
Seroprevalence of IgG and Subclasses against the Nucleocapsid of SARS-CoV-2 in Health Workers
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Sample Collection
2.2. Indirect ELISA for Total IgG
2.3. Standardization and Detection of IgG Subclasses
2.4. Statistical Analysis
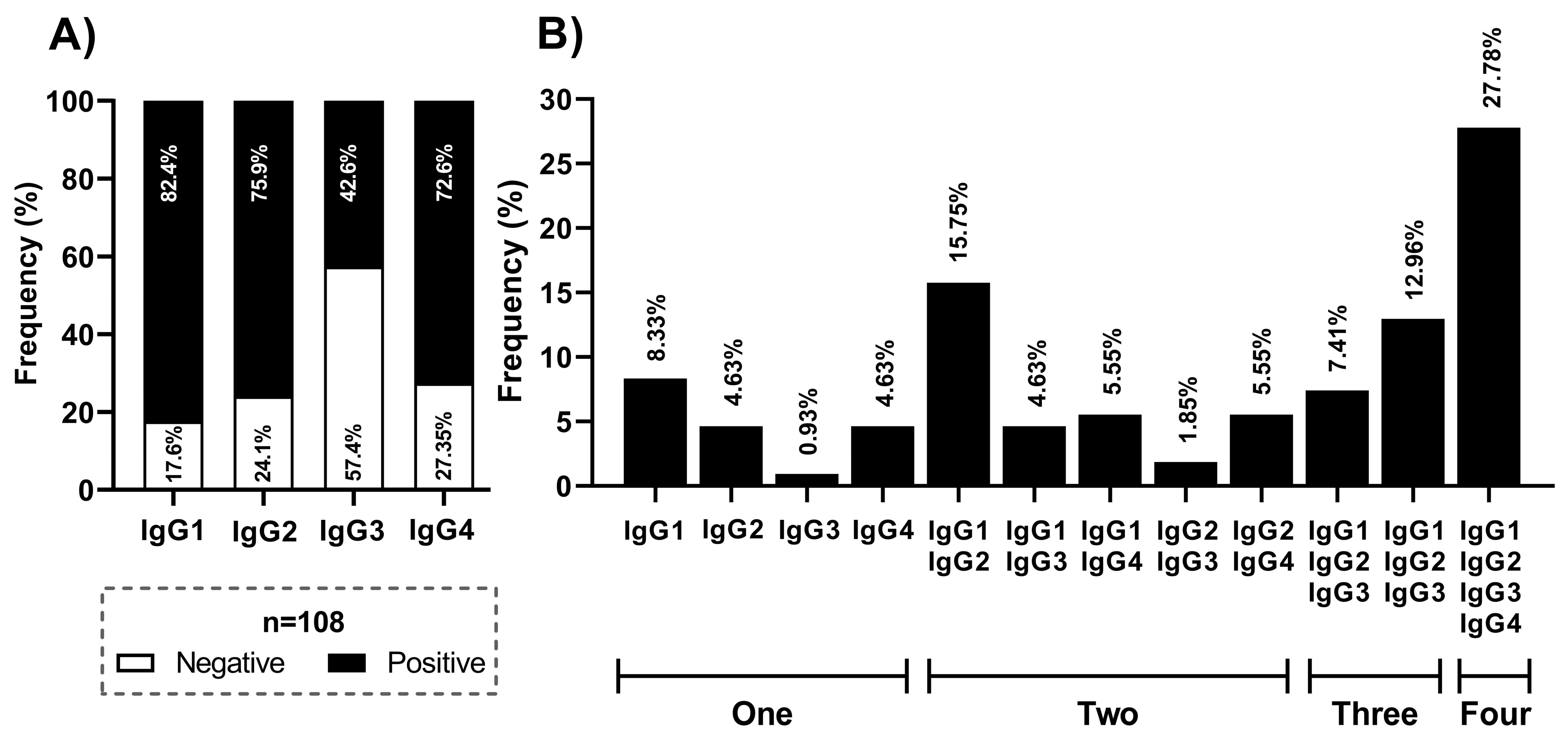
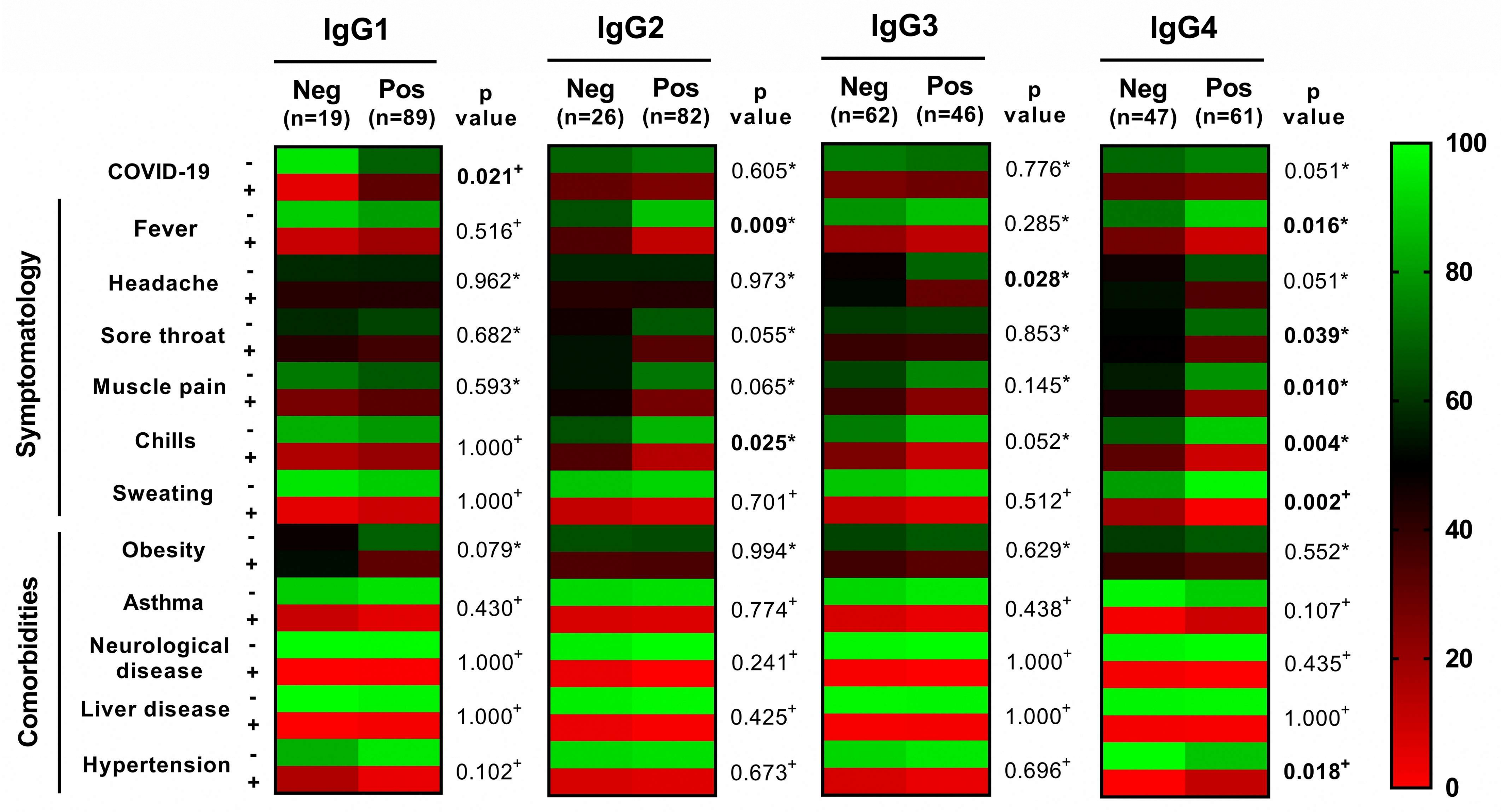
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, D.; Lee, J.-Y.; Yang, J.-S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, C.; Zhong, Q.; Gao, G.F. Overview of SARS-CoV-2 genome-encoded proteins. Sci. China Life Sci. 2022, 65, 280–294. [Google Scholar] [CrossRef]
- Hartenian, E.; Nandakumar, D.; Lari, A.; Ly, M.; Tucker, J.M.; Glaunsinger, B.A. The molecular virology of coronaviruses. J. Biol. Chem. 2020, 295, 12910–12934. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M.G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: The Major Cell Entry Receptor for SARS-CoV-2. Lung 2020, 198, 867–877. [Google Scholar] [CrossRef]
- Kadam, S.B.; Sukhramani, G.S.; Bishnoi, P.; Pable, A.A.; Barvkar, V.T. SARS-CoV-2, the pandemic coronavirus: Molecular and structural insights. J. Basic Microbiol. 2021, 61, 180–202. [Google Scholar] [CrossRef] [PubMed]
- McBride, R.; van Zyl, M.; Fielding, B.C. The Coronavirus Nucleocapsid Is a Multifunctional Protein. Viruses 2014, 6, 2991–3018. [Google Scholar] [CrossRef] [Green Version]
- Ni, L.; Ye, F.; Cheng, M.-L.; Feng, Y.; Deng, Y.-Q.; Zhao, H.; Wei, P.; Ge, J.; Gou, M.; Li, X.; et al. Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals. Immunity 2020, 52, 971–977.e3. [Google Scholar] [CrossRef]
- Burbelo, P.D.; Riedo, F.X.; Morishima, C.; Rawlings, S.; Smith, D.; Das, S.; Strich, J.R.; Chertow, D.S.; Davey, R.T.; Cohen, J.I. Sensitivity in Detection of Antibodies to Nucleocapsid and Spike Proteins of Severe Acute Respiratory Syndrome Coronavirus 2 in Patients With Coronavirus Disease 2019. J. Infect. Dis. 2020, 222, 206–213. [Google Scholar] [CrossRef]
- Guo, L.; Ren, L.; Yang, S.; Xiao, M.; Chang, D.; Yang, F.; Dela Cruz, C.S.; Wang, Y.; Wu, C.; Xiao, Y.; et al. Profiling Early Humoral Response to Diagnose Novel Coronavirus Disease (COVID-19). Clin. Infect. Dis. 2020, 71, 778–785. [Google Scholar] [CrossRef] [Green Version]
- Rak, A.; Donina, S.; Zabrodskaya, Y.; Rudenko, L.; Isakova-Sivak, I. Cross-Reactivity of SARS-CoV-2 Nucleocapsid-Binding Antibodies and Its Implication for COVID-19 Serology Tests. Viruses 2022, 14, 2041. [Google Scholar] [CrossRef]
- Adam, M.H.; Mohamoud, J.H.; Mohamood, A.S.; Mohamed, A.A.; Garba, B.; Dirie, N.I. Seroprevalence of Anti-SARS-CoV-2 Antibodies in Benadir Region, Somalia. Vaccines 2022, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Murhekar, M.V.; Bhatnagar, T.; Thangaraj, J.W.V.; Saravanakumar, V.; Santhosh Kumar, M.; Selvaraju, S.; Rade, K.; Kumar, C.P.G.; Sabarinathan, R.; Asthana, S.; et al. Seroprevalence of IgG antibodies against SARS-CoV-2 among the general population and healthcare workers in India, June–July 2021: A population-based cross-sectional study. PLoS Med. 2021, 18, e1003877. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chen, G.; Jin, Z.; Zhang, Z.; Zhang, B.; He, J.; Yin, S.; Huang, J.; Fan, M.; Li, Z.; et al. Anti-SARS-CoV-2 IgG levels in relation to disease severity of COVID-19. J. Med. Virol. 2022, 94, 380–383. [Google Scholar] [CrossRef]
- Plebani, M.; Padoan, A.; Fedeli, U.; Schievano, E.; Vecchiato, E.; Lippi, G.; Lo Cascio, G.; Porru, S.; Palù, G. SARS-CoV-2 serosurvey in health care workers of the Veneto Region. Clin. Chem. Lab. Med. (CCLM) 2020, 58, 2107–2111. [Google Scholar] [CrossRef]
- Talbot, L.R.; Romeiser, J.L.; Spitzer, E.D.; Gan, T.J.; Singh, S.M.; Fries, B.C.; Bennett-Guerrero, E. Prevalence of IgM and IgG antibodies to SARS-CoV-2 in health care workers at a tertiary care New York hospital during the Spring COVID-19 surge. Perioper Med. 2021, 10, 7. [Google Scholar] [CrossRef]
- Violán, C.; Torán-Monserrat, P.; Quirant, B.; Lamonja-Vicente, N.; Carrasco-Ribelles, L.A.; Chacón, C.; Manresa-Dominguez, J.M.; Ramos-Roure, F.; Dacosta-Aguayo, R.; Palacios-Fernández, C.; et al. Kinetics of humoral immune response over 17 months of COVID-19 pandemic in a large cohort of healthcare workers in Spain: The ProHEpiC-19 study. BMC Infect. Dis. 2022, 22, 721. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Lau, E.H.Y.; Wu, P.; Deng, X.; Wang, J.; Hao, X.; Lau, Y.C.; Wong, J.Y.; Guan, Y.; Tan, X.; et al. Temporal dynamics in viral shedding and transmissibility of COVID-19. Nat. Med. 2020, 26, 672–675. [Google Scholar] [CrossRef] [Green Version]
- Napodano, C.; Marino, M.; Stefanile, A.; Pocino, K.; Scatena, R.; Gulli, F.; Rapaccini, G.L.; Delli Noci, S.; Capozio, G.; Rigante, D.; et al. Immunological Role of IgG Subclasses. Immunol. Investig. 2021, 50, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.W.; Cavacini, L. Structure and function of immunoglobulins. J. Allergy Clin. Immunol. 2010, 125, S41–S52. [Google Scholar] [CrossRef] [Green Version]
- de Taeye, S.W.; Rispens, T.; Vidarsson, G. The Ligands for Human IgG and Their Effector Functions. Antibodies 2019, 8, 30. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Li, T.; Xia, X.; Su, B.; Li, H.; Feng, Y.; Han, J.; Wang, X.; Jia, L.; Bao, Z.; et al. Different Profiles of Antibodies and Cytokines Were Found Between Severe and Moderate COVID-19 Patients. Front. Immunol. 2021, 12, 723585. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Jia, T.; Chen, J.; Zeng, S.; Qiu, Z.; Wu, S.; Li, X.; Lei, Y.; Wang, X.; Wu, W.; et al. The Characterization of Disease Severity Associated IgG Subclasses Response in COVID-19 Patients. Front. Immunol. 2021, 12, 632814. [Google Scholar] [CrossRef] [PubMed]
- Yates, J.L.; Ehrbar, D.J.; Hunt, D.T.; Girardin, R.C.; Dupuis, A.P.; Payne, A.F.; Sowizral, M.; Varney, S.; Kulas, K.E.; Demarest, V.L.; et al. Serological analysis reveals an imbalanced IgG subclass composition associated with COVID-19 disease severity. Cell Rep. Med. 2021, 2, 100329. [Google Scholar] [CrossRef]
- Mutambudzi, M.; Niedzwiedz, C.; Macdonald, E.B.; Leyland, A.; Mair, F.; Anderson, J.; Celis-Morales, C.; Cleland, J.; Forbes, J.; Gill, J.; et al. Occupation and risk of severe COVID-19: Prospective cohort study of 120 075 UK Biobank participants. Occup. Envirion. Med. 2021, 78, 307–314. [Google Scholar] [CrossRef]
- Cortés-Sarabia, K.; Cruz-Rangel, A.; Flores-Alanis, A.; Salazar-García, M.; Jiménez-García, S.; Rodríguez-Martínez, G.; Reyes-Grajeda, J.P.; Rodríguez-Téllez, R.I.; Patiño-López, G.; Parra-Ortega, I.; et al. Clinical features and severe acute respiratory syndrome-coronavirus-2 structural protein-based serology of Mexican children and adolescents with coronavirus disease 2019. PLoS ONE 2022, 17, e0273097. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, J.T.; Hirschhorn, L.R.; Gray, E.L.; Wallia, A.; Carnethon, M.; Zembower, T.R.; Ho, J.; DeYoung, B.J.; Zhu, A.; Rasmussen-Torvik, L.J.; et al. Serologic Status and SARS-CoV-2 Infection over 6 Months of Follow Up in Healthcare Workers in Chicago: A Cohort Study. Infect. Control Hosp. Epidemiol. 2022, 43, 1207–1215. [Google Scholar] [CrossRef]
- Racine-Brzostek, S.E.; Yang, H.S.; Jack, G.A.; Chen, Z.; Chadburn, A.; Ketas, T.J.; Francomano, E.; Klasse, P.J.; Moore, J.P.; McDonough, K.A.; et al. Postconvalescent SARS-CoV-2 IgG and Neutralizing Antibodies are Elevated in Individuals with Poor Metabolic Health. J. Clin. Endocrinol. Metab. 2021, 106, e2025–e2034. [Google Scholar] [CrossRef]
- Radzikowska, U.; Ding, M.; Tan, G.; Zhakparov, D.; Peng, Y.; Wawrzyniak, P.; Wang, M.; Li, S.; Morita, H.; Altunbulakli, C.; et al. Distribution of ACE2, CD147, CD26, and other SARS-CoV-2 associated molecules in tissues and immune cells in health and in asthma, COPD, obesity, hypertension, and COVID-19 risk factors. Allergy 2020, 75, 2829–2845. [Google Scholar] [CrossRef]
- Tan, A.T.; Linster, M.; Tan, C.W.; Le Bert, N.; Chia, W.N.; Kunasegaran, K.; Zhuang, Y.; Tham, C.Y.L.; Chia, A.; Smith, G.J.D.; et al. Early induction of functional SARS-CoV-2-specific T cells associates with rapid viral clearance and mild disease in COVID-19 patients. Cell Rep. 2021, 34, 108728. [Google Scholar] [CrossRef]
- Menni, C.; Valdes, A.M.; Polidori, L.; Antonelli, M.; Penamakuri, S.; Nogal, A.; Louca, P.; May, A.; Figueiredo, J.C.; Hu, C.; et al. Symptom prevalence, duration, and risk of hospital admission in individuals infected with SARS-CoV-2 during periods of omicron and delta variant dominance: A prospective observational study from the ZOE COVID Study. Lancet 2022, 399, 1618–1624. [Google Scholar] [CrossRef]
- Garcia-Basteiro, A.L.; Moncunill, G.; Tortajada, M.; Vidal, M.; Guinovart, C.; Jiménez, A.; Santano, R.; Sanz, S.; Méndez, S.; Llupià, A.; et al. Seroprevalence of antibodies against SARS-CoV-2 among health care workers in a large Spanish reference hospital. Nat. Commun. 2020, 11, 3500. [Google Scholar] [CrossRef]
- Collins, A.M.; Jackson, K.J.L. A Temporal Model of Human IgE and IgG Antibody Function. Front. Immunol. 2013, 4, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stapleton, N.M.; Andersen, J.T.; Stemerding, A.M.; Bjarnarson, S.P.; Verheul, R.C.; Gerritsen, J.; Zhao, Y.; Kleijer, M.; Sandlie, I.; de Haas, M.; et al. Competition for FcRn-mediated transport gives rise to short half-life of human IgG3 and offers therapeutic potential. Nat. Commun. 2011, 2, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG Subclasses and Allotypes: From Structure to Effector Functions. Front. Immunol. 2014, 5, 00520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moncunill, G.; Mayor, A.; Santano, R.; Jiménez, A.; Vidal, M.; Tortajada, M.; Sanz, S.; Méndez, S.; Llupià, A.; Aguilar, R.; et al. SARS-CoV-2 Seroprevalence and Antibody Kinetics Among Health Care Workers in a Spanish Hospital after 3 Months of Follow-up. J. Infect. Dis. 2021, 223, 62–71. [Google Scholar] [CrossRef]
- Whitcombe, A.L.; McGregor, R.; Craigie, A.; James, A.; Charlewood, R.; Lorenz, N.; Dickson, J.M.; Sheen, C.R.; Koch, B.; Fox-Lewis, S.; et al. Comprehensive analysis of SARS-CoV-2 antibody dynamics in New Zealand. Clin. Transl. Immunol. 2021, 10, e1261. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics n (%) | Total (n = 253) | IgG Negative (n = 145) | IgG Positive (n = 108) | p Value |
|---|---|---|---|---|
| Age (Median p25–p75) | 38 (32.5–43.5) | 39 (33–43) | 38 (32–44.5) | 0.7478 ° |
| Sex | ||||
| Female | 192 (75.89) | 115 (79.31) | 77 (71.30) | 0.140 * |
| Male | 61 (24.11) | 30 (20.69) | 31 (28.70) | |
| COVID-19 | ||||
| Negative | 205 (81.03) | 126 (86.90) | 79 (73.15) | 0.006 * |
| Positive | 48 (18.97) | 19 (13.10) | 29 (26.85) | |
| Workplace | ||||
| Doctor | 52 (20.55) | 27 (18.62) | 25 (23.15) | 0.286 * |
| Nurse | 148 (58.50) | 92 (63.45) | 56 (51.85) | |
| Stretcher-bearer | 14 (5.53) | 6 (4.14) | 8 (7.41) | |
| Administrative | 16 (6.32) | 9 (6.21) | 7 (6.48) | |
| Dentist | 2 (0.79) | 0 (0.00) | 2 (1.84) | |
| Psychologist | 6 (2.37) | 2 (1.38) | 4 (3.70) | |
| Social worker | 9 (3.56) | 6 (4.14) | 3 (2.78) | |
| User service | 1 (0.40) | 0 (0.00) | 1 (0.93) | |
| Laboratory | 4 (1.58) | 3 (2.06) | 1 (0.93) | |
| Physiotherapist | 1 (0.40) | 0 (0.00) | 1 (0.93) | |
| Diabetes | ||||
| Negative | 241 (95.26) | 139 (95.86) | 102 (94.44) | 0.600 * |
| Positive | 12 (4.74) | 6 (4.14) | 6 (5.56) | |
| Obesity | ||||
| Negative | 174 (68.77) | 104 (71.72) | 70 (64.81) | 0.241 * |
| Positive | 79 (31.23) | 41 (28.28) | 38 (35.19) | |
| Asthma | ||||
| Negative | 244 (96.44) | 143 (98.62) | 101 (93.52) | 0.040 + |
| Positive | 9 (3.56) | 2 (1.38) | 7 (3.56) | |
| Hypertension | ||||
| Negative | 233 (92.09) | 132 (91.03) | 101 (93.52) | 0.469 * |
| Positive | 20 (7.91) | 13 (8.97) | 7 (6.48) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés-Sarabia, K.; Palomares-Monterrubio, K.H.; Velázquez-Moreno, J.O.; Luna-Pineda, V.M.; Leyva-Vázquez, M.A.; Vences-Velázquez, A.; Dircio-Maldonado, R.; Del Moral-Hernández, O.; Illades-Aguiar, B. Seroprevalence of IgG and Subclasses against the Nucleocapsid of SARS-CoV-2 in Health Workers. Viruses 2023, 15, 955. https://doi.org/10.3390/v15040955
Cortés-Sarabia K, Palomares-Monterrubio KH, Velázquez-Moreno JO, Luna-Pineda VM, Leyva-Vázquez MA, Vences-Velázquez A, Dircio-Maldonado R, Del Moral-Hernández O, Illades-Aguiar B. Seroprevalence of IgG and Subclasses against the Nucleocapsid of SARS-CoV-2 in Health Workers. Viruses. 2023; 15(4):955. https://doi.org/10.3390/v15040955
Chicago/Turabian StyleCortés-Sarabia, Karen, Kenet Hisraim Palomares-Monterrubio, Jesús Omar Velázquez-Moreno, Víctor Manuel Luna-Pineda, Marco Antonio Leyva-Vázquez, Amalia Vences-Velázquez, Roberto Dircio-Maldonado, Oscar Del Moral-Hernández, and Berenice Illades-Aguiar. 2023. "Seroprevalence of IgG and Subclasses against the Nucleocapsid of SARS-CoV-2 in Health Workers" Viruses 15, no. 4: 955. https://doi.org/10.3390/v15040955