
A Web Tool to Estimate Baseline Anti-Spike Monoclonal Antibody Efficacy Based on Regional Genomic Surveillance

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Clinically Authorized Anti-Spike Monoclonal Antibodies
2.2. Inferring Baseline Resistance to Anti-Spike Monoclonal Antibodies
2.3. Calculating the Prevalence of a Given Mutation in the Database
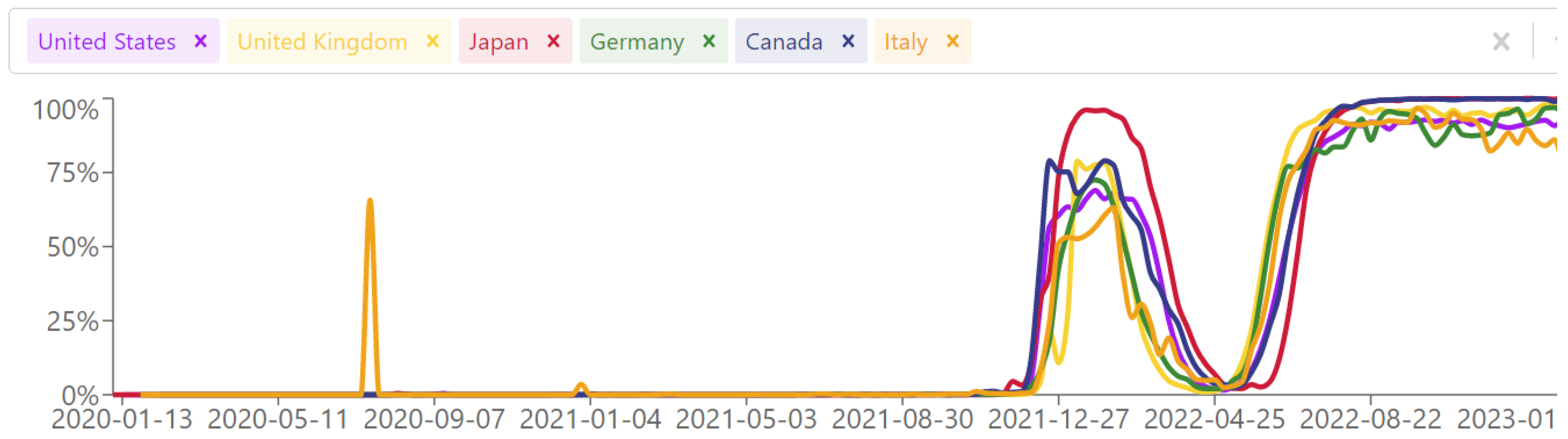
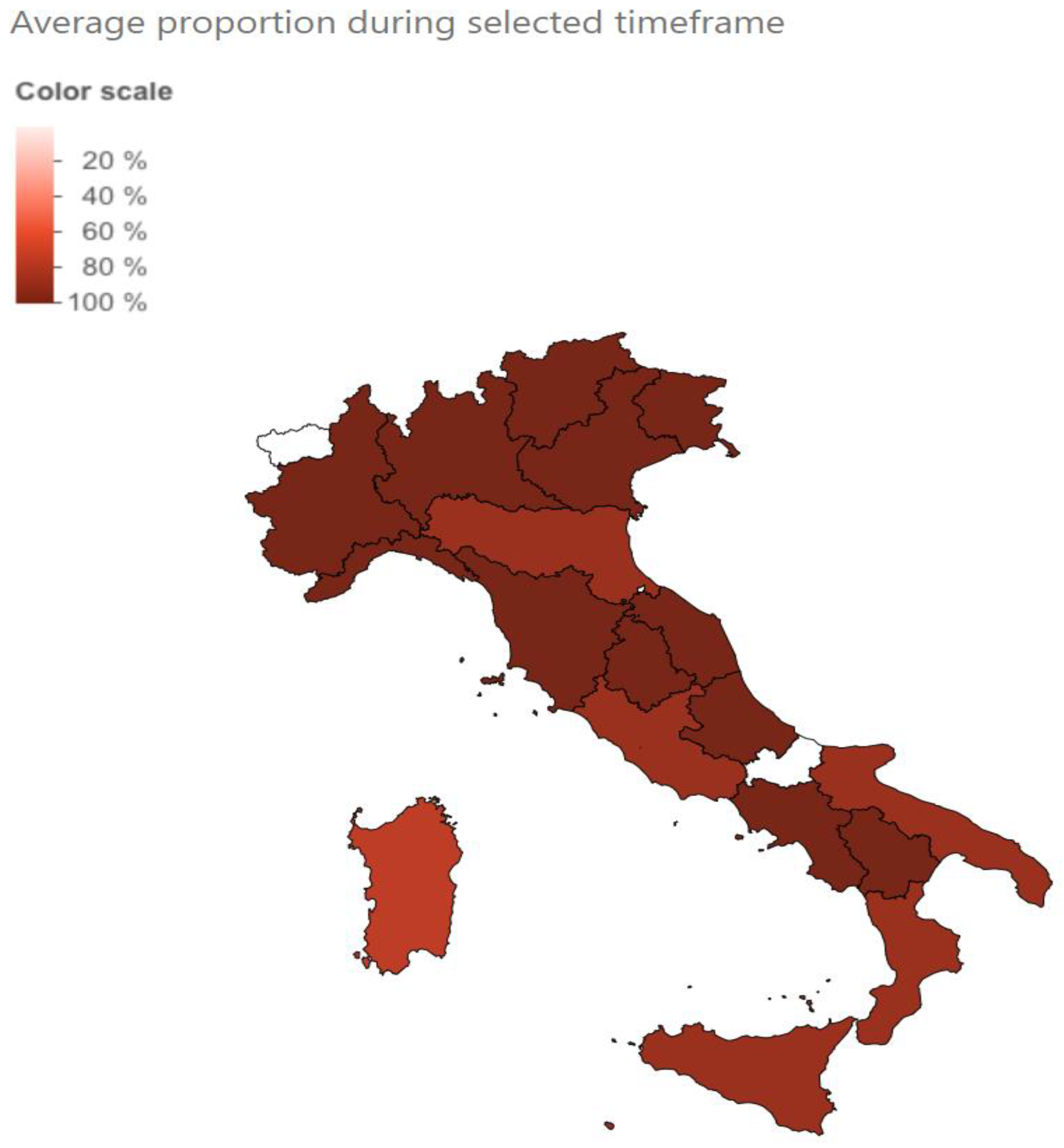
3. Results
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Focosi, D.; McConnell, S.; Casadevall, A.; Cappello, E.; Valdiserra, G.; Tuccori, M. Monoclonal antibody therapies against SARS-CoV-2. Lancet Infect. Dis. 2022, 22, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Maggi, F.; Franchini, M.; McConnell, S.; Casadevall, A. Analysis of immune escape variants from antibody-based therapeutics against COVID-19: A systematic review. Int. J. Mol. Sci. 2022, 23, 29. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Quiroga, R.; McConnell, S.; Johnson, M.C.; Casadevall, A. Convergent evolution in SARS-CoV-2 Spike creates a variant soup from which new COVID-19 waves emerge. Int. J. Mol. Sci. 2023, 24, 2264. [Google Scholar] [CrossRef] [PubMed]
- Elbe, S.; Buckland-Merrett, G. Data, disease and diplomacy: GISAID's innovative contribution to global health. Glob. Chall. 2017, 1, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Tzou, P.L.; Tao, K.; Nouhin, J.; Rhee, S.-Y.; Hu, B.D.; Pai, S.; Parkin, N.; Shafer, R.W. Coronavirus Antiviral Research Database (CoV-RDB): An Online Database Designed to Facilitate Comparisons between Candidate Anti-Coronavirus Compounds. Viruses 2020, 12, 1006. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Nadeau, S.; Yared, M.; Voinov, P.; Xie, N.; Roemer, C.; Stadler, T. CoV-Spectrum: Analysis of globally shared SARS-CoV-2 data to identify and characterize new variants. Bioinformatics 2021, 38, 1735–1737. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.S.; O’Donoghue, A.; Mechanic, O.; Dechen, T.; Stevens, J. Administration of Anti–SARS-CoV-2 Monoclonal Antibodies After US Food and Drug Administration Deauthorization. JAMA Netw. Open 2022, 5, e2228997. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Tuccori, M. Prescription of Anti-Spike Monoclonal Antibodies in COVID-19 Patients with Resistant SARS-CoV-2 Variants in Italy. Pathogens 2022, 11, 823. [Google Scholar] [CrossRef] [PubMed]
- ETF Warns that Monoclonal Antibodies May Not Be Effective against Emerging Strains of SARS-CoV-2. Available online: https://www.ema.europa.eu/en/news/etf-warns-monoclonal-antibodies-may-not-be-effective-against-emerging-strains-sars-cov-2 (accessed on 24 January 2023).
- Harman, K.; Nash, S.G.; Webster, H.H.; Groves, N.; Hardstaff, J.; Bridgen, J.; Blomquist, P.B.; Hope, R.; Ashano, E.; Myers, R.; et al. Comparison of the risk of hospitalisation among BA.1 and BA.2 COVID-19 cases treated with Sotrovimab in the community in England. arXiv 2022, arXiv:2022.2010.2021.22281171. [Google Scholar]
- Wu, M.Y.; Carr, E.J.; Harvey, R.; Mears, H.V.; Kjaer, S.; Townsley, H.; Hobbs, A.; Ragno, M.; Herman, L.S.; Adams, L.; et al. WHO's Therapeutics and COVID-19 Living Guideline on mAbs needs to be reassessed. Lancet 2022, 400, 2193–2196. [Google Scholar] [CrossRef] [PubMed]
- Stadler, E.; Chai, K.L.; Schlub, T.E.; Cromer, D.; Polizzotto, M.N.; Kent, S.J.; Beecher, C.; White, H.; Turner, T.; Skoetz, N.; et al. Determinants of passive antibody efficacy in SARS-CoV-2 infection. arXiv 2022, arXiv:2022.2003.2021.22272672. [Google Scholar]
- Stadler, E.; Burgess, M.T.; Schlub, T.E.; Chai, K.L.; McQuilten, Z.K.; Wood, E.M.; Polizzotto, M.N.; Kent, S.J.; Cromer, D.; Davenport, M.P.; et al. Monoclonal antibody levels and protection from COVID-19. arXiv 2022, arXiv:2022.2011.2022.22282199. [Google Scholar]
- Cao, Y.; Jian, F.; Wang, J.; Yu, Y.; Song, W.; Yisimayi, A.; Wang, J.; An, R.; Zhang, N.; Wang, Y.; et al. Imprinted SARS-CoV-2 humoral immunity induces convergent Omicron RBD evolution. Nature 2022, 614, 521–539. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Query | Number of Sequences as of 31 January 2023 | Submitted in the Past 10 Days |
|---|---|---|---|
| Cilgavimab resistance (ALL LINEAGES) | [1-of: S:346I, S:444E, S:444Q, S:444R, S:445A, S:446S, S:450D, S:450K, S:452R] | 1.662.667 | 85.075 |
| Tixagevimab resistance (ALL LINEAGES) | [1-of: S:371F, S:484A, S:484D, S:484K, S:486S, S:486V, S:493R, S:498R, S:990A, S:1009I] | 1.757.503 | 90.423 |
| Bebtelovimab resistance (ALL LINEAGES) | [1-of: S:444N, S:444Q, S:444T, S:445A, S:445F, S:445G, S:446D, S:446R, S:446V, S:499H, S:499R, S:449S] | 365.241 | 40.544 |
| Sotrovimab resistance (ALL LINEAGES) | [1-of: S:337H, S:337L, S:337R, S:337T, S:340A, S:340D, S:340G, S:340K, S:340Q, S:340V, S:356T, S:371F, S:371L, S:377K] | 1.689.826 | 86.695 |
| Casirivimab resistance (ALL LINEAGES) | [1-of: S:371F, S:405N, S:406D, S:406W, S:417E, S:417R, S:417N, S:417T, S:417V, S:445T, S:453F, S:455F, S:475R, S:484A, S:484K, S:484Q, S:486K, S:486L, S:486R, S:486S, S:486V, S:487R, S:493E, S:493K, S:493Q] | 1.761.550 | 91.282 |
| Imdevimab resistance (ALL LINEAGES) | [1-of: S:371F, S:371L, S:373P, S:406W, S:439K, S:440K, S:444L, S:444M, S:444N, S:444Q, S:444T, S:445A, S:446S, S:446V, S:450D, S:498H, S:498R, S:499S] | 1.758.700 | 90.426 |
| Casirivimab + imdevimab resistance (ALL LINEAGES) | [1-of: S:371F, S:405N, S:406D, S:406W, S:417E, S:417R, S:417N, S:417T, S:417V, S:445T, S:453F, S:455F, S:475R, S:484A, S:484K, S:484Q, S:486K, S:486L, S:486R, S:486S, S:486V, S:487R, S:493E, S:493K, S:493Q] and [1-of: S:371F, S:371L, S:373P, S:406W, S:439K, S:440K, S:444L, S:444M, S:444N, S:444Q, S:444T, S:445A, S:446S, S:446V, S:450D, S:498H, S:498R, S:499S] | 1.758.195 | 90.390 |
| Evusheld™ (tixagevimab + cilgavimab) resistance (ALL LINEAGES) | [1-of: S:371F, S:484A, S:484D, S:484K, S:486S, S:486V, S:493R, S:498R, S:990A, S:1009I] and [1-of: S:346I, S:444E, S:444Q, S:444R, S:445A, S:446S, S:450D, S:450K, S:452R] | 1.661.161 | 84.579 |
| bamlanivimab resistance (ALL LINEAGES) | [1-of: S:452R, S:484A, S:484D, S:484K, S:484Q, S:486R, S:486V, S:490L, S:490S, S:493H, S:493K, S:493L, S:493R, S:494P, S:494R] | 1.712.794 | 89.253 |
| etesevimab resistance (ALL LINEAGES) | [1-of: S:371F, S:371L, S:417N, S:417T, S:420N, S:456A, S:456K, S:456R, S:460K, S:460S, S:460T, S:460Y, S:475R, S:475V, S:486R, S:486L, S:486V, S:487R, S:493K, S:493L, S:493R] | 1.758.046 | 90.373 |
| bamlanivimab + etesevimab resistance | [1-of: S:371F, S:371L, S:417N, S:417T, S:420N, S:456A, S:456K, S:456R, S:460K, S:460S, S:460T, S:460Y, S:475R, S:475V, S:486R, S:486L, S:486V, S:487R, S:493K, S:493L, S:493R] and [1-of: S:452R, S:484A, S:484D, S:484K, S:484Q, S:486R, S:486V, S:490L, S:490S, S:493H, S:493K, S:493L, S:493R, S:494P, S:494R] | 1.711.633 | 88.723 |
| regdanvimab resistance | [1-of: S:452R, S:484K, S:493R] | 1.548.951 | 72.218 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Focosi, D. A Web Tool to Estimate Baseline Anti-Spike Monoclonal Antibody Efficacy Based on Regional Genomic Surveillance. Viruses 2023, 15, 1048. https://doi.org/10.3390/v15051048
Focosi D. A Web Tool to Estimate Baseline Anti-Spike Monoclonal Antibody Efficacy Based on Regional Genomic Surveillance. Viruses. 2023; 15(5):1048. https://doi.org/10.3390/v15051048
Chicago/Turabian StyleFocosi, Daniele. 2023. "A Web Tool to Estimate Baseline Anti-Spike Monoclonal Antibody Efficacy Based on Regional Genomic Surveillance" Viruses 15, no. 5: 1048. https://doi.org/10.3390/v15051048