 | 4.76 | 23.94 | 1.03 | 4.34 |
| Sample | Commercial Real-Time RT-PCR Detection Kit | Triplex Real-Time RT-PCR Assay | ||||
|---|---|---|---|---|---|---|
| PEDV | PoRV | PDCoV | PEDV | PoRV | PDCoV | |
| 1 | + | - | - | + | - | - |
| 2 | + | - | - | + | - | - |
| 3 | + | + | - | + | + | - |
| 4 | + | + | - | + | + | - |
| 5 | + | - | - | + | - | - |
| 6 | + | + | - | + | + | - |
| 7 | + | + | - | + | + | - |
| 8 | + | + | - | + | + | - |
| 9 | - | + | - | - | + | - |
| 10 | + | + | - | + | + | - |
| 11 | + | - | - | + | - | - |
| 12 | - | + | - | - | + | - |
| 13 | - | + | - | - | + | - |
| 14 | - | + | - | - | + | - |
| 15 | - | + | - | - | + | - |
| 16 | - | - | + | - | - | + |
| PEDV+ | PoRV+ | PDCoV+ | PEDV+PoRV+ | PDCoV+PoRV+ |
|---|---|---|---|---|
| 58/112 | 67/112 | 3/112 | 26/112 | 2/112 |
| No. | Strain Name | G[P] Type |
|---|---|---|
| 1 | PoRV-01 | G9 [P23] |
| 2 | PoRV-02 | G3 [P23] |
| 3 | PoRV-03 | G4 [P13] |
| 4 | PoRV-04 | G4 [P23] |
| 5 | PoRV-05 | G11 [P13] |
| 6 | PoRV-06 | G9 [P13] |
| 7 | PoRV-07 | G9 [P23] |
| 8 | PoRV-08 | G4 [P13] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, W.; Fan, M.; Zhu, Z.; Li, X. Establishment and Application of a Triplex Real-Time RT-PCR Assay for Differentiation of PEDV, PoRV, and PDCoV. Viruses 2023, 15, 1238. https://doi.org/10.3390/v15061238
Hou W, Fan M, Zhu Z, Li X. Establishment and Application of a Triplex Real-Time RT-PCR Assay for Differentiation of PEDV, PoRV, and PDCoV. Viruses. 2023; 15(6):1238. https://doi.org/10.3390/v15061238
Chicago/Turabian StyleHou, Wenwen, Maodi Fan, Zhenbang Zhu, and Xiangdong Li. 2023. "Establishment and Application of a Triplex Real-Time RT-PCR Assay for Differentiation of PEDV, PoRV, and PDCoV" Viruses 15, no. 6: 1238. https://doi.org/10.3390/v15061238