
Development of an Indirect Enzyme-Linked Immunosorbent Assay Based on the Yeast-Expressed CO-26K-Equivalent Epitope-Containing Antigen for Detection of Serum Antibodies against Porcine Epidemic Diarrhea Virus
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Vectors, Cells, and Sera
2.2. Phylogenetic and Conservative Analysis
2.3. Construction of Recombinant pPIC9K-COE Plasmid
2.4. Expression and Purification of the Recombinant COE Protein
2.5. Western Blotting
2.6. Confirmation of the Serum Samples
2.7. Establishment of the COE-iELISA Method
2.8. Determination of the Cut-Off Value
2.9. Cross-Reactivity and Repeatability Assay
2.10. Detection of Vaccinated Serum Samples by COE-iELISA
3. Results
3.1. Phylogenetic and Conservative Analysis of PEDVs
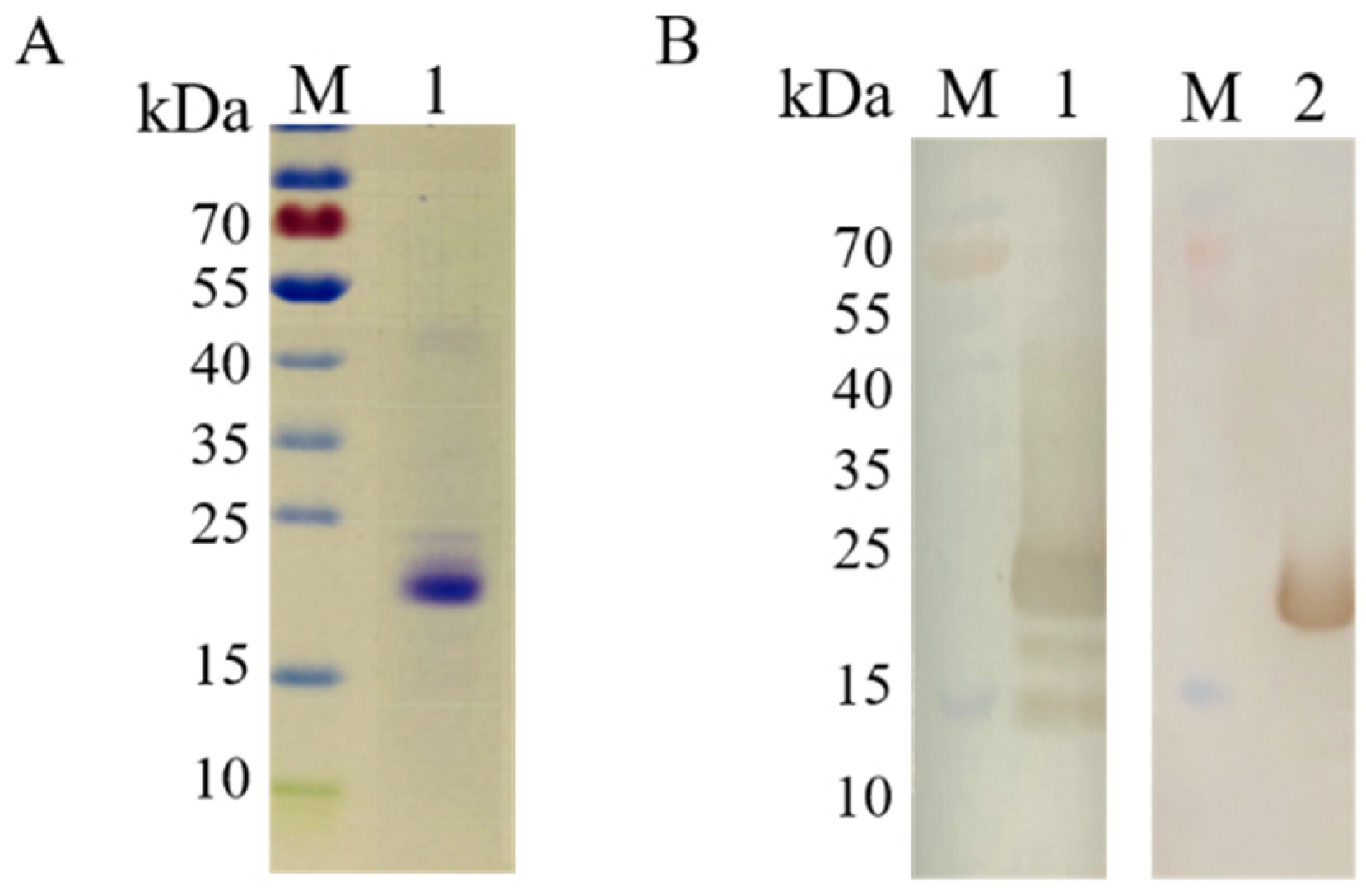
3.2. Expression and Identification of Recombinant COE Protein
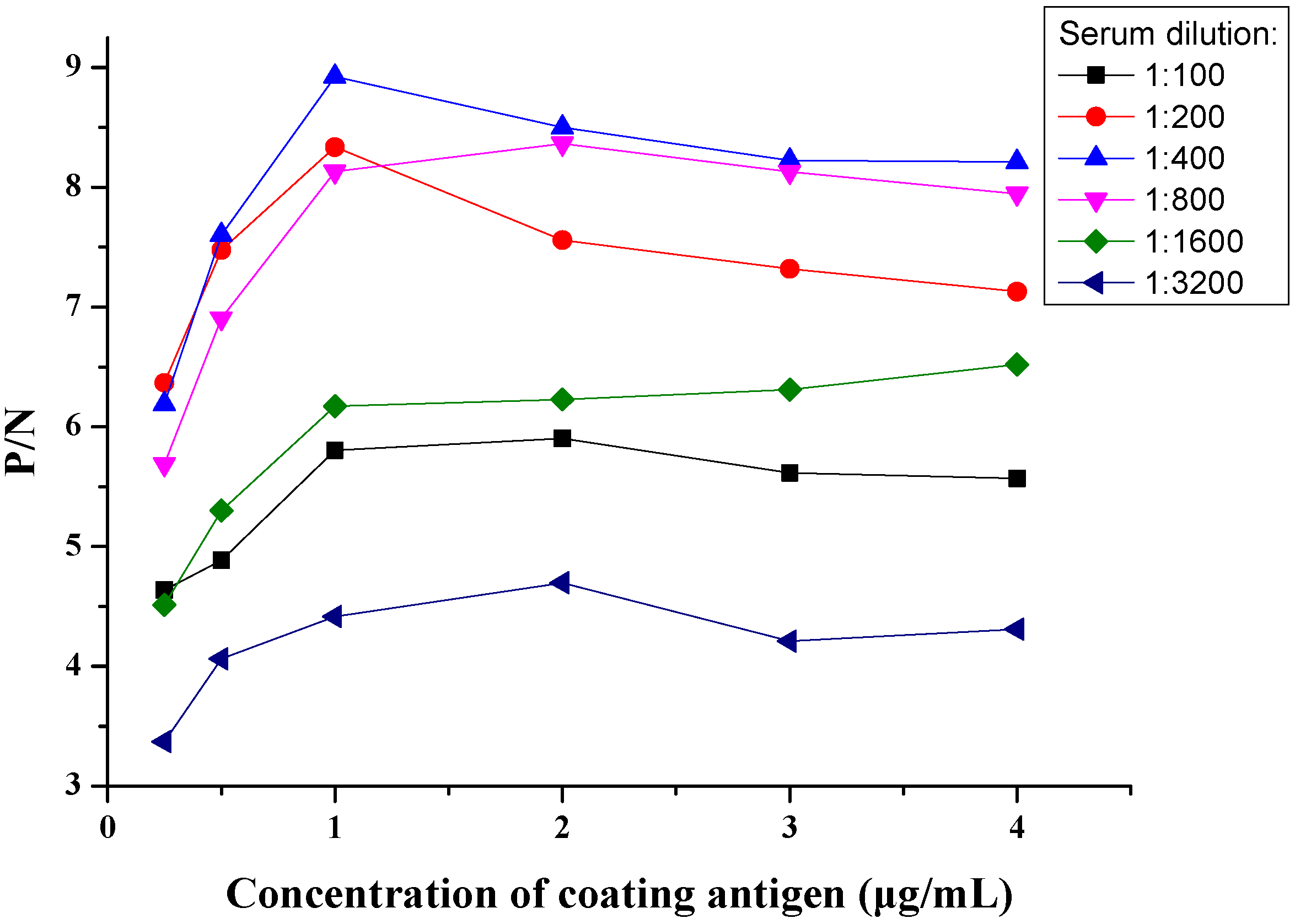
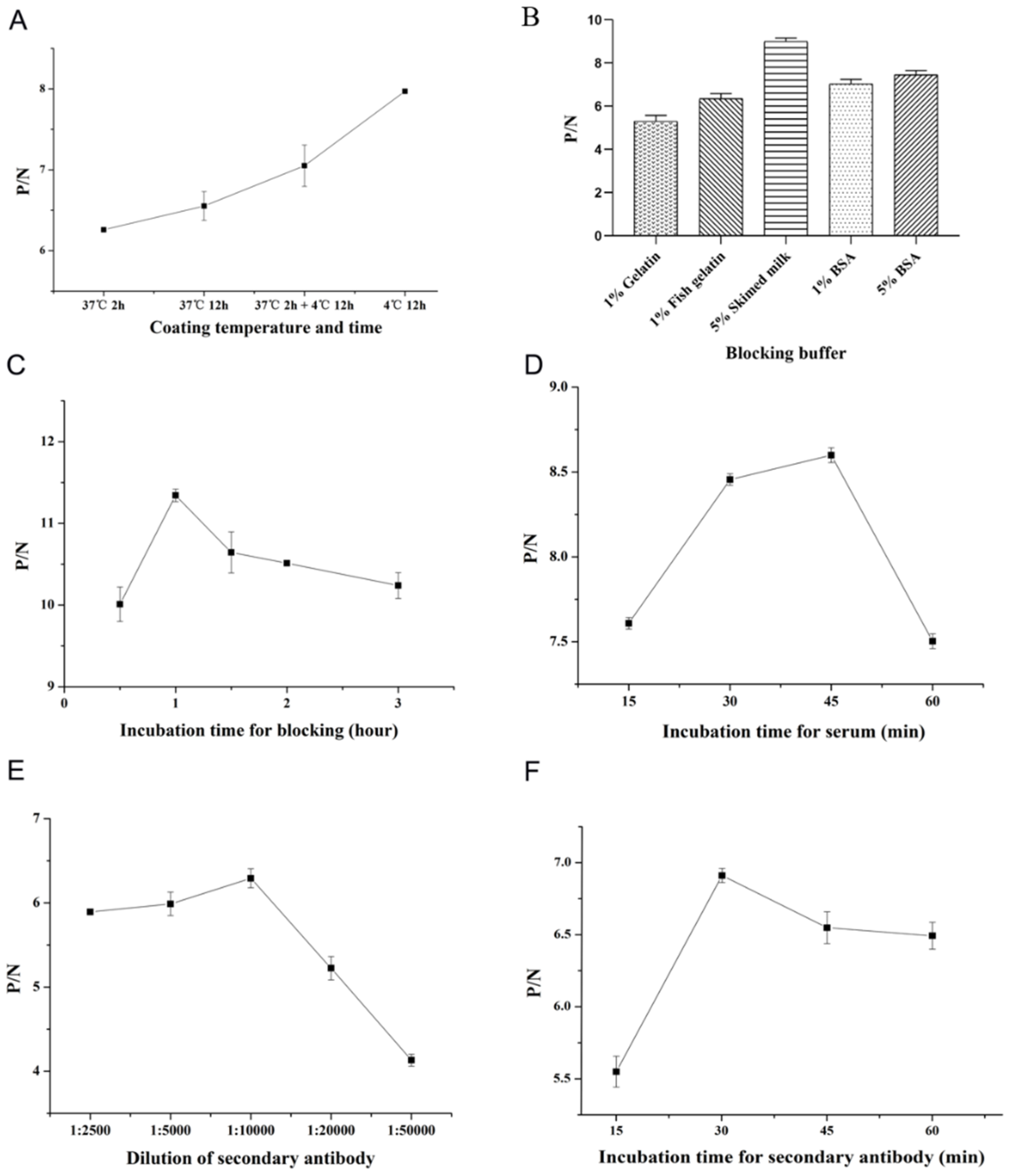
3.3. Optimization of COE-iELISA Reaction Conditions
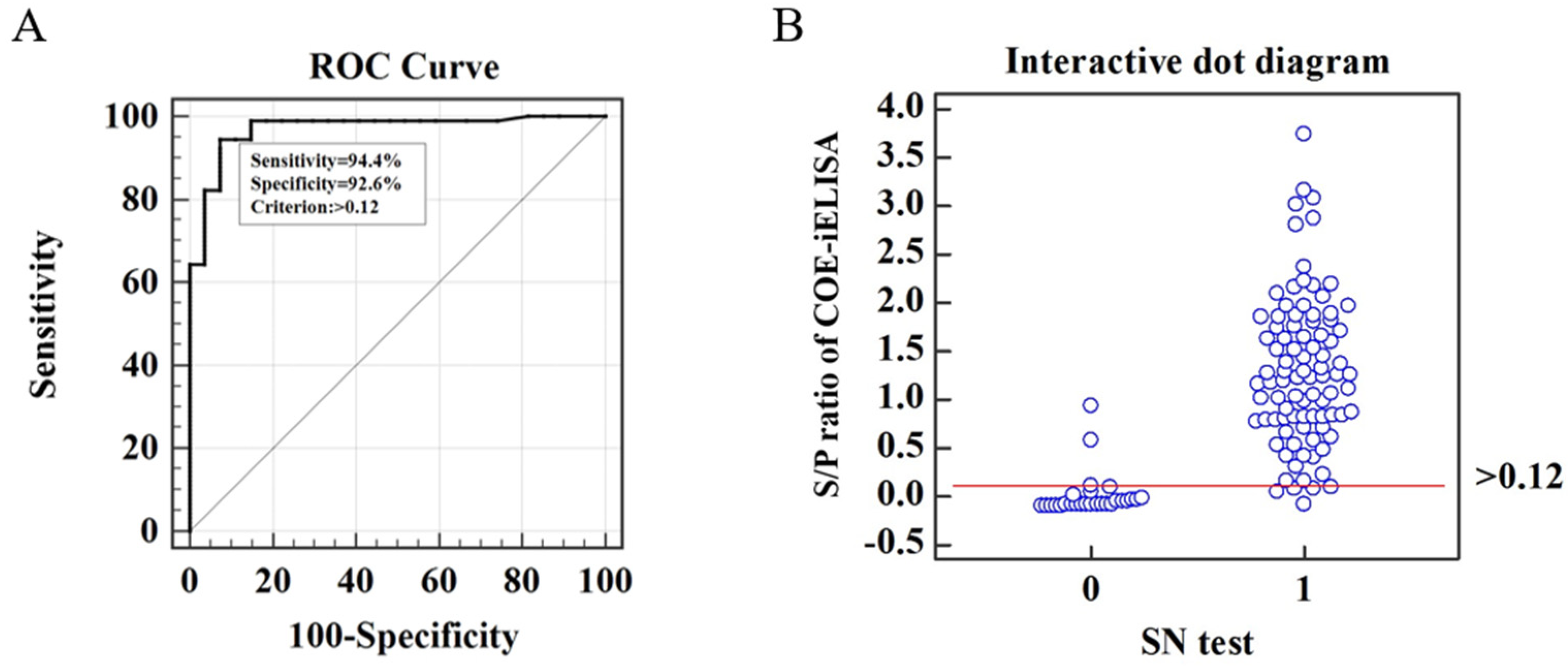
3.4. Cut-Off Value of COE-iELISA
3.5. Cross-Reactivity and Repeatability of COE-iELISA
3.6. Detection of Vaccinated Serum Samples of COE-iELISA
3.7. Clinical Application
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gonzalez, J.M.; Gomez-Puertas, P.; Cavanagh, D.; Gorbalenya, A.; Enjuanes, L. A comparative sequence analysis to revise the current taxonomy of the family Coronaviridae. Arch. Virol. 2003, 148, 2207–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Fang, L.; Xiao, S. Porcine epidemic diarrhea in China. Virus Res. 2016, 226, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Wood, E. An apparently new syndrome of porcine epidemic diarrhoea. Vet. Rec. 1977, 100, 243–244. [Google Scholar] [CrossRef]
- Gerber, P.F.; Gong, Q.; Huang, Y.-W.; Wang, C.; Holtkamp, D.; Opriessnig, T. Detection of antibodies against porcine epidemic diarrhea virus in serum and colostrum by indirect ELISA. Vet. J. 2014, 202, 33–36. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, T.; Li, Z.; Guo, X.; Tian, Y.; Li, Y.; Xiao, S. A novel biotinylated nanobody-based blocking ELISA for the rapid and sensitive clinical detection of porcine epidemic diarrhea virus. J. Nanobiotechnol. 2019, 17, 96. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Hu, Z.; Fan, M.; Shao, Z.; Yu, Q.; Li, X. Development of an indirect ELISA to detect PEDV specific IgA antibody based on a PEDV epidemic strain. BMC Vet. Res. 2022, 18, 319. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Kocherhans, R.; Bridgen, A.; Ackermann, M.; Tobler, K. Completion of the Porcine Epidemic Diarrhoea Coronavirus (PEDV) Genome Sequence. Virus Genes 2001, 23, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Chen, W.; Huang, J.; Jin, L.; Zhou, Y.; Chen, J.; Zhang, N.; Wu, D.; Sun, E.; Liu, G. Generation, identification, and functional analysis of monoclonal antibodies against porcine epidemic diarrhea virus nucleocapsid. Appl. Microbiol. Biotechnol. 2019, 103, 3705–3714. [Google Scholar] [CrossRef]
- Shan, Y.; Gao, Q.; Mao, J.; Zheng, J.; Xu, X.; Zhang, C.; Huang, X.; Xu, J.; Shi, F.; Yue, M.; et al. Establishment of enzyme-linked immunosorbent assays based on recombinant S1 and its truncated proteins for detection of PEDV IgA antibody. BMC Vet. Res. 2022, 18, 154. [Google Scholar] [CrossRef]
- Chang, C.-Y.; Peng, J.-Y.; Cheng, Y.-H.; Chang, Y.-C.; Wu, Y.-T.; Tsai, P.-S.; Chiou, H.-Y.; Jeng, C.-R.; Chang, H.-W. Development and comparison of enzyme-linked immunosorbent assays based on recombinant trimeric full-length and truncated spike proteins for detecting antibodies against porcine epidemic diarrhea virus. BMC Vet. Res. 2019, 15, 421. [Google Scholar] [CrossRef] [PubMed]
- Gimenez-Lirola, L.G.; Zhang, J.; Carrillo-Avila, J.A.; Chen, Q.; Magtoto, R.; Poonsuk, K.; Baum, D.H.; Piñeyro, P.; Zimmerman, J. Reactivity of Porcine Epidemic Diarrhea Virus Structural Proteins to Antibodies against Porcine Enteric Coronaviruses: Diagnostic Implications. J. Clin. Microbiol. 2017, 55, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Knuchel, M.; Ackermann, M.; Müller, H.K.; Kihm, U. An ELISA for detection of antibodies against porcine epidemic diarrhoea virus (PEDV) based on the specific solubility of the viral surface glycoprotein. Vet. Microbiol. 1992, 32, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.X.; Lien, N.T.K.; Thu, H.T.; Duy, N.D.; Duong, B.T.T.; Quyen, D.V. Changes in the spike and nucleocapsid protein of porcine epidemic diarrhea virus strain in Vietnam-a molecular potential for the vaccine development? PeerJ 2021, 9, e12329. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.T.; Nguyen, G.T.; Pham, N.B.; Le, V.P.; Trinh, T.B.N.; Vu, T.H.; Phan, H.T.; Conrad, U.; Chu, H.H. Plant-Derived Trimeric CO-26K-Equivalent Epitope Induced Neutralizing Antibodies against Porcine Epidemic Diarrhea Virus. Front. Immunol. 2020, 11, 02152. [Google Scholar] [CrossRef]
- Song, D.; Park, B. Porcine epidemic diarrhoea virus: A comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar] [CrossRef]
- Duong, B.T.T.; Thao, P.T.P.; Hoa, N.T.; Thu, H.T.; Phuoc, M.H.; Le, T.H.; Van Quyen, D. Molecular analysis reveals a distinct subgenogroup of porcine epidemic diarrhea virus in northern Vietnam in 2018–2019. Arch. Virol. 2022, 167, 2337–2346. [Google Scholar] [CrossRef]
- Sun, Y.-G.; Li, R.; Xie, S.; Qiao, S.; Li, Q.; Chen, X.-X.; Deng, R.; Zhang, G. Identification of a novel linear B-cell epitope within the collagenase equivalent domain of porcine epidemic diarrhea virus spike glycoprotein. Virus Res. 2019, 266, 34–42. [Google Scholar] [CrossRef]
- Huy, N.-X.; Kim, S.-H.; Yang, M.-S.; Kim, T.-G. Immunogenicity of a neutralizing epitope from porcine epidemic diarrhea virus: M cell targeting ligand fusion protein expressed in transgenic rice calli. Plant Cell Rep. 2012, 31, 1933–1942. [Google Scholar] [CrossRef]
- Chang, S.-H.; Bae, J.-L.; Kang, T.-J.; Kim, J.; Chung, G.-H.; Lim, C.-W.; Laude, H.; Yang, M.-S.; Jang, Y.-S. Identification of the epitope region capable of inducing neutralizing antibodies against the porcine epidemic diarrhea virus. Mol. Cell. 2002, 14, 295–299. [Google Scholar]
- Yeo, S.-G.; Hernandez, M.; Krell, P.J.; Nagy, E.E. Cloning and Sequence Analysis of the Spike Gene of Porcine Epidemic Diarrhea Virus Chinju99. Virus Genes 2003, 26, 239–246. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-H.; Hong, C.-C.; Seak, J. An ELISA for antibodies against infectious bronchitis virus using an S1 spike polypeptide. Vet. Microbiol. 2002, 85, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; He, H.; Duan, M. Development and evaluation of a recombinant CP23 antigen-based ELISA for serodiagnosis of Cryptosporidium parvum. Exp. Parasitol. 2009, 121, 157–162. [Google Scholar] [CrossRef]
- Greiner, M.; Sohr, D.; Göbel, P. A modified ROC analysis for the selection of cut-off values and the definition of intermediate results of serodiagnostic tests. J. Immunol. Method. 1995, 185, 123–132. [Google Scholar] [CrossRef]
- Oh, J.S.; Song, D.S.; Yang, J.S.; Song, J.Y.; Moon, H.J.; Kim, T.Y.; Park, B.K. Comparison of an enzyme-linked immunosorbent assay with serum neutralization test for serodiagnosis of porcine epidemic diarrhea virus infection. J. Vet. Sci. 2005, 6, 349–352. [Google Scholar] [CrossRef]
- Sondik, E.J. Clinical evaluation of test strategies. A decision analysis of parameter estimation. Clin. Lab. Med. 1982, 2, 821–833. [Google Scholar] [CrossRef] [PubMed]
- DeSilva, B.; Smith, W.; Weiner, R.; Kelley, M.; Smolec, J.; Lee, B.; Khan, M.; Tacey, R.; Hill, H.; Celniker, A. Recommendations for the Bioanalytical Method Validation of Ligand-Binding Assays to Support Pharmacokinetic Assessments of Macromolecules. Pharm. Res. 2003, 20, 1885–1900. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, B.; Liu, G. Rapid and efficient detection methods of pathogenic swine enteric coronaviruses. Appl. Microbiol. Biotechnol. 2020, 104, 6091–6100. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ma, Z.; Li, Y.; Gao, S.; Xiao, S. Porcine epidemic diarrhea virus: Molecular mechanisms of attenuation and vaccines. Microb. Pathog. 2020, 149, 104553. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.F.; Lelli, D.; Zhang, J.; Strandbygaard, B.; Moreno, A.; Lavazza, A.; Perulli, S.; Bøtner, A.; Comtet, L.; Roche, M.; et al. Diagnostic evaluation of assays for detection of antibodies against porcine epidemic diarrhea virus (PEDV) in pigs exposed to different PEDV strains. Prev. Vet. Med. 2016, 135, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.-H.; Li, Y.-Q.; Pan, Y.-Q.; Guo, Y.-Y.; Guo, F.; Shi, R.-Z.; Xing, L. Additional file 1 of The spike glycoprotein genes of porcine epidemic diarrhea viruses isolated in China. Vet. Res. 2021, 52, 87. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei, M.S.; Ahmed, M. Enzyme-Linked Immunosorbent Assay (ELISA). Method. Mol. Biol. 2022, 2508, 115–134. [Google Scholar] [PubMed]
- Bae, J.-L.; Lee, J.-G.; Kang, T.-J.; Jang, H.-S.; Jang, Y.-S.; Yang, M.-S. Induction of antigen-specific systemic and mucosal immune responses by feeding animals transgenic plants expressing the antigen. Vaccine 2003, 21, 4052–4058. [Google Scholar] [CrossRef]
- Huy, N.-X.; Kim, Y.-S.; Jun, S.-C.; Jin, Z.; Park, S.-M.; Yang, M.-S.; Kim, T.-G. Production of a heat-labile enterotoxin B subunit-porcine epidemic diarrhea virus-neutralizing epitope fusion protein in transgenic lettuce (Lactuca sativa). Biotechnol. Bioprocess Eng. 2009, 14, 731–737. [Google Scholar] [CrossRef]
- Fan, J.-H.; Zuo, Y.-Z.; Shen, X.-Q.; Gu, W.-Y.; Di, J.-M. Development of an enzyme-linked immunosorbent assay for the monitoring and surveillance of antibodies to porcine epidemic diarrhea virus based on a recombinant membrane protein. J. Virol. Method. 2015, 225, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, F.; Fan, B.; Muhammad, H.M.; Zou, Y.; Jiang, P. Development of an indirect ELISA based on a truncated S protein of the porcine epidemic diarrhea virus. Can. J. Microbiol. 2015, 61, 811–817. [Google Scholar] [CrossRef]
- Okda, F.; Liu, X.; Singrey, A.; Clement, T.; Nelson, J.; Christopher-Hennings, J.; Nelson, E.A.; Lawson, S. Development of an indirect ELISA, blocking ELISA, fluorescent microsphere immunoassay and fluorescent focus neutralization assay for serologic evaluation of exposure to North American strains of Porcine Epidemic Diarrhea Virus. BMC Vet. Res. 2015, 11, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myint, O.; Yoshida, A.; Sekiguchi, S.; Van Diep, N.; Fuke, N.; Izzati, U.Z.; Hirai, T.; Yamaguchi, R. Development of indi-rect enzyme-linked immunosorbent assay for detection of porcine epidemic diarrhea virus specific antibodies (IgG) in serum of naturally infected pigs. BMC Vet. Res. 2019, 15, 409. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Luo, Y.; Liang, X.; Wang, X. A Review of Bioactive Compounds against Porcine Enteric Coronaviruses. Viruses 2022, 14, 2217. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, Y.; Wang, X.; Zhou, J.; Ma, L.; Li, J.; Yang, L.; Ouyang, H.; Yuan, H.; Pang, D. Transmissible Gastroenteritis Virus: An Update Review and Perspective. Viruses 2023, 15, 359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Luo, S.; Gu, J.; Li, Z.; Li, K.; Yuan, W.; Ye, Y.; Li, H.; Ding, Z.; Song, D.; et al. Prevalence and phylogenetic analysis of porcine diarrhea associated viruses in southern China from 2012 to 2018. BMC Vet. Res. 2019, 15, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Query: SC1402 | S Protein | Domain COE |
|---|---|---|
| GI (n = 7) | 98.06 ± 1.96 | 98.16 ± 2.33 |
| GⅡ (n = 45) | 92.67 ± 0.89 | 96.39 ± 0.71 |
| Mutations | Mutation Rate (%) | Mutations | Mutation Rate (%) |
|---|---|---|---|
| 500L → M/P | 3.85 | 589S → N | 1.92 |
| 516A → S/T | 75.00 | 590L → P | 3.84 |
| 519G → D | 1.92 | 593G → S | 90.38 |
| 520H → L/S/Y/P/R | 21.15 | 600F → L | 1.92 |
| 521S → I | 1.92 | 604E → A/D | 7.69 |
| 522G → S | 1.92 | 607S → G | 5.77 |
| 525L → I | 9.62 | 608G → S | 1.92 |
| 526I → V | 3.84 | 611F → L | 5.77 |
| 527A → V | 1.92 | 612T → V | 1.92 |
| 535F → L | 19.23 | 627T → M | 1.92 |
| 547I → T | 1.92 | 629K → T | 5.77 |
| 548T → S | 86.54 | 631L → F | 1.92 |
| 562K → N/T | 9.62 | 632Q → E/R/G/V | 94.23 |
| 563S → K | 1.92 | 634V → I | 1.92 |
| 565D → H | 7.69 | 635T → I | 1.92 |
| Virus Anti-Serum | M ± SD | Test Value (S/P) | Result |
|---|---|---|---|
| PEDV | 1.279 ± 0.025 | 1 | + |
| PRRSV | 0.185 ± 0.014 | 0.016 | − |
| PRV | 0.094 ± 0.010 | −0.066 | − |
| CSFV | 0.174 ± 0.008 | 0.006 | − |
| PDCoV | 0.183 ± 0.011 | 0.014 | − |
| Negative serum | 0.167 ± 0.016 | 0 | − |
| Sample No. | Intra-Assay | Inter-Assay | ||
|---|---|---|---|---|
| M ± SD | CV (%) | M ± SD | CV (%) | |
| 1 | 0.089 ± 0.003 | 3.26 | 0.094 ± 0.008 | 6.59 |
| 2 | 0.232 ± 0.014 | 5.92 | 0.213 ± 0.015 | 6.85 |
| 3 | 0.474 ± 0.020 | 4.24 | 0.487 ± 0.034 | 7.01 |
| 4 | 0.773 ± 0.038 | 4.96 | 0.733 ± 0.013 | 1.74 |
| 5 | 1.317 ± 0.030 | 2.30 | 1.342 ± 0.038 | 2.87 |
| 6 | 1.792 ± 0.028 | 1.58 | 1.767 ± 0.015 | 0.86 |
| 7 | 2.026 ± 0.047 | 2.30 | 2.076 ± 0.069 | 3.33 |
| 8 | 2.774 ± 0.105 | 3.78 | 2.641 ± 0.060 | 2.27 |
| Samples | Number | COE-iELISA | |
|---|---|---|---|
| + | − | ||
| Vaccinated serum | 130 (+) | 130 | 0 |
| 34 (−) | 1 | 33 | |
| Samples | COE-iELISA | Number | Commercial ELISA Kit | Agreement (%) | Kappa Value | |
|---|---|---|---|---|---|---|
| + | − | |||||
| Clinical sera | + | 171 (A) | 168 (B) | 3 | 95.08% | 0.88 |
| − | 73 (C) | 9 | 64 (D) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Li, L.; Su, X.; Li, J.; Liao, J.; Yang, J.; Xiao, Z.; Wen, P.; Wang, H. Development of an Indirect Enzyme-Linked Immunosorbent Assay Based on the Yeast-Expressed CO-26K-Equivalent Epitope-Containing Antigen for Detection of Serum Antibodies against Porcine Epidemic Diarrhea Virus. Viruses 2023, 15, 882. https://doi.org/10.3390/v15040882
Yang X, Li L, Su X, Li J, Liao J, Yang J, Xiao Z, Wen P, Wang H. Development of an Indirect Enzyme-Linked Immunosorbent Assay Based on the Yeast-Expressed CO-26K-Equivalent Epitope-Containing Antigen for Detection of Serum Antibodies against Porcine Epidemic Diarrhea Virus. Viruses. 2023; 15(4):882. https://doi.org/10.3390/v15040882
Chicago/Turabian StyleYang, Xuqiong, Liping Li, Xiaona Su, Jiadong Li, Jiaming Liao, Jinyi Yang, Zhili Xiao, Peng Wen, and Hong Wang. 2023. "Development of an Indirect Enzyme-Linked Immunosorbent Assay Based on the Yeast-Expressed CO-26K-Equivalent Epitope-Containing Antigen for Detection of Serum Antibodies against Porcine Epidemic Diarrhea Virus" Viruses 15, no. 4: 882. https://doi.org/10.3390/v15040882