
MAVS-Based Reporter Systems for Real-Time Imaging of EV71 Infection and Antiviral Testing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies and Reagents
2.3. MAVS-Based Reporter Cell Lines Generation
2.4. Microscopy and Image Taking
2.5. Western Blot Analysis
2.6. Viral RNA Quantification by RT-qPCR
2.7. Statistical Analysis
3. Results
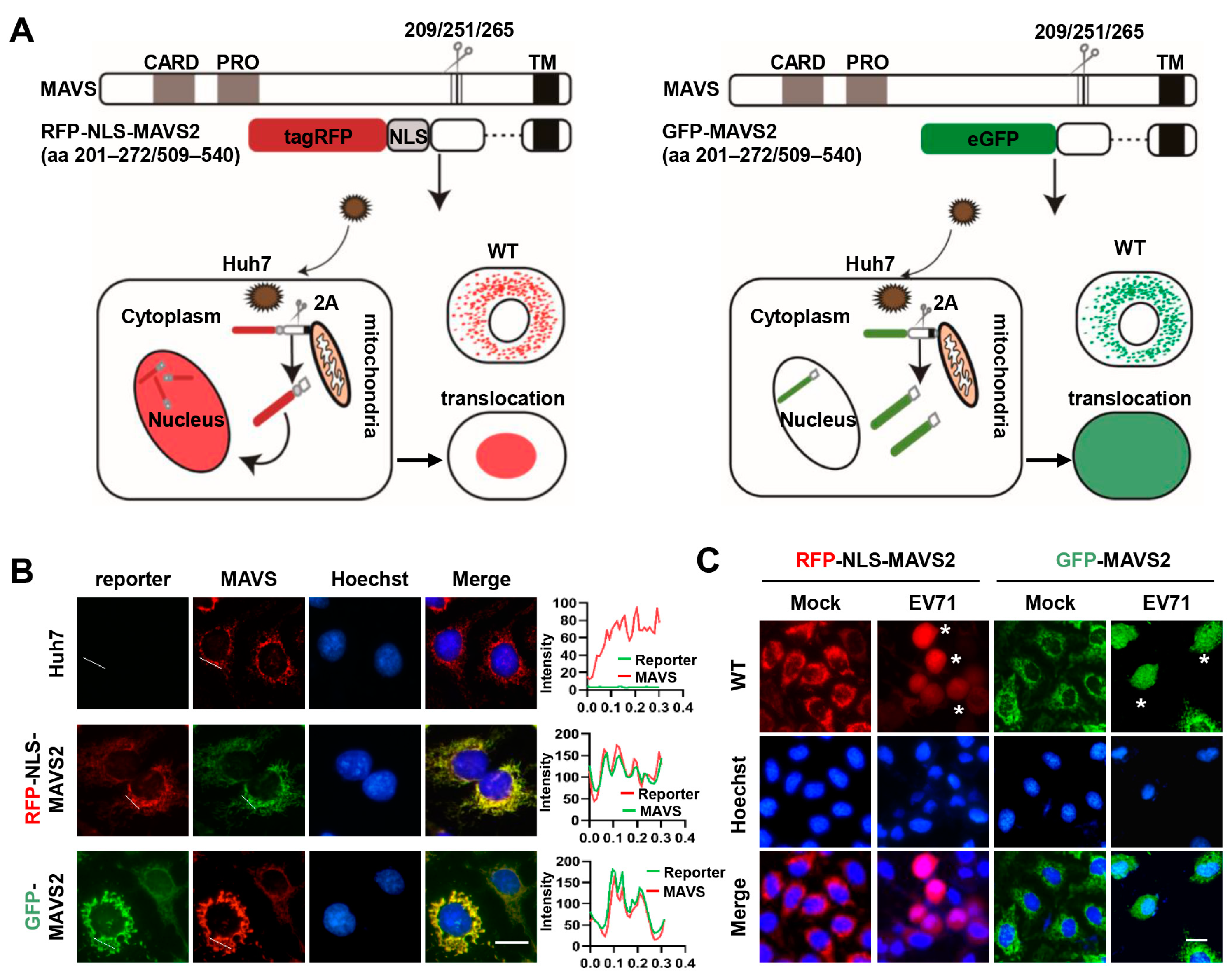
3.1. MAVS-Based Reporter Systems for Detection of EV 71 Infection
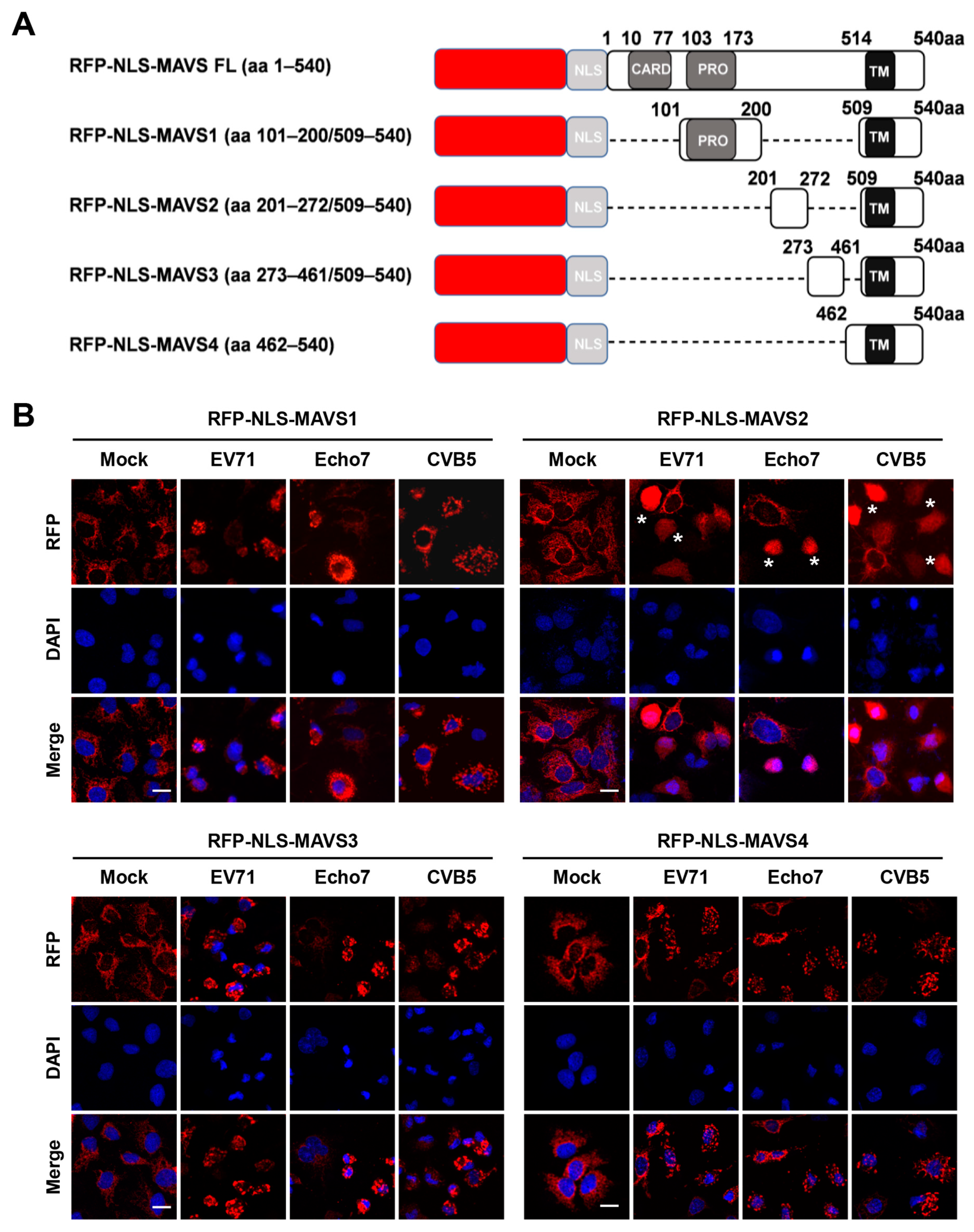
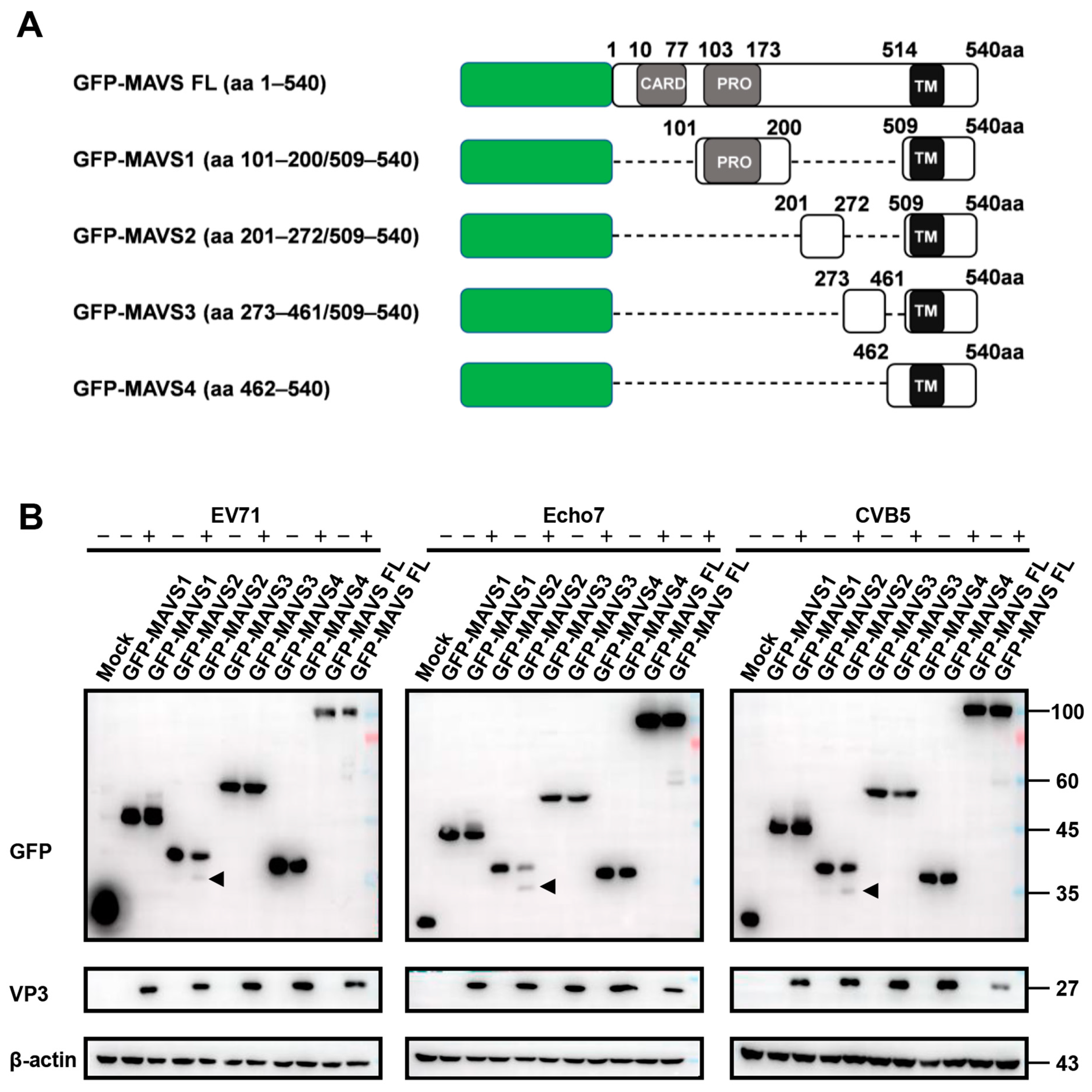
3.2. RFP-NLS-MAVS Reporter Could Be Cleaved by Different Enteroviruses
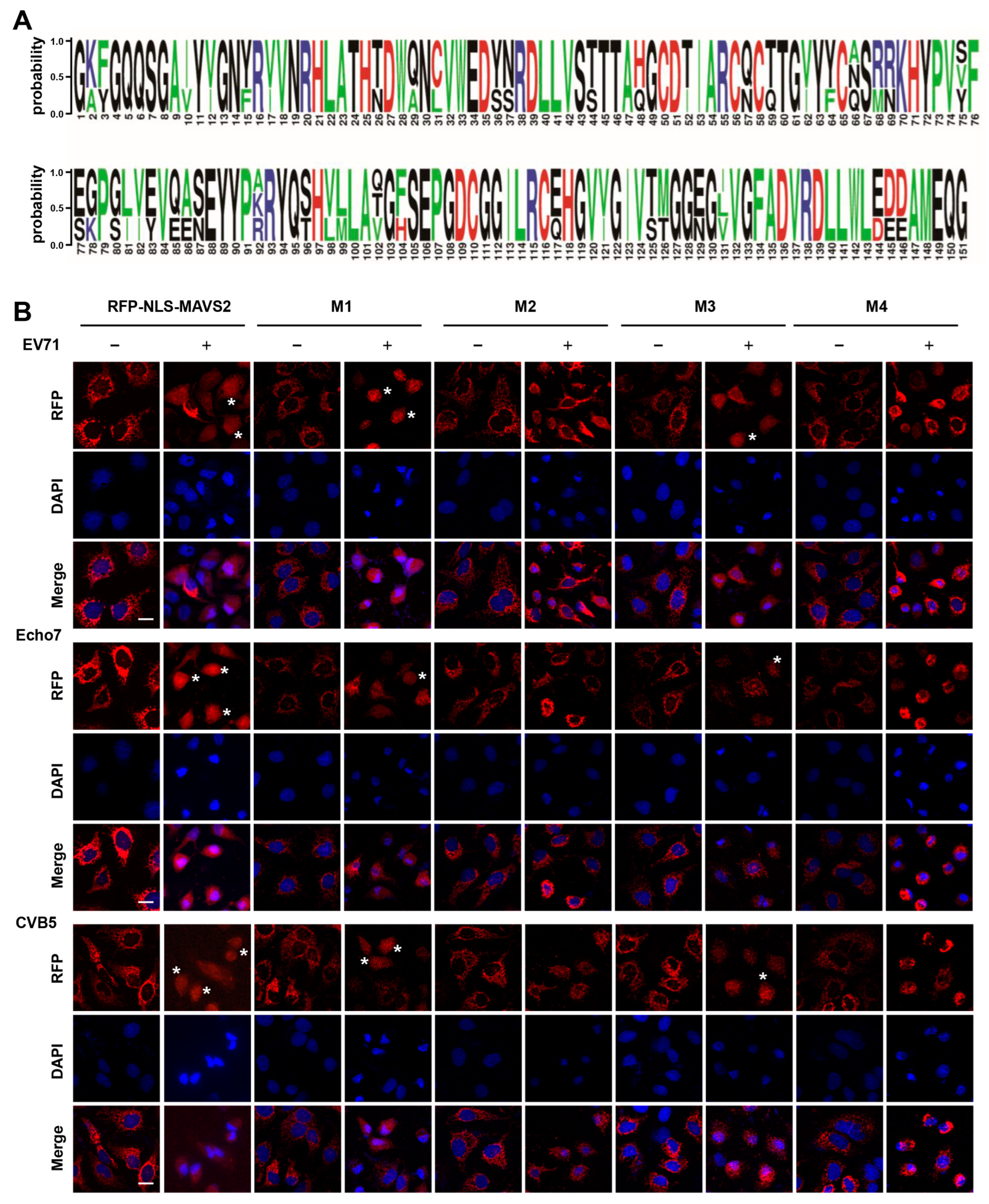
3.3. EV71, Echo7, and CVB5 Cleave the RFP-NLS-MAVS Reporter at the Same Residues
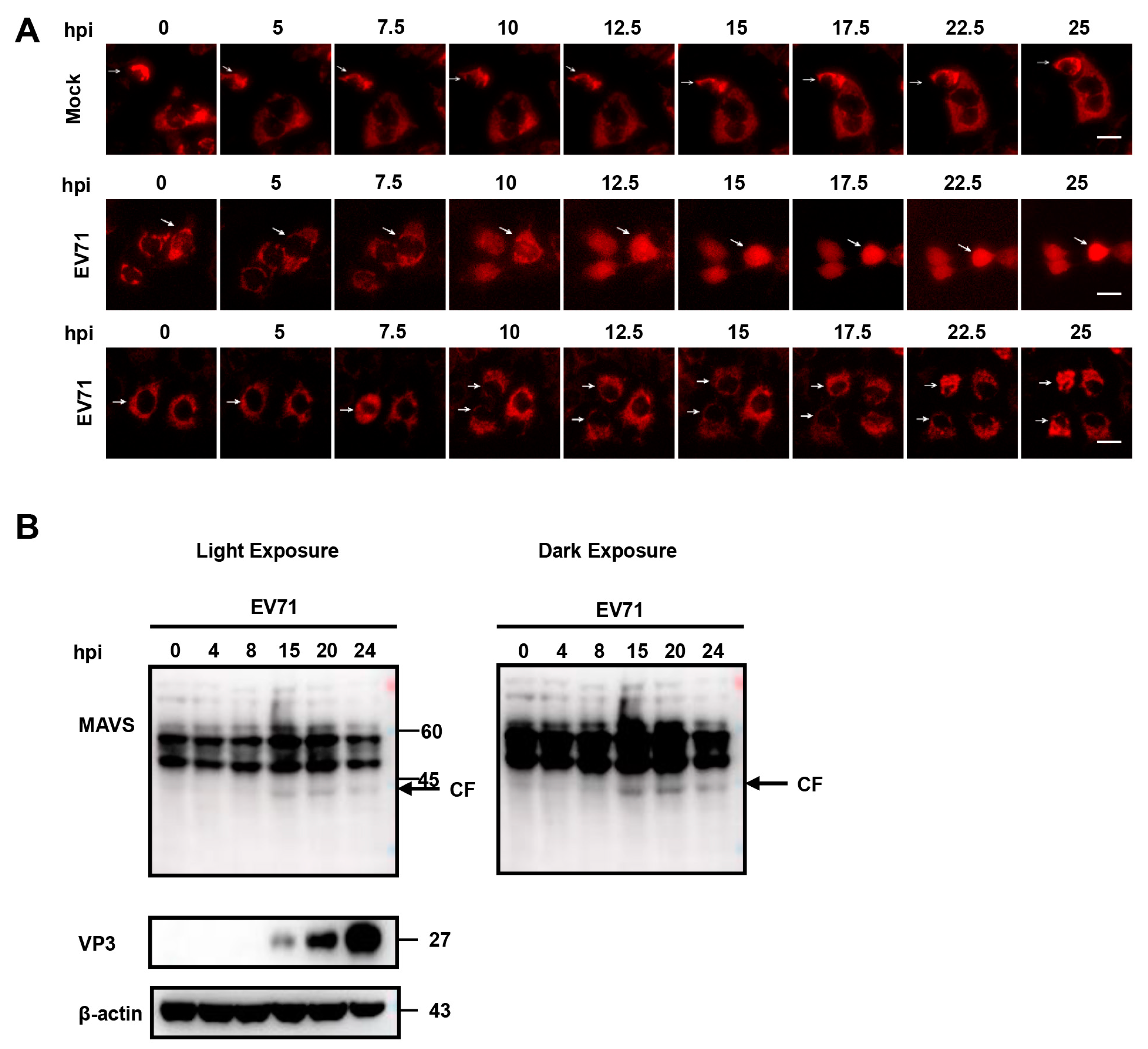
3.4. Time-Lapse Live-Cell Imaging of EV 71 Infection
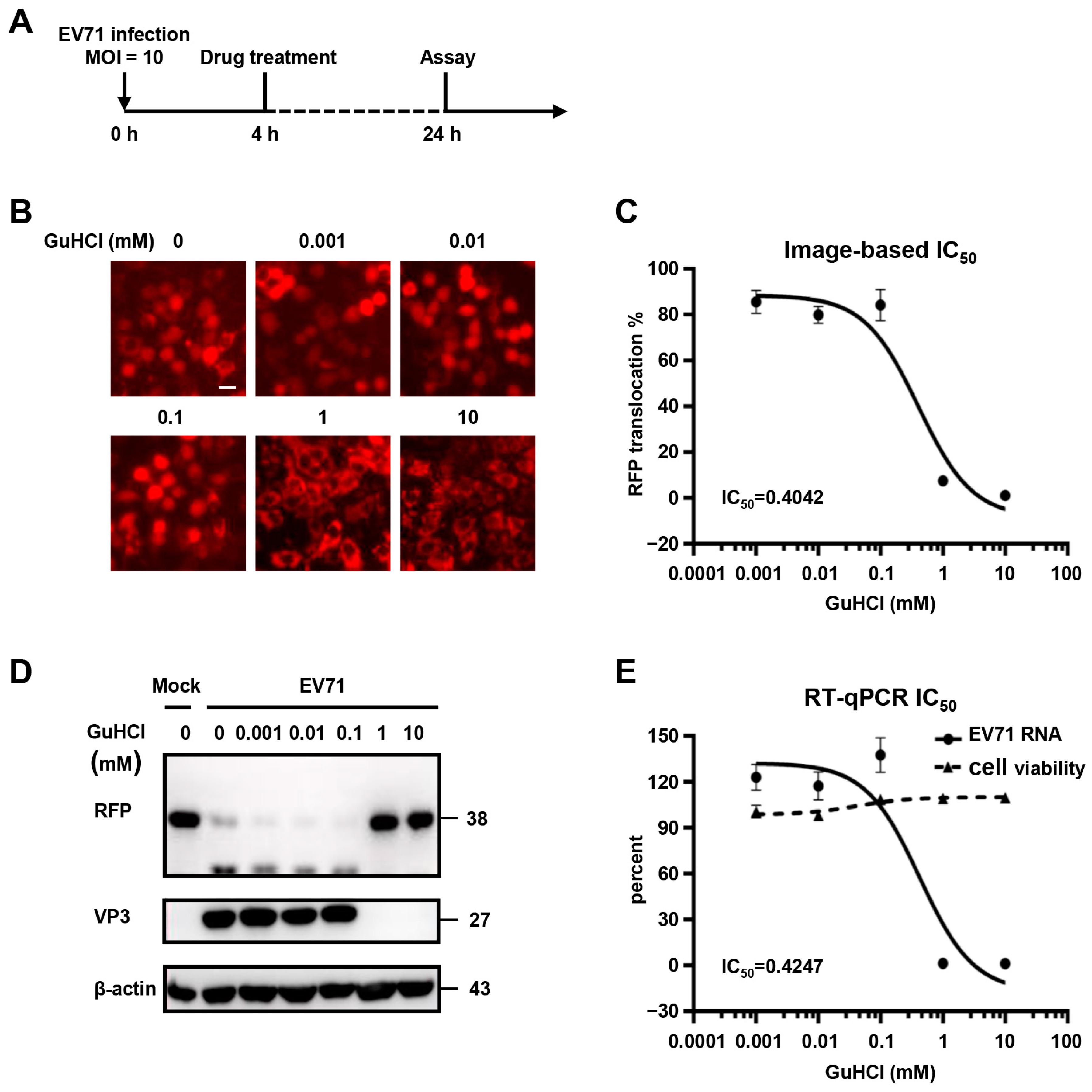
3.5. Validation of Fluorescence-Based Reporter System for Antiviral Drug Screening
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Lin, Q.; Zhang, Q.; Hu, J.; Tursun, G.; Deng, Y.; Guo, C.; Wang, X. Molecular Analysis of Caprine Enterovirus Circulating in China during 2016–2021: Evolutionary Significance. Viruses 2022, 14, 1051. [Google Scholar] [CrossRef]
- Glaser, C.; Wilson, M.R. Enteroviruses: The elephants in the room. Lancet Infect. Dis. 2020, 20, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Suresh, S.; Forgie, S.; Robinson, J. Non-polio Enterovirus detection with acute flaccid paralysis: A systematic review. J. Med. Virol. 2018, 90, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Hober, D.; Sauter, P. Pathogenesis of type 1 diabetes mellitus: Interplay between enterovirus and host. Nat. Rev. Endocrinol. 2010, 6, 279–289. [Google Scholar] [CrossRef]
- Wang, C.; Yang, R.; Yang, F.; Han, Y.; Ren, Y.; Xiong, X.; Wang, X.; Bi, Y.; Li, L.; Qiu, Y.; et al. Echovirus 11 infection induces pyroptotic cell death by facilitating NLRP3 inflammasome activation. PLoS Pathog. 2022, 18, e1010787. [Google Scholar] [CrossRef]
- Puenpa, J.; Wanlapakorn, N.; Vongpunsawad, S.; Poovorawan, Y. The History of Enterovirus A71 Outbreaks and Molecular Epidemiology in the Asia-Pacific Region. J. Biomed. Sci. 2019, 26, 75. [Google Scholar] [CrossRef]
- Kinobe, R.; Wiyatno, A.; Artika, I.M.; Safari, D. Insight into the Enterovirus A71: A review. Rev. Med. Virol. 2022, 32, e2361. [Google Scholar] [CrossRef]
- Rueca, M.; Lanini, S.; Giombini, E.; Messina, F.; Castilletti, C.; Ippolito, G.; Capobianchi, M.R.; Valli, M.B. Detection of recombinant breakpoint in the genome of human enterovirus E11 strain associated with a fatal nosocomial outbreak. Virol. J. 2022, 19, 97. [Google Scholar] [CrossRef]
- Wang, S.H.; Wang, K.; Zhao, K.; Hua, S.C.; Du, J. The Structure, Function, and Mechanisms of Action of Enterovirus Non-structural Protein 2C. Front. Microbiol. 2020, 11, 615965. [Google Scholar] [CrossRef]
- Baggen, J.; Thibaut, H.J.; Strating, J.; van Kuppeveld, F.J.M. The life cycle of non-polio enteroviruses and how to target it. Nat. Rev. Microbiol. 2018, 16, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the regulation of innate and adaptive immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Kanneganti, T.D. Innate immunity: The first line of defense against SARS-CoV-2. Nat. Immunol. 2022, 23, 165–176. [Google Scholar] [CrossRef]
- Rehwinkel, J.; Gack, M.U. RIG-I-like receptors: Their regulation and roles in RNA sensing. Nat. Rev. Immunol. 2020, 20, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.R.; Shrestha, P.; Yang, B.; Chen, Y.; Liu, S.; Maarouf, M.; Chen, J.L. Acute Infection of Viral Pathogens and Their Innate Immune Escape. Front. Microbiol. 2021, 12, 672026. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF 3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef]
- Li, X.D.; Sun, L.; Seth, R.B.; Pineda, G.; Chen, Z.J. Hepatitis C virus protease NS3/4A cleaves mitochondrial antiviral signaling protein off the mitochondria to evade innate immunity. Proc. Natl. Acad. Sci. USA 2005, 102, 17717–17722. [Google Scholar] [CrossRef]
- Feng, H.; Sander, A.L.; Moreira-Soto, A.; Yamane, D.; Drexler, J.F.; Lemon, S.M. Hepatovirus 3ABC proteases and evolution of mitochondrial antiviral signaling protein (MAVS). J. Hepatol. 2019, 71, 25–34. [Google Scholar] [CrossRef]
- Pang, L.L.; Yuan, X.H.; Shao, C.S.; Li, M.Z.; Wang, Y.; Wang, H.M.; Xie, G.C.; Xie, Z.P.; Yuan, Y.; Zhou, D.M.; et al. The suppression of innate immune response by human rhinovirus C. Biochem. Biophys. Res. Commun. 2017, 490, 22–28. [Google Scholar] [CrossRef]
- Mukherjee, A.; Morosky, S.A.; Delorme-Axford, E.; Dybdahl-Sissoko, N.; Oberste, M.S.; Wang, T.; Coyne, C.B. The coxsackievirus B 3C protease cleaves MAVS and TRIF to attenuate host type I interferon and apoptotic signaling. PLoS Pathog. 2011, 7, e1001311. [Google Scholar] [CrossRef]
- Feng, Q.; Langereis, M.A.; Lork, M.; Nguyen, M.; Hato, S.V.; Lanke, K.; Emdad, L.; Bhoopathi, P.; Fisher, P.B.; Lloyd, R.E.; et al. Enterovirus 2Apro targets MDA5 and MAVS in infected cells. J. Virol. 2014, 88, 3369–3378. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xi, X.; Lei, X.; Zhang, X.; Cui, S.; Wang, J.; Jin, Q.; Zhao, Z. Enterovirus 71 protease 2Apro targets MAVS to inhibit anti-viral type I interferon responses. PLoS Pathog. 2013, 9, e1003231. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhao, X.; Xun, M.; Ma, C.; Wang, H. Reverse Genetic Approaches for the Generation of Full Length and Subgenomic Replicon of EV71 Virus. Front. Microbiol. 2021, 12, 665879. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xun, M.; Fan, T.; Li, X.; Yao, H.; Li, X.; Wu, B.; Yang, H.; Ma, C.; Wang, H. Construction of Coxsackievirus B5 viruses with luciferase reporters and their applications in vitro and in vivo. Virol. Sin. 2023, 38. in press. [Google Scholar]
- Zhao, X.; Yuan, H.; Yang, H.; Liu, Y.; Xun, M.; Li, X.; Fan, T.; Wu, B.; Guo, S.; Wang, H. N-Acetyltransferase 8 Promotes Viral Replication by Increasing the Stability of Enterovirus 71 Nonstructural Proteins. J. Virol. 2022, 96, e0011922. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xiang, X.; Pu, Z.; Long, Y.; Xiao, D.; Zhang, W.; Li, Q.; Li, X.; Li, S.; Shao, Z.; et al. Epidemic pattern of hand-foot-and-mouth disease in Xi’an, China from 2008 through 2015. BMC Infect. Dis. 2019, 19, 19. [Google Scholar] [CrossRef]
- Chen, B.S.; Lee, H.C.; Lee, K.M.; Gong, Y.N.; Shih, S.R. Enterovirus and Encephalitis. Front. Microbiol. 2020, 11, 261. [Google Scholar] [CrossRef]
- Chuang, Y.Y.; Huang, Y.C. Enteroviral infection in neonates. J. Microbiol. Immunol. Infect. 2019, 52, 851–857. [Google Scholar] [CrossRef]
- Watzinger, F.; Ebner, K.; Lion, T. Detection and monitoring of virus infections by real-time PCR. Mol. Aspects Med. 2006, 27, 254–298. [Google Scholar] [CrossRef]
- Guo, D.; Yu, X.; Wang, D.; Li, Z.; Zhou, Y.; Xu, G.; Yuan, B.; Qin, Y.; Chen, M. SLC35B2 Acts in a Dual Role in the Host Sulfation Required for EV71 Infection. J. Virol. 2022, 96, e0204221. [Google Scholar] [CrossRef]
- Shang, B.; Deng, C.; Ye, H.; Xu, W.; Yuan, Z.; Shi, P.Y.; Zhang, B. Development and characterization of a stable eGFP enterovirus 71 for antiviral screening. Antiviral Res. 2013, 97, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Yao, X.; Chen, W.; Wang, F.; He, H.; Liu, L.; He, Y.; Chen, J.; Jiang, P.; Zhang, R.; et al. Dissecting complicated viral spreading of enterovirus 71 using in situ bioorthogonal fluorescent labeling. Biomaterials 2018, 181, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Pan, H.; Yao, X.; He, H.; Liu, L.; Luo, Y.; Zhou, H.; Zheng, M.; Zhang, R.; Ma, Y.; et al. Bioorthogonal Metabolic Labeling Utilizing Protein Biosynthesis for Dynamic Visualization of Nonenveloped Enterovirus 71 Infection. ACS Appl. Mater. Interfaces 2020, 12, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.T.; Catanese, M.T.; Law, L.M.; Khetani, S.R.; Syder, A.J.; Ploss, A.; Oh, T.S.; Schoggins, J.W.; MacDonald, M.R.; Bhatia, S.N.; et al. Real-time imaging of hepatitis C virus infection using a fluorescent cell-based reporter system. Nat. Biotechnol. 2010, 28, 167–171. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Bender, S.; Reuter, A.; Eberle, F.; Einhorn, E.; Binder, M.; Bartenschlager, R. Activation of Type I and III Interferon Response by Mitochondrial and Peroxisomal MAVS and Inhibition by Hepatitis C Virus. PLoS Pathog. 2015, 11, e1005264. [Google Scholar] [CrossRef]
- Zheng, X.; Guo, R.; Liu, Q.; Wakae, K.; Watanabe, N.; Fukano, K.; Que, L.; Li, Y.; Aly, H.H.; Watashi, K.; et al. Identification of natural compounds extracted from crude drugs as novel inhibitors of hepatitis C virus. Biochem. Biophys. Res. Commun. 2021, 567, 1–8. [Google Scholar] [CrossRef]
- Kim, H.Y.; Li, X.; Jones, C.T.; Rice, C.M.; Garcia, J.M.; Genovesio, A.; Hansen, M.A.; Windisch, M.P. Development of a multiplex phenotypic cell-based high throughput screening assay to identify novel hepatitis C virus antivirals. Antiviral Res. 2013, 99, 6–11. [Google Scholar] [CrossRef]
- Sokolinskaya, E.L.; Putlyaeva, L.V.; Polinovskaya, V.S.; Lukyanov, K.A. Genetically Encoded Fluorescent Sensors for SARS-CoV-2 Papain-like Protease PLpro. Int. J. Mol. Sci. 2022, 23, 7826. [Google Scholar] [CrossRef]
- Paci, G.; Caria, J.; Lemke, E.A. Cargo transport through the nuclear pore complex at a glance. J. Cell Sci. 2021, 134, jcs247874. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Yang, E.; Li, X.; Fan, T.; Guo, S.; Yang, H.; Wu, B.; Wang, H. MAVS-Based Reporter Systems for Real-Time Imaging of EV71 Infection and Antiviral Testing. Viruses 2023, 15, 1064. https://doi.org/10.3390/v15051064
Li X, Yang E, Li X, Fan T, Guo S, Yang H, Wu B, Wang H. MAVS-Based Reporter Systems for Real-Time Imaging of EV71 Infection and Antiviral Testing. Viruses. 2023; 15(5):1064. https://doi.org/10.3390/v15051064
Chicago/Turabian StyleLi, Xiaozhen, E Yang, Xinyu Li, Tingting Fan, Shangrui Guo, Hang Yang, Bo Wu, and Hongliang Wang. 2023. "MAVS-Based Reporter Systems for Real-Time Imaging of EV71 Infection and Antiviral Testing" Viruses 15, no. 5: 1064. https://doi.org/10.3390/v15051064