

Temporal Dynamic of the Ratio between Monopartite Begomoviruses and Their Associated Betasatellites in Plants, and Its Modulation by the Viral Gene βC1


Abstract
:1. Introduction
2. Material and Methods
2.1. Viruses, Plants and Insects
2.2. Extraction of Total DNA and RNA and Quantification of Begomoviruses, Betasatellites and Gene Expression Level
2.3. Analysis of the Temporal Dynamics of the Quantities of Begomoviruses and Betasatellites and the Expression of AV1 and βC1 in Plants
2.4. Analysis of the Effects of the TbCSB/TbCSV Ratio in Agrobacteria Inoculum on the Quantities of TbCSV and TbCSB and the TbCSB/TbCSV Ratio in Plants
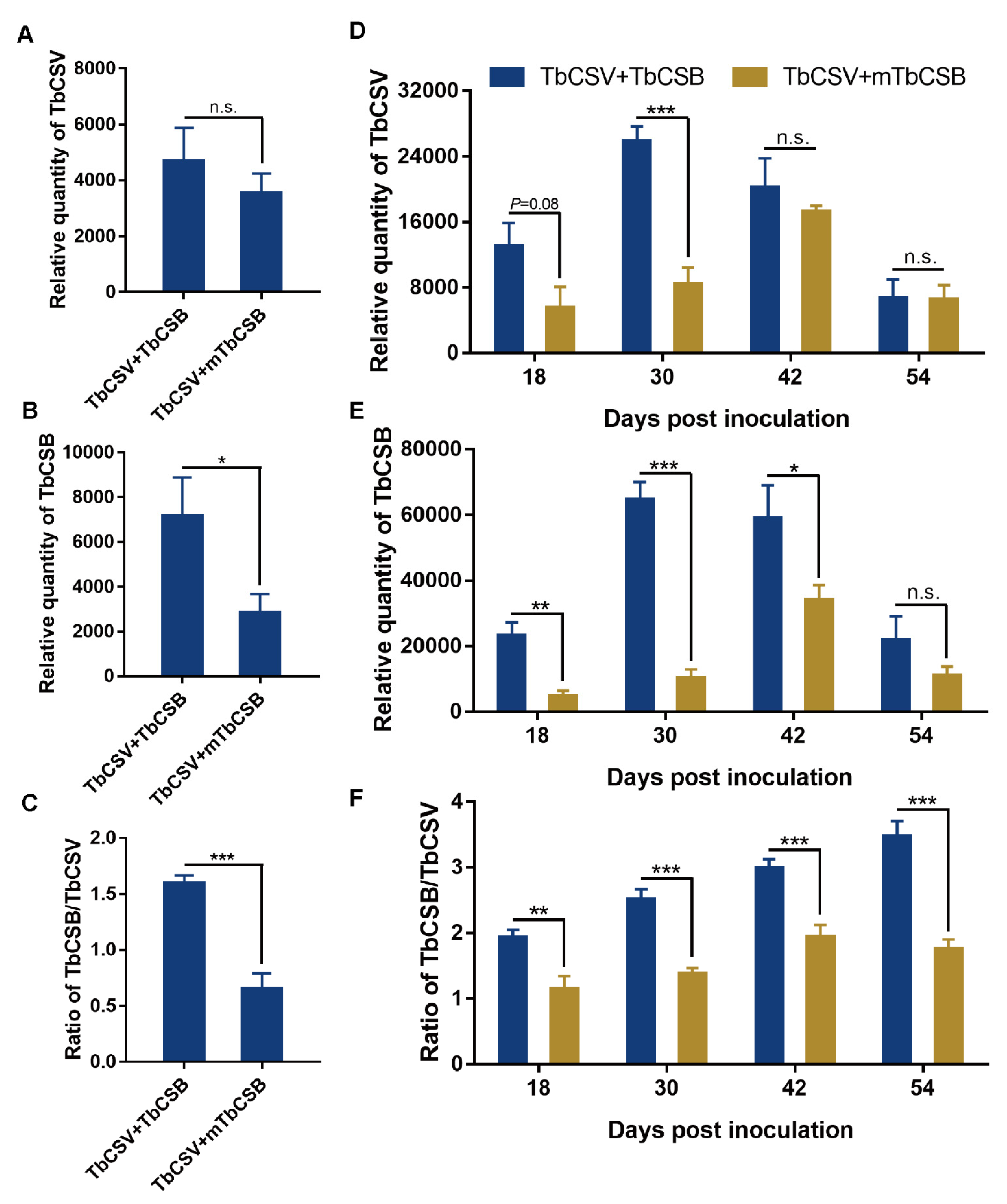
2.5. Analysis of the Effects of βC1 Null-Mutation on the Quantities of Begomoviruses and Betasatellites and the Betasatellite/Begomovirus Ratio
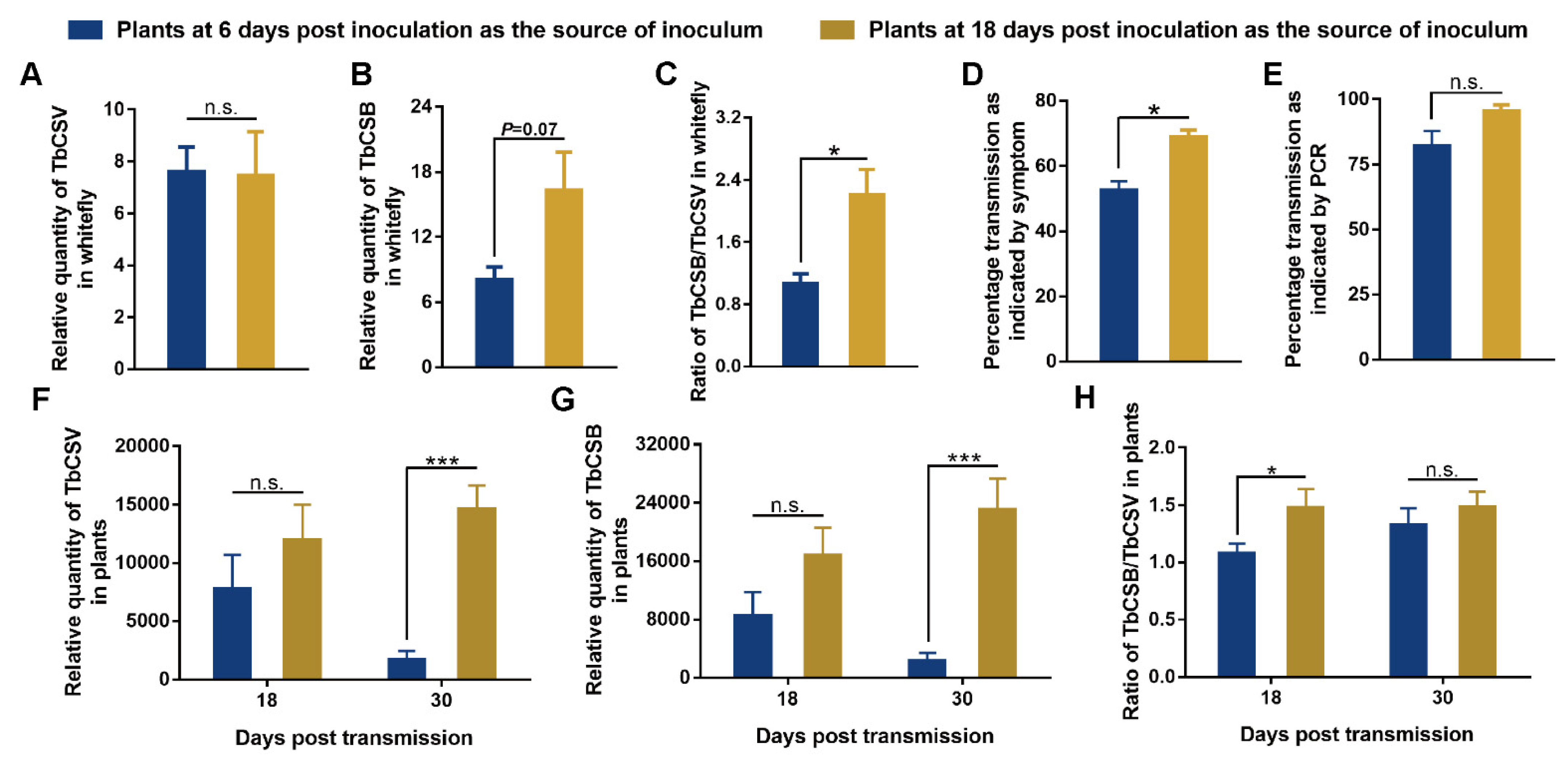
2.6. Analysis of the Effects of TbCSB/TbCSV Ratio in the Plants as Source of Inoculum on Whitefly-Mediated Virus Transmission
2.7. Statistical Analysis
3. Results
3.1. Symptoms Induced by TbCSV and TbCSB in Common Tobacco Plants
3.2. Temporal Dynamics of the Quantities of TbCSV and TbCSB and the TbCSB/TbCSV Ratio in Plants
3.3. Effects of the TbCSB/TbCSV Ratio in Agrobacteria Inoculum on the Quantities of TbCSV and TbCSB and the TbCSB/TbCSV Ratio in Plants
3.4. Effects of βC1 Null-Mutation on the Quantities of TbCSV and TbCSB and the TbCSB/TbCSV Ratio in Common Tobacco Plants
3.5. Effects of the TbCSB/TbCSV Ratio in the Plants as Source of Inoculum on Whitefly-Mediated Virus Transmission
3.6. Temporal Dynamics of the Expression of TbCSV AV1 and TbCSB βC1 and the βC1/AV1 Ratio in Common Tobacco Plants
3.7. Temporal Dynamics of the Quantities of CLCuMuV and CLCuMuB and the CLCuMuB/CLCuMuV Ratio in Plants and Their Response to βC1 Null-Mutation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, R.A.C. Global plant virus disease pandemics and epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef]
- Navas-Castillo, J.; Fiallo-Olivé, E.; Sánchez-Campos, S. Emerging virus diseases transmitted by whiteflies. Annu. Rev. Phytopathol. 2011, 49, 219–248. [Google Scholar] [CrossRef] [PubMed]
- Rojas, M.R.; Macedo, M.A.; Maliano, M.R.; Soto-Aguilar, M.; Souza, J.O.; Briddon, R.W.; Kenyon, L.; Bustamante, R.F.R.; Zerbini, F.M.; Adkins, S.; et al. World management of geminiviruses. Annu. Rev. Phytopathol. 2018, 56, 637–677. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Navas-Castillo, J. Begomoviruses: What is the secret(s) of their success? Trends Plant Sci. 2023, in press. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Lett, J.M.; Martin, D.P.; Roumagnac, P.; Varsani, A.; Zerbini, F.M.; Navas-Castillo, J.; ICTV Report Consortium. ICTV virus taxonomy profile: Geminiviridae 2021. J. Gen. Virol. 2021, 102, 001696. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://ictv.global/report/chapter/geminiviridae/geminiviridae (accessed on 12 December 2022).
- Brown, J.K.; Zerbini, F.M.; Navas-Castillo, J.; Moriones, E.; Ramos-Sobrinho, R.; Silva, J.C.F.; Fiallo-Olivé, E.; Briddon, R.W.; Hernández-Zepeda, C.; Idris, A.; et al. Revision of Begomovirus taxonomy based on pairwise sequence comparisons. Arch. Virol. 2015, 160, 1593–1619. [Google Scholar] [CrossRef]
- Fiallo-Olivé, E.; Pan, L.L.; Liu, S.S.; Navas-Castillo, J. Transmission of begomoviruses and other whitefly-borne viruses: Dependence on the vector species. Phytopathology 2020, 110, 10–17. [Google Scholar] [CrossRef]
- Wang, X.W.; Blanc, S. Insect transmission of plant single-stranded DNA viruses. Annu. Rev. Èntomol. 2021, 66, 389–405. [Google Scholar] [CrossRef]
- Zhou, X.P. Advances in understanding begomovirus satellites. Annu. Rev. Phytopathol. 2013, 51, 357–381. [Google Scholar] [CrossRef]
- Gnanasekaran, P.; Chakraborty, S. Biology of viral satellites and their role in pathogenesis. Curr. Opin. Virol. 2018, 33, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Badar, U.; Venkataraman, S.; AbouHaidar, M.; Hefferon, K. Molecular interactions of plant viral satellites. Virus Genes 2020, 57, 1–22. [Google Scholar] [CrossRef]
- Aimone, C.D.; De León, L.; Dallas, M.M.; Ndunguru, J.; Ascencio-Ibáñez, J.T.; Hanley-Bowdoin, L. A new type of satellite associated with cassava mosaic begomoviruses. J. Virol. 2021, 95, e00432-21. [Google Scholar] [CrossRef] [PubMed]
- Gnanasekaran, P.; KishoreKumar, R.; Bhattacharyya, D.; VinothKumar, R.; Chakraborty, S. Multifaceted role of geminivirus associated betasatellite in pathogenesis. Mol. Plant Pathol. 2019, 20, 1019–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.L.; Guo, W.; Li, F.F.; Sunter, G.; Zhou, X.P. Geminivirus-associated betasatellites: Exploiting chinks in the antiviral arsenal of plants. Trends Plant Sci. 2019, 24, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Idris, A.M.; Mansoor, S.; Bedford, I.D.; Dhawan, P.; Rishi, N.; Siwatch, S.S.; Abdel-Salam, A.M.; et al. Diversity of DNAβ, a satellite molecule associated with some monopartite begomoviruses. Virology 2003, 312, 106–121. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Song, Y.; Wang, Y.Q.; Zhou, X.P. Functional analysis of a novel βV1 gene identified in a geminivirus betasatellite. Sci. China Life Sci. 2020, 63, 688–696. [Google Scholar] [CrossRef]
- Voorburg, C.M.; Yan, Z.; Bergua-Vidal, M.; Wolters, A.M.A.; Bai, Y.L.; Kormelink, R. Ty-1, a universal resistance gene against geminiviruses that is compromised by co-replication of a betasatellite. Mol. Plant Pathol. 2020, 21, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Li, F.F.; Yang, X.L.; Bisaro, D.M.; Zhou, X.P. The βC1 protein of geminivirus-betasatellite complexes: A target and repressor of host defenses. Mol. Plant 2018, 11, 1424–1426. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Zhou, X.P.; Zhang, Z.K.; Qi, Y.J. Tobacco curly shoot virus isolated in Yunnan is a distinct species of Begomovirus. Chin. Sci. Bull. 2002, 47, 197–200. [Google Scholar] [CrossRef]
- Li, Z.H.; Xie, Y.; Zhou, X.P. Tobacco curly shoot virus DNAβ is not necessary for infection but intensifies symptoms in a host-dependent manner. Phytopathology 2005, 95, 902–908. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.H.; Hu, G.J.; Xie, Y.; Zhou, X.P. Construction and pathogenicity evaluation of infectious clones of cotton leaf curl Multan virus infecting Hibiscus rosa-sinensis and Gossypium hirtusum. Acta Phytopathol. Sin. 2015, 45, 198–204, (In Chinese with English Abstract). [Google Scholar]
- Pan, L.L.; Chen, Q.F.; Guo, T.; Wang, X.R.; Li, P.; Wang, X.W.; Liu, S.S. Differential efficiency of a begomovirus to cross the midgut of different species of whiteflies results in variation of virus transmission by the vectors. Sci. China Life Sci. 2018, 61, 1254–1265. [Google Scholar] [CrossRef]
- Ruan, Y.M.; Luan, J.B.; Zang, L.S.; Liu, S.S. Observing and recording copulation events of whiteflies on plants using a video camera. Èntomol. Exp. Appl. 2007, 124, 229–233. [Google Scholar] [CrossRef]
- Conflon, D.; Granier, M.; Tiendrébéogo, F.; Gentite, P.; Peterschmitt, M.; Urbino, C. Accumulation and transmission of alphasatellite, betasatellite and tomato yellow leaf curl virus in susceptible and Ty-1-resistant tomato plants. Virus Res. 2018, 253, 124–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicard, A.; Michalakis, Y.; GutieÂrrez, S.; Blanc, S. The strange lifestyle of multipartite viruses. PLoS Pathog. 2016, 12, e1005819. [Google Scholar] [CrossRef] [Green Version]
- Michalakis, Y.; Blanc, S. The curious strategy of multipartite viruses. Annu. Rev. Virol. 2020, 7, 203–218. [Google Scholar] [CrossRef]
- Sicard, A.; Yvon, M.; Timchenko, T.; Gronenborn, B.; Michalakis, Y.; Gutierrez, S.; Blanc, S. Gene copy number is differentially regulated in a multipartite virus. Nat. Commun. 2013, 4, 2248. [Google Scholar] [CrossRef] [Green Version]
- Moreau, Y.; Gil, P.; Exbrayat, A.; Rakotoarivony, I.; Bréard, E.; Sailleau, C.; Viarouge, C.; Zientara, S.; Savini, G.; Goffredo, M.; et al. The genome segments of bluetongue virus differ in copy number in a host-specific manner. J. Virol. 2021, 95, e01834-20. [Google Scholar] [CrossRef]
- Gallet, R.; Mattia, J.D.; Ravel, S.; Zeddam, J.; Vitalis, R.; Michalakis, Y.; Blanc, S. Gene copy number variations at the within-host population level modulate gene expression in a multipartite virus. Virus Evol. 2022, 8, veac058. [Google Scholar] [CrossRef] [PubMed]
- Kon, T.; Gilbertson, R.L. Two genetically related begomoviruses causing tomato leaf curl disease in Togo and Nigeria differ in virulence and host range but do not require a betasatellite for induction of disease symptoms. Arch. Virol. 2012, 157, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Zaidi, S.S.; Shakir, S.; Amin, I.; Mansoor, S. An insight into cotton leaf curl multan Multan betasatellite, the most important component of cotton leaf curl disease complex. Viruses 2017, 9, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mubin, M.; Ijaz, S.; Nahid, N.; Hassan, M.; Younus, A.; Qazi, J.; Nawaz-ul-Rehman, M.S. Journey of begomovirus betasatellite molecules: From satellites to indispensable partners. Virus Genes 2020, 56, 16–26. [Google Scholar] [CrossRef]
- Rojas, M.R.; Hagen, C.; Lucas, W.J.; Gilbertson, R.L. Exploiting chinks in the plant’s armor: Evolution and emergence of geminiviruses. Annu. Rev. Phytopathol. 2005, 43, 361–394. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.F.; Tao, X.R.; Xie, Y.; Fauquet, C.M.; Zhou, X.P. A DNA beta associated with tomato yellow leaf curl china virus is required for symptom induction. J. Virol. 2005, 78, 13966–13974. [Google Scholar] [CrossRef] [Green Version]
- Eini, O.; Behjatnia, S.A.A. The minimal sequence essential for replication and movement of cotton leaf curl Multan betasatellite DNA by a helper virus in plant cells. Virus Genes 2016, 52, 679–687. [Google Scholar] [CrossRef]
- Zhang, T.; Xu, X.B.; Huang, C.J.; Qian, Y.J.; Li, Z.H.; Zhou, X.P. A novel DNA motif contributes to selective replication of a geminivirus-associated betasatellite by a helper virus-encoded replication-related protein. J. Virol. 2016, 90, 2077–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Application |
|---|---|---|
| TbCSV-AV1-RTF | GAAGCGTCCAGCAGATATAA | Quantification of TbCSV and AV1 |
| TbCSV-AV1-RTR | GGGACATCAGGACTTCTGTA | |
| TbCSB-βC1-RTF | AATCACCAGCACTGGCAAAG | Quantification of TbCSb and βC1 |
| TbCSB-βC1-RTR | CCGCTTCTTGCATCATCAGG | |
| Nt-Actin-RTF | GTGTTAGCCACACTGTCCCA | Quantification of N. tabacum actin |
| Nt-Actin-RTR | TCAGTCAAGTCACGACCAGC | |
| Nb-Actin-RTF | GCGAGTAAACCCGTAAGG | Quantification of N. benthamiana actin |
| Nb-Actin-RTR | GCTCAGGCATAGTTCACC | |
| TbCSV-A-PCRF | ACAGAAGTCCTGATGTCCCT | PCR detection of TbCSV |
| TbCSV-A-PCRR | AGAGCACCAGAACCGTCC | |
| CLCuMuV-AV1-RTF | ACACTTGTGCAGTCCCAGAG | Quantification of CLCuMuV and AV1 |
| CLCuMuV-AV1-RTR | CACTTCAACCGTCCATTCAC | |
| CLCuMuB-βC1-RTF | AGCCGTTGAAGTCGAATGGA | Quantification of CLCuMuB and βC1 |
| CLCuMuB-βC1-RTR | GACGAGGAGCAGAACAAACA | |
| TbCSB-FLF | CCGGAATTCGAAACCACTACGCTACGCAG | Cloning of TbCSB |
| TbCSB-FLR | CGGGGTACCTACCCTCCCAGGGGTACAC | |
| TbCSB-TBF | TGTTGTATTTAATTGTCTAATTTGTTCT | Mutagenesis of TbCSB |
| TbCSB-TBR | TAGACAATTAAATACAACAACAAGAAGGGC | |
| CLCuMuB-FLF | CGGGGTACCACTACGCTACGCAGCAGCC | Cloning of CLCuMuB |
| CLCuMuB-FLR | CGGGGTACCTACCCTCCCAGGGGTACAC | |
| CLCuMuB-TBF | CGGGGTACCTACCCTCCCAGGGGTACAC | Mutagenesis of CLCuMuB |
| CLCuMuB-TBR | TAGACGAGGAGCAGAACAAACACGCAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-J.; Liu, Y.-M.; Li, H.-Y.; Liu, S.-S.; Pan, L.-L. Temporal Dynamic of the Ratio between Monopartite Begomoviruses and Their Associated Betasatellites in Plants, and Its Modulation by the Viral Gene βC1. Viruses 2023, 15, 954. https://doi.org/10.3390/v15040954
Wu Y-J, Liu Y-M, Li H-Y, Liu S-S, Pan L-L. Temporal Dynamic of the Ratio between Monopartite Begomoviruses and Their Associated Betasatellites in Plants, and Its Modulation by the Viral Gene βC1. Viruses. 2023; 15(4):954. https://doi.org/10.3390/v15040954
Chicago/Turabian StyleWu, Yi-Jie, Yi-Ming Liu, Heng-Yu Li, Shu-Sheng Liu, and Li-Long Pan. 2023. "Temporal Dynamic of the Ratio between Monopartite Begomoviruses and Their Associated Betasatellites in Plants, and Its Modulation by the Viral Gene βC1" Viruses 15, no. 4: 954. https://doi.org/10.3390/v15040954