
Prevalences of Tobamovirus Contamination in Seed Lots of Tomato and Capsicum

Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Incidences of Contamination in Smaller Seed Lots
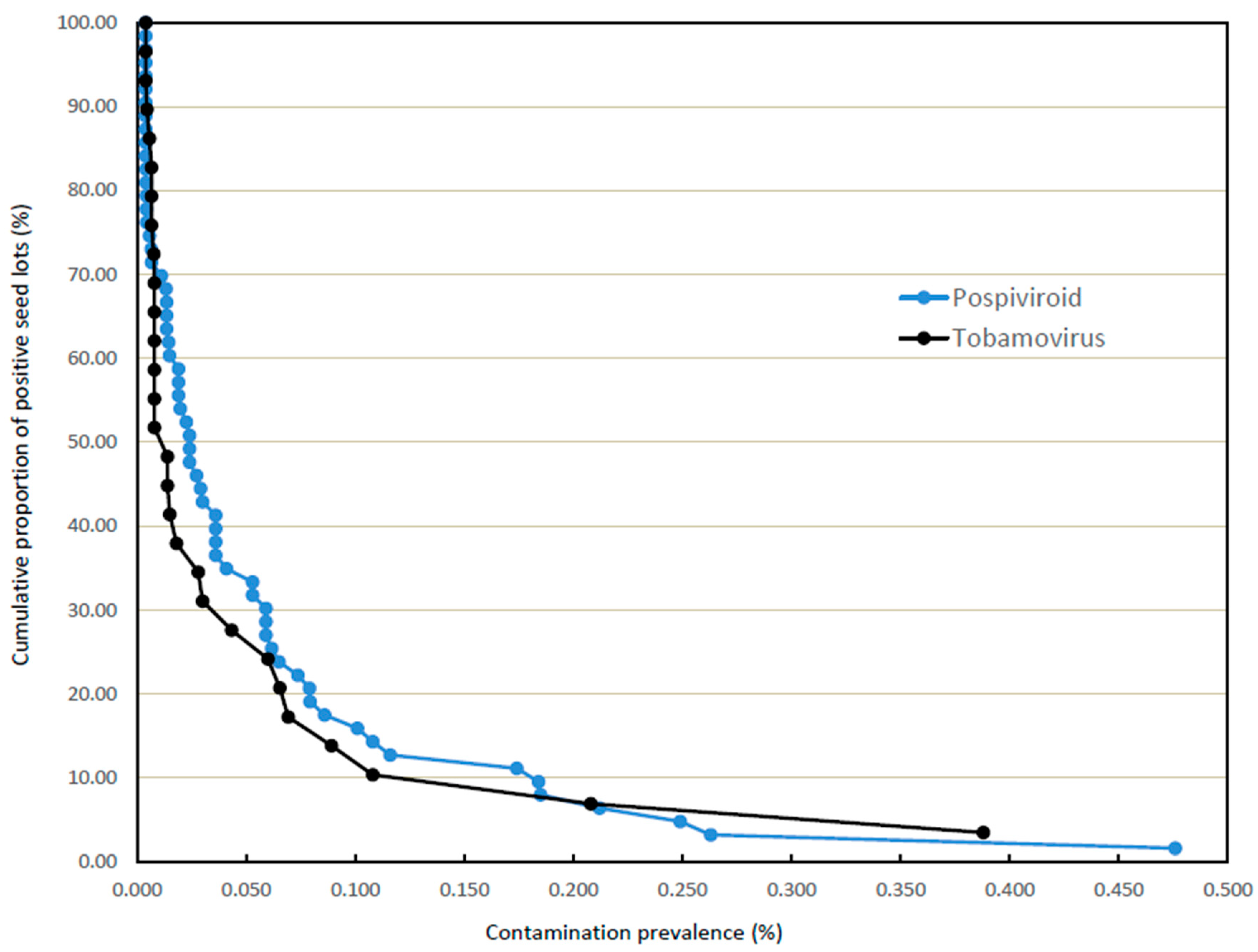
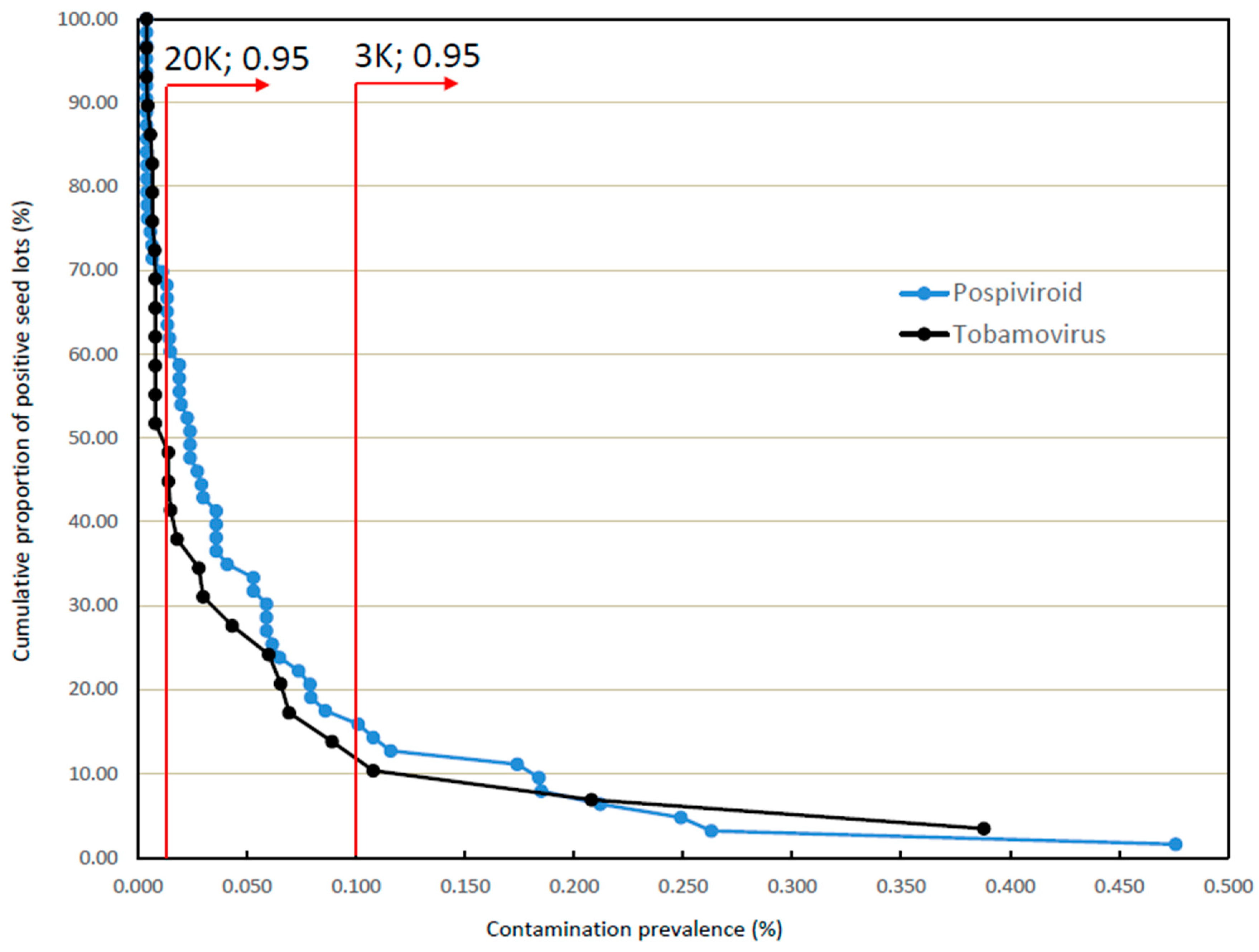
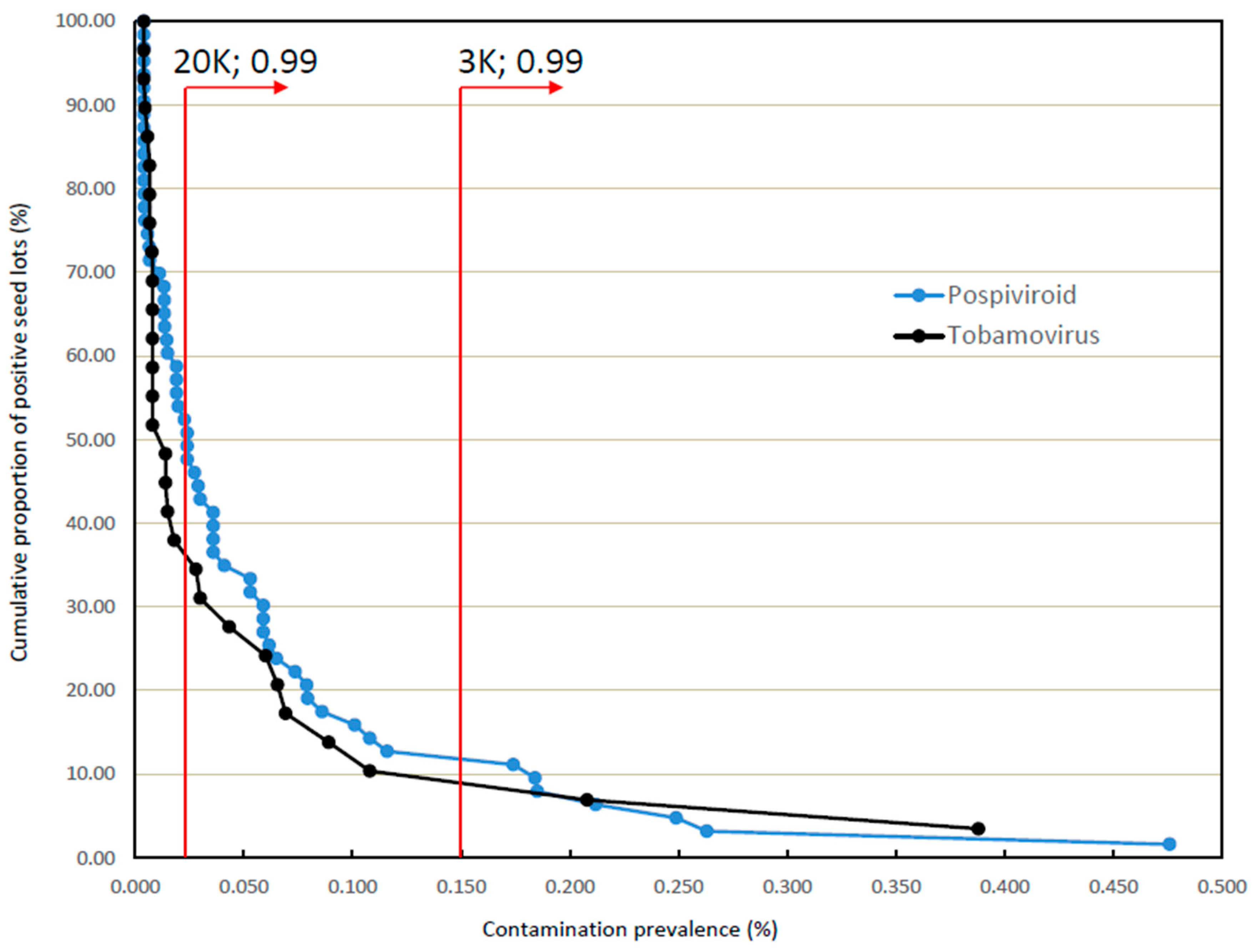
3.2. Incidences and Prevalences of Contamination in Larger Seed Lots
- For a given pest species, jurisdictions that impose more rigorous at-border testing regimes for seed contamination should record fewer incidences of post-border pest incursions than those using less rigorous testing regimes; and
- For a given jurisdiction, pests that are subject to more rigorous at-border testing regimes should display lower incidences of post-border incursion than those subject to less rigorous regimes.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davino, S.; Caruso, A.G.; Bertacca, S.; Barone, S.; Panno, S. Tomato Brown Rugose Fruit Virus: Seed transmission rate and efficacy of different seed disinfection treatments. Plants 2020, 9, 1615. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.M.; Sulaiman, A.; Samarah, N.; Turina, M.; Vallino, M. Localization and mechanical transmission of tomato brown rugose fruit virus in tomato seeds. Plant Dis. 2022, 106, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Dombrovsky, A.; Smith, E. Seed transmission of tobamoviruses: Aspects of global disease distribution. In Advances in Seed Biology; Jimenez-Lopez, J.C., Ed.; Intech Open: London, UK, 2017; pp. 234–260. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, D.; Mintoff, S.; Kurz, N.; Neilsen, M.; Patel, S.; Constable, F.; Tran-Nguyen, L. Investigating the Longevity and Infectivity of Cucumber green mottle mosaic virus in Soils of the Northern Territory, Australia. Plants 2022, 11, 883. [Google Scholar] [CrossRef] [PubMed]
- International Seed Federation. Detection of Infectious Tomato Brown Rugose Fruit Virus (ToBRFV) in Tomato and Pepper Seed; International Seed Federation: Nyon, Switzerland, 2019. [Google Scholar]
- International Seed Testing Association. International Rules for Seed Testing 2019 7-028: Detection of Infectious Tobacco Mosaic Virus and Tomato Mosaic Virus in Solanum Lycopersicum (Tomato) Seed by the Local Lesion Assay (Indexing) on Nicotiana Tabacum Plants; International Seed Testing Association: Bassersdorf, Switzerland, 2019. [Google Scholar]
- Constable, F.; Chambers, G.; Penrose, L.; Daly, A.; Mackie, J.; Davis, K.; Rodoni, B.; Gibbs, M. Viroid-infected tomato and capsicum seed shipments to Australia. Viruses 2019, 11, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall, D.; Penrose, L.; Daly, A.; Constable, F.; Gibbs, M. Prevalences of pospiviroid contamination in large seed lots of tomato and capsicum, and related seed testing considerations. Viruses 2019, 11, 1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelock, D.A.; Kinoti, W.M.; Bottcher, C.; Wildman, O.; Dall, D.; Rodoni, B.C.; Constable, F.E. Tomato mottle mosaic virus intercepted by Australian biosecurity in Capsicum annuum seed. Australas. Plant Dis. Notes 2020, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Levitzky, N.; Smith, E.; Lachman, O.; Luria, N.; Mizrahi, Y.; Bakelman, H.; Sela, N.; Laskar, O.; Milrot, E.; Dombrovsky, A. The bumblebee Bombus terrestris carries a primary inoculum of Tomato brown rugose fruit virus contributing to disease spread in tomatoes. PLoS ONE 2019, 14, e0210871. [Google Scholar] [CrossRef] [PubMed]
- European Plant Protection Organisation. PRA—Express PRA for Tomato Brown Rugose Fruit Virus (eppo.int). 2019. Available online: https://pra.eppo.int/pra/bd40c6b4-64bf-4dc3-964e-a94b8c80934e (accessed on 13 March 2023).
- European Plant Protection Organisation. PRA—Quick Scan for Tomato Mottle Mosaic Virus (eppo.int). 2020. Available online: https://pra.eppo.int/pra/991b6c2c-1a97-4b5f-93eb-d5f141c61e36 (accessed on 13 March 2023).
- Kitajima, M.; Sassi, H.P.; Torrey, J.R. Pepper mild mottle virus as a water quality indicator. NPJ Clean Water 2018, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Fowkes, A.R.; Botermans, M.; Frew, L.; de Koning, P.P.; Buxton-Kirk, A.; Westenberg, M.; Ward, R.; Schenk, M.; Webster, G.; Alraiss, K.; et al. First report of Tomato mottle mosaic virus in Solanum lycopersicum seeds in The Netherlands and intercepted in seed imported from Asia. New Dis. Rep. 2022, 45, e12067. [Google Scholar] [CrossRef]
- McKenna, C.; Morrison, A. Guide 3: Natural Experiments; Scottish Government Social Research Group Social Science Methods Series. Social Research: Methods guides. Available online: https://www.gov.scot/publications/social-research-methods-guides/ (accessed on 13 March 2023).




{kind=link}
{kind=link}
{kind=link}
| Virus Identity a. | Tomato | Capsicum | Combined | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ≤400 b | 800–2000 b | 2400–8000 b | ≤400 a | 800–2000 | 2400–8000 | ≤400 a | 800–2000 | 2400–8000 | |
| PMMoV | 1 | 2 | 2 | 8 | 5 | 7 | 9 | 7 | 9 |
| TMV | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 |
| ToMV | 6 | 10 | 4 | 0 | 2 | 3 | 6 | 12 | 7 |
| ToBRFV c | 23 | 4 | 3 | 0 | 0 | 2 | 23 | 4 | 5 |
| ToMMV c | 25 | 4 | 2 | 6 | 0 | 2 | 31 | 4 | 4 |
| Total detections | 55 | 20 | 12 | 14 | 8 | 14 | 69 | 28 | 26 |
| Total tested | 217 | 140 | 74 | 74 | 91 | 63 | 291 | 231 | 137 |
| % contaminated | 25.3 | 14.3 | 16.2 | 18.9 | 8.8 | 22.0 | 23.7 | 12.1 | 19.0 |
| Tomato | Capsicum | Combined | ||||
|---|---|---|---|---|---|---|
| Virus Identity a | Total Detections | % Contaminated | Total Detections | % Contaminated | Total Detections | % Contaminated |
| PMMoV | 1 | 1.4 | 10 | 21.3 | 11 | 9.3 |
| TMV | 0 | – | 1 | 2.1 | 1 | 0.8 |
| ToMV | 7 | 9.9 | 8 | 17.0 | 15 | 12.7 |
| ToMMV b | 3 | 4.2 | 1 | 2.1 | 4 | 3.4 |
| Total detections | 11 | 15.5 | 20 | 42.6 | 31 | 26.3 |
| Total tested | 71 | 47 | 118 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dall, D.J.; Lovelock, D.A.; Penrose, L.D.J.; Constable, F.E. Prevalences of Tobamovirus Contamination in Seed Lots of Tomato and Capsicum. Viruses 2023, 15, 883. https://doi.org/10.3390/v15040883
Dall DJ, Lovelock DA, Penrose LDJ, Constable FE. Prevalences of Tobamovirus Contamination in Seed Lots of Tomato and Capsicum. Viruses. 2023; 15(4):883. https://doi.org/10.3390/v15040883
Chicago/Turabian StyleDall, David J., David A. Lovelock, Lindsay D. J. Penrose, and Fiona E. Constable. 2023. "Prevalences of Tobamovirus Contamination in Seed Lots of Tomato and Capsicum" Viruses 15, no. 4: 883. https://doi.org/10.3390/v15040883