
Comparison of Genotype II African Swine Fever Virus Strain SY18 Challenge Models
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Pigs
2.3. Animal Experiments
2.4. Observation of Clinical Presentations
2.5. Sample Collection
2.6. ASFV Viral Load Analysis
2.7. Statistical Analysis
3. Results
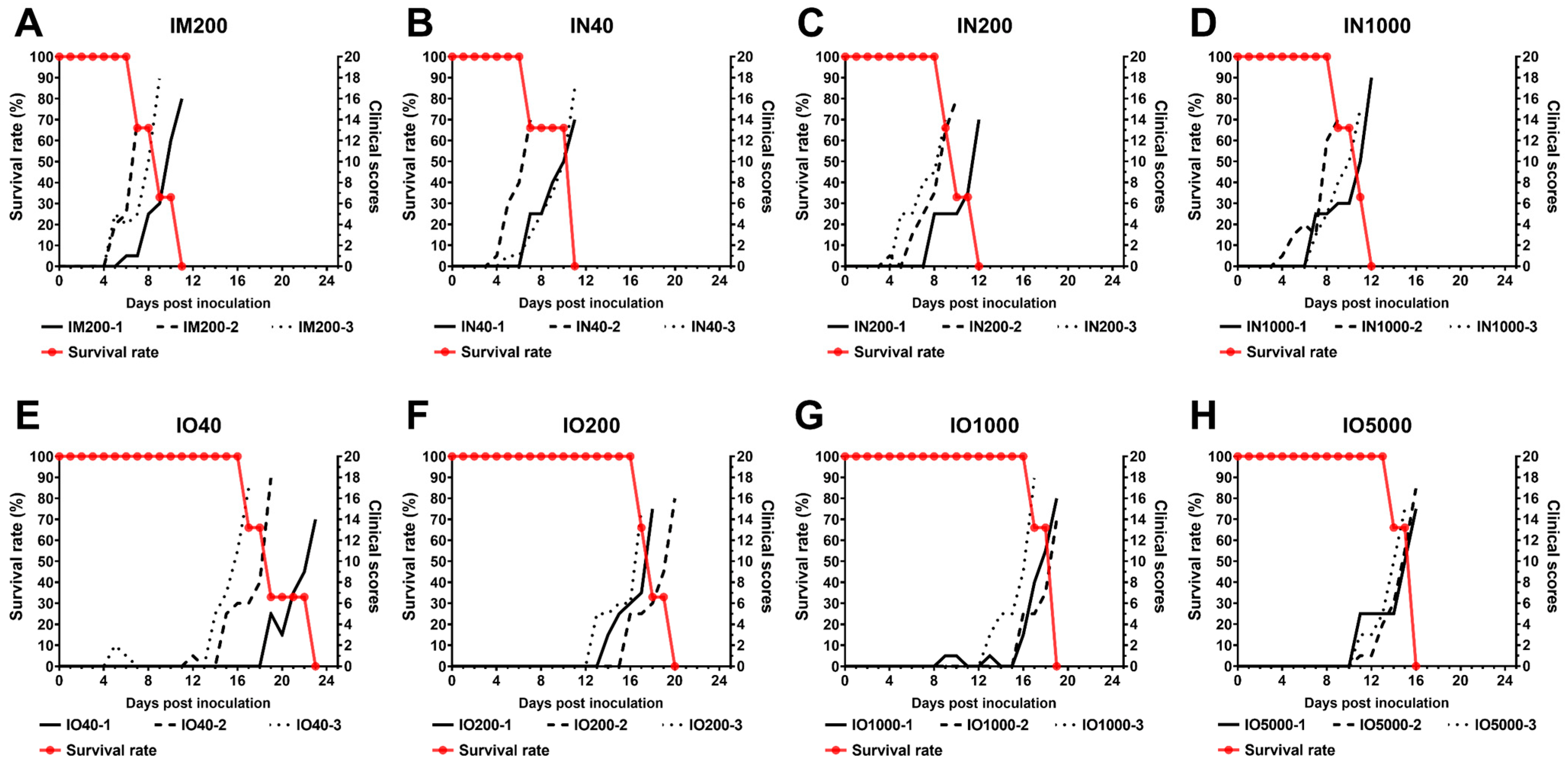
3.1. Disease Progression and Clinical Signs
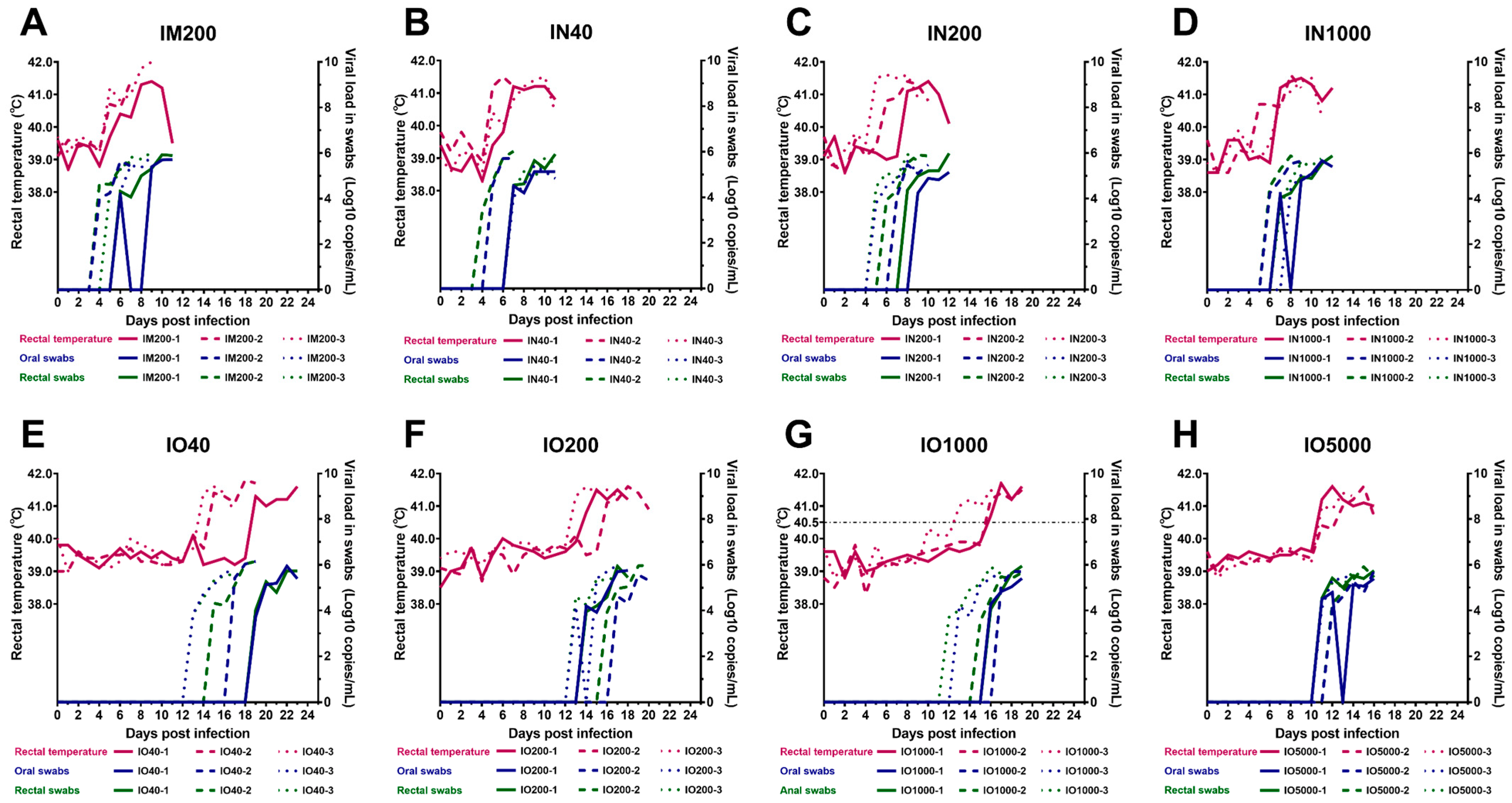
3.2. Virus Shedding via Oral and Rectal Routes
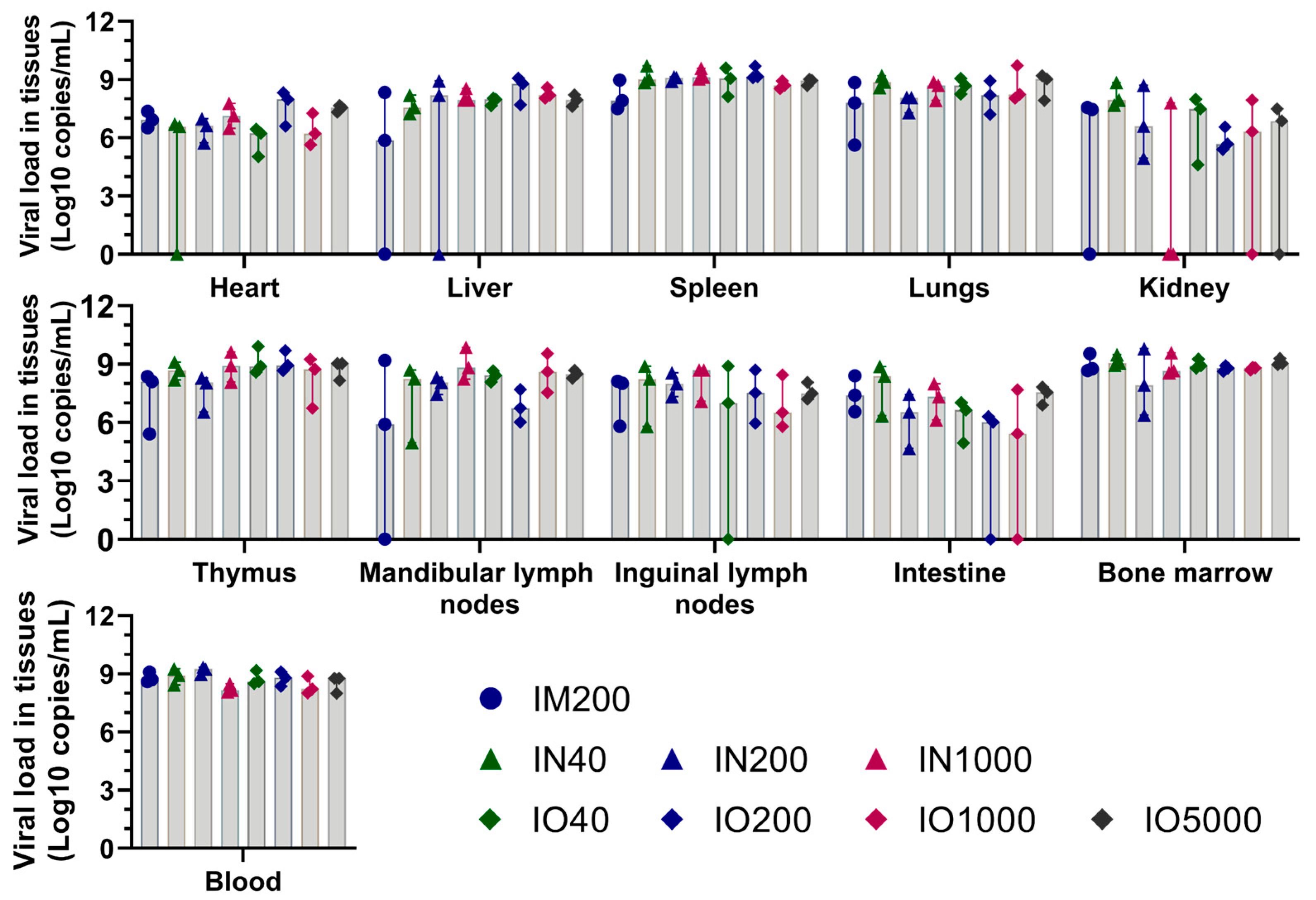
3.3. Viral Load in Tissues and Blood
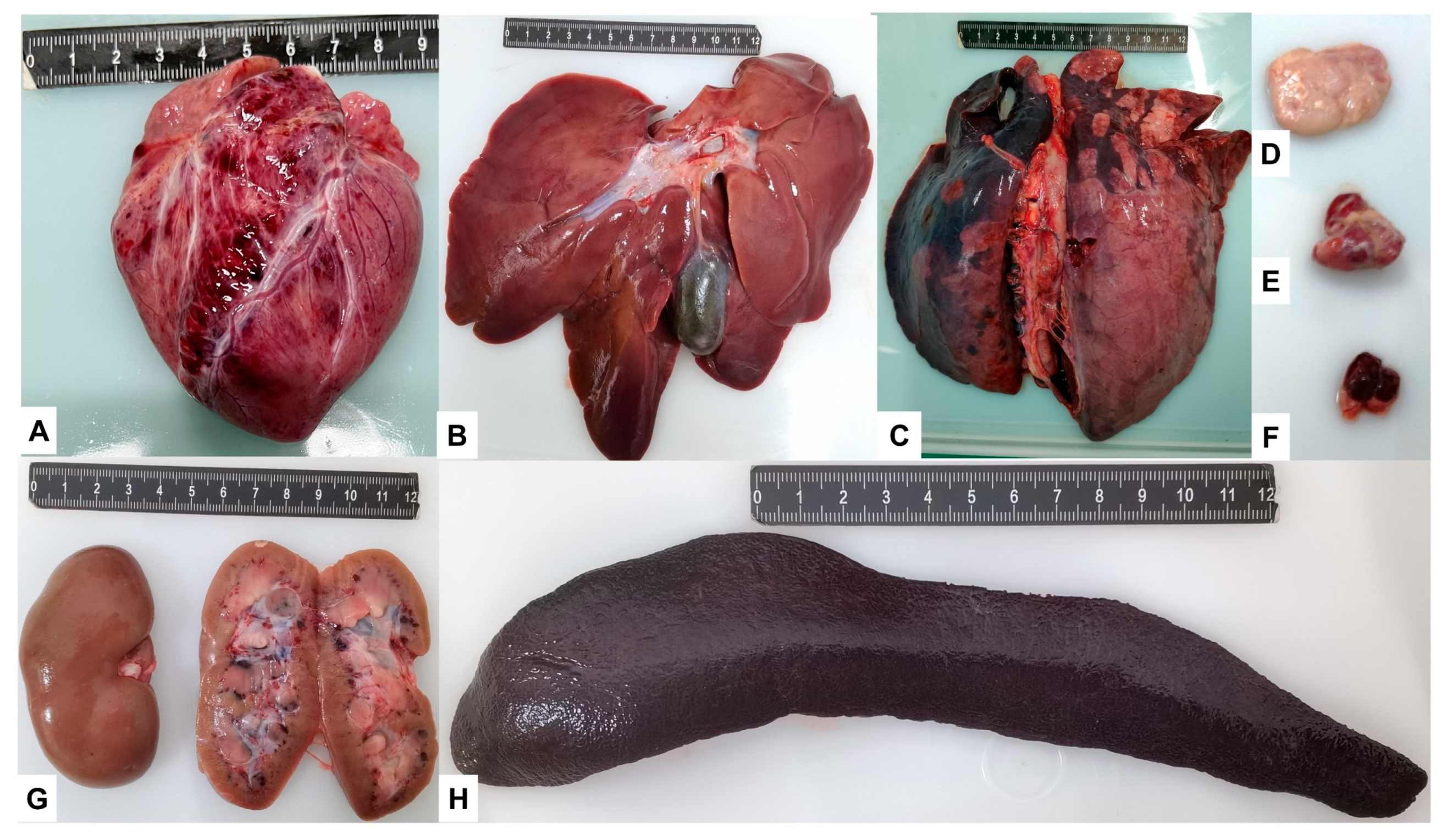
3.4. Post-Mortem Lesions
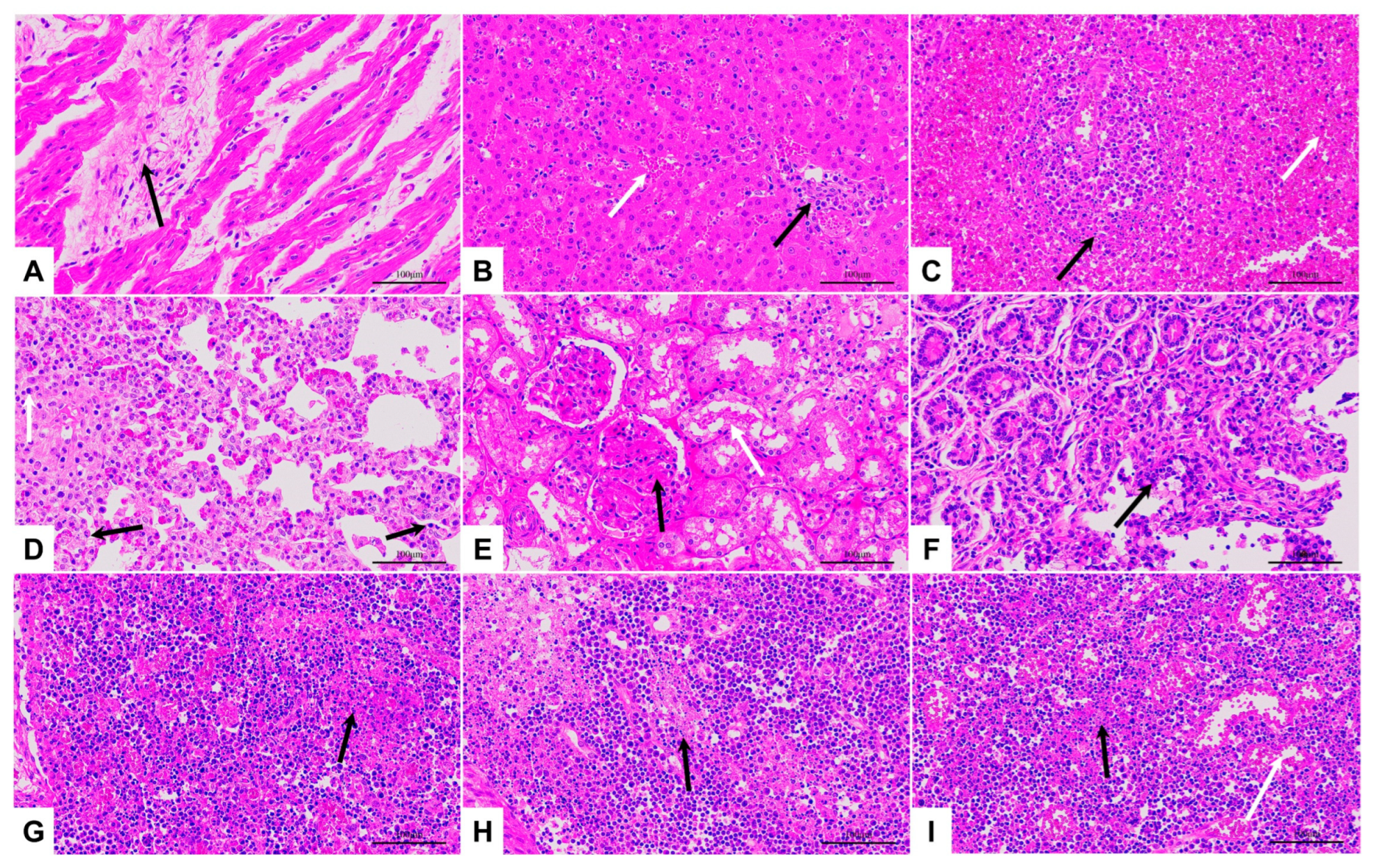
3.5. Pathological Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacolina, L.; Penrith, M.-L.; Bellini, S.; Chenais, E.; Jori, F.; Montoya, M.; Ståhl, K.; Gavier-Widén, D. (Eds.) Understanding and Combatting African Swine Fever: A European Perspective; Wageningen Academic Publishers: Wageningen, The Netherlands, 2021; ISBN 978-90-8686-357-0. [Google Scholar]
- Dixon, L.K.; Chapman, D.A.G.; Netherton, C.L.; Upton, C. African Swine Fever Virus Replication and Genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, R.J.; Michaud, V.; Heath, L.; Hutchings, G.; Oura, C.; Vosloo, W.; Dwarka, R.; Onashvili, T.; Albina, E.; Dixon, L.K. African Swine Fever Virus Isolate, Georgia, 2007. Emerg. Infect. Dis. 2008, 14, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Ankhanbaatar, U.; Sainnokhoi, T.; Khanui, B.; Ulziibat, G.; Jargalsaikhan, T.; Purevtseren, D.; Settypalli, T.B.K.; Flannery, J.; Dundon, W.G.; Basan, G.; et al. African Swine Fever Virus Genotype II in Mongolia, 2019. Transbound. Emerg. Dis. 2021, 68, 2787–2794. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.; Huang, L.; Zhang, X.; Zhang, J.; Shen, D.; Zhang, Z.; Wang, Z.; Huo, H.; Wang, W.; Huangfu, H.; et al. Genotype I African Swine Fever Viruses Emerged in Domestic Pigs in China and Caused Chronic Infection. Emerg. Microbes Infect. 2021, 10, 2183–2193. [Google Scholar] [CrossRef]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef] [Green Version]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andrés, G. A Proteomic Atlas of the African Swine Fever Virus Particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, C.; Soler, A.; Nurmoja, I.; Cano-Gómez, C.; Cvetkova, S.; Frant, M.; Woźniakowski, G.; Simón, A.; Pérez, C.; Nieto, R.; et al. Dynamics of African Swine Fever Virus (ASFV) Infection in Domestic Pigs Infected with Virulent, Moderate Virulent and Attenuated Genotype II ASFV European Isolates. Transbound. Emerg. Dis. 2021, 68, 2826–2841. [Google Scholar] [CrossRef]
- Sun, E.; Zhang, Z.; Wang, Z.; He, X.; Zhang, X.; Wang, L.; Wang, W.; Huang, L.; Xi, F.; Huangfu, H.; et al. Emergence and Prevalence of Naturally Occurring Lower Virulent African Swine Fever Viruses in Domestic Pigs in China in 2020. Sci. China Life Sci. 2021, 64, 752–765. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.G.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D.P. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef]
- Borca, M.V.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Gay, C.G.; Gladue, D.P. ASFV-G-∆I177L as an Effective Oral Nasal Vaccine against the Eurasia Strain of Africa Swine Fever. Viruses 2021, 13, 765. [Google Scholar] [CrossRef]
- Tran, X.H.; Le, T.T.P.; Nguyen, Q.H.; Do, T.T.; Nguyen, V.D.; Gay, C.G.; Borca, M.V.; Gladue, D.P. African Swine Fever Virus Vaccine Candidate ASFV-G-ΔI177L Efficiently Protects European and Native Pig Breeds against Circulating Vietnamese Field Strain. Transbound. Emerg. Dis. 2022, 69, e497–e504. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Puertas, P.; Rodríguez, F.; Oviedo, J.M.; Brun, A.; Alonso, C.; Escribano, J.M. The African Swine Fever Virus Proteins P54 and P30 Are Involved in Two Distinct Steps of Virus Attachment and Both Contribute to the Antibody-Mediated Protective Immune Response. Virology 1998, 243, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neilan, J.G.; Zsak, L.; Lu, Z.; Burrage, T.G.; Kutish, G.F.; Rock, D.L. Neutralizing Antibodies to African Swine Fever Virus Proteins P30, P54, and P72 Are Not Sufficient for Antibody-Mediated Protection. Virology 2004, 319, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Ke, J.; Zhang, J.; Yang, J.; Yue, H.; Zhou, X.; Qi, Y.; Zhu, R.; Miao, F.; Li, Q.; et al. African Swine Fever Virus Bearing an I226R Gene Deletion Elicits Robust Immunity in Pigs to African Swine Fever. J. Virol. 2021, 95, e0119921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, T.; Zhang, J.; Qi, Y.; Miao, F.; Bo, Z.; Wang, L.; Guo, X.; Zhou, X.; Yang, J.; et al. Construction and Immunoprotective Characterization of Gene Deleted African Swine Fever Virus Vaccine and Candidates. Chin. J. Vet. Sci. 2019, 39, 1421–1427. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Chen, T.; Yang, J.; Yue, H.; Wang, L.; Zhou, X.; Qi, Y.; Han, X.; Ke, J.; et al. Deletion of the L7L-L11L Genes Attenuates ASFV and Induces Protection against Homologous Challenge. Viruses 2021, 13, 255. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, J.; Zhang, Y.; Yang, J.; Wang, L.; Qi, Y.; Han, X.; Zhou, X.; Miao, F.; Chen, T.; et al. Cytokine Storm in Domestic Pigs Induced by Infection of Virulent African Swine Fever Virus. Front. Vet. Sci. 2021, 7, 601641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ke, J.; Zhang, J.; Yue, H.; Chen, T.; Li, Q.; Zhou, X.; Qi, Y.; Zhu, R.; Wang, S.; et al. I267L Is Neither the Virulence- Nor the Replication-Related Gene of African Swine Fever Virus and Its Deletant Is an Ideal Fluorescent-Tagged Virulence Strain. Viruses 2021, 14, 53. [Google Scholar] [CrossRef] [PubMed]
- King, K.; Chapman, D.; Argilaguet, J.M.; Fishbourne, E.; Hutet, E.; Cariolet, R.; Hutchings, G.; Oura, C.A.L.; Netherton, C.L.; Moffat, K.; et al. Protection of European Domestic Pigs from Virulent African Isolates of African Swine Fever Virus by Experimental Immunisation. Vaccine 2011, 29, 4593–4600. [Google Scholar] [CrossRef] [Green Version]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.S.; Drew, T.W. Development of a TaqMan® PCR Assay with Internal Amplification Control for the Detection of African Swine Fever Virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Ward, M.P. Risk Factors for the Spread of African Swine Fever in China: A Systematic Review of Chinese-language Literature. Transbound. Emerg. Dis. 2022, 69, e1289–e1298. [Google Scholar] [CrossRef]
- Mighell, E.; Ward, M.P. African Swine Fever Spread across Asia, 2018–2019. Transbound. Emerg. Dis. 2021, 68, 2722–2732. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, H.; Hou, L.; Yang, C.; Wen, Y. Advance of African Swine Fever Virus in Recent Years. Res. Vet. Sci. 2021, 136, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Bishop, R.P.; Fleischauer, C.; de Villiers, E.P.; Okoth, E.A.; Arias, M.; Gallardo, C.; Upton, C. Comparative Analysis of the Complete Genome Sequences of Kenyan African Swine Fever Virus Isolates within P72 Genotypes IX and X. Virus Genes 2015, 50, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.T.; Truong, A.D.; Dang, A.K.; Ly, D.V.; Nguyen, C.T.; Chu, N.T.; Nguyen, H.T.; Dang, H.V. Genetic Characterization of African Swine Fever Viruses Circulating in North Central Region of Vietnam. Transbound. Emerg. Dis. 2021, 68, 1697–1699. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; O’Donnell, V.; Silva, E.; Espinoza, N.; Velazquez-Salinas, L.; Moran, K.; Daite, D.A.; Barrette, R.; Faburay, B.; Holland, R.; et al. Experimental Infection of Domestic Pigs with an African Swine Fever Virus Field Strain Isolated in 2021 from the Dominican Republic. Viruses 2022, 14, 1090. [Google Scholar] [CrossRef]
- Petrovan, V.; Rathakrishnan, A.; Islam, M.; Goatley, L.C.; Moffat, K.; Sanchez-Cordon, P.J.; Reis, A.L.; Dixon, L.K. Role of African Swine Fever Virus Proteins EP153R and EP402R in Reducing Viral Persistence in Blood and Virulence in Pigs Infected with BeninΔDP148R. J. Virol. 2022, 96, e0134021. [Google Scholar] [CrossRef]
- Ramirez-Medina, E.; Vuono, E.; Silva, E.; Rai, A.; Valladares, A.; Pruitt, S.; Espinoza, N.; Velazquez-Salinas, L.; Borca, M.V.; Gladue, D.P. Evaluation of the Deletion of MGF110-5L-6L on Swine Virulence from the Pandemic Strain of African Swine Fever Virus and Use as a DIVA Marker in Vaccine Candidate ASFV-G-ΔI177L. J. Virol. 2022, 96, e00597-22. [Google Scholar] [CrossRef]
- Howey, E.B.; O’Donnell, V.; de Carvalho Ferreira, H.C.; Borca, M.V.; Arzt, J. Pathogenesis of Highly Virulent African Swine Fever Virus in Domestic Pigs Exposed via Intraoropharyngeal, Intranasopharyngeal, and Intramuscular Inoculation, and by Direct Contact with Infected Pigs. Virus Res. 2013, 178, 328–339. [Google Scholar] [CrossRef]
- Niederwerder, M.C.; Stoian, A.M.M.; Rowland, R.R.R.; Dritz, S.S.; Petrovan, V.; Constance, L.A.; Gebhardt, J.T.; Olcha, M.; Jones, C.K.; Woodworth, J.C.; et al. Infectious Dose of African Swine Fever Virus When Consumed Naturally in Liquid or Feed. Emerg. Infect. Dis. 2019, 25, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Plowright, W.; Parker, J.; Staple, R.F. The Growth of a Virulent Strain of African Swine Fever Virus in Domestic Pigs. J. Hyg. 1968, 66, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greig, A. Pathogenesis of African Swine Fever in Pigs Naturally Exposed to the Disease. J. Comp. Pathol. 1972, 82, 73–79. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Subgroups | Routes of Injection | Dose | Pigs (n) | No. |
|---|---|---|---|---|---|
| IM | IM200 | intramuscular | 200 TCID50 | 3 | IM200-1 IM200-2 IM200-3 |
| IN | IN40 | intranasal | 40 TCID50 | 3 | IN40-1 IN40-2 IN40-3 |
| IN200 | intranasal | 200 TCID50 | 3 | IN200-1 IN200-2 IN200-3 | |
| IN1000 | intranasal | 1000 TCID50 | 3 | IN1000-1 IN1000-2 IN1000-3 | |
| IO | IO40 | intraoral | 40 TCID50 | 3 | IO40-1 IO40-2 IO40-3 |
| IO200 | intraoral | 200 TCID50 | 3 | IO200-1 IO200-2 IO200-3 | |
| IO1000 | intraoral | 1000 TCID50 | 3 | IO1000-1 IO1000-2 IO1000-3 | |
| IO5000 | intraoral | 5000 TCID50 | 3 | IO5000-1 IO5000-2 IO5000-3 |
| Group | No. Infected/Total | Disease Process | Viral Load in the Shedding | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Pyrexia Days to Onset (±SD) a | Duration (±SD) | Total Days Survival (±SD) | Oral Shedding | Rectal Shedding | |||||
| Days to the Onset (±SD) | Maximum Titer (±SD) b | Days to the Onset (±SD) | Maximum Titer (±SD) b | ||||||
| IM | IM200 | 3/3 | 5.0(±1.7) | 4.0(±1.0) | 9.0(±2.0) | 5.3 (±1.2) | 5.8 (±0.1) | 5.0 (±1.0) | 5.8 (±0.2) |
| IN | Overall | 9/9 | 5.3(±1.1) d | 4.9(±0.8) d | 10.2(±1.6) d | 6.7 (±1.3) d | 5.5 (±0.2) d | 6.3 (±1.2) d | 5.9 (±0.1) d |
| IN40 | 3/3 | 5.3(±1.2) | 4.3(±1.2) | 9.7(±2.3) | 6.3 (±1.2) | 5.4 (±0.3) | 6.0 (±1.7) | 5.9 (±0.2) | |
| IN200 | 3/3 | 5.3(±1.7) | 5.0(±0.0) | 10.3(±1.5) | 7.0 (±2.0) | 5.4 (±0.2) | 6.3 (±15) | 6.0 (±0) | |
| IN1000 | 3/3 | 5.3(±1.2) | 5.3(±0.6) | 11.7(±1.5) | 7.0 (±1.0) | 5.6 (±0.2) | 6.7 (±0.6) | 5.9 (±0.1) | |
| IO | Overall | 12/12 | 13.3(±2.3) c, d | 4.6(±0.8) d | 17.9(±2.3) c, d | 14.4 (±2.7) c, d | 5.7 (±0.2) d | 13.9 (±2.4) c, d | 5.8 (±0.2) c, d |
| IO40 | 3/3 | 15.0(±2.6) c | 4.7(±0.6) | 19.7(±3.1) c | 16.3 (±3.1) c | 5.9 (±0.3) | 15.7 (±3.1) c | 5.8 (±0.3) | |
| IO200 | 3/3 | 13.3(±1.5) c | 5.0(±0.0) | 18.3(±1.5) c | 14.7 (±2.1) c | 5.8 (±0.2) | 14.3 (±1.5) c | 5.8 (±0.3) | |
| IO1000 | 3/3 | 14.0(±1.7) c | 4.3(±0.6) | 18.3(±1.2) c | 15.3 (±2.1) c | 5.6 (±0.2) | 14.3 (±2.1) c | 5.8 (±0.1) | |
| IO5000 | 3/3 | 11.0(±1.7) c | 4.3(±1.5) | 15.3(±1.2) c | 11.3 (±0.6) c | 5.6 (±0.2) | 11.3 (±0.6) c | 5.8 (±0.1) | |
| Clinical Signs and Gross Lesions | IM | IN | IO | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IM200 | Overall | IN40 | IN200 | IN1000 | Overall | IO40 | IO200 | IO1000 | IO5000 | |||
| Clinical Signs | Fever (≥40.5 °C) | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | |
| Lost appetite | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | ||
| Reluctance to stand | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | ||
| Wheezing/ coughing | 1/3 | 6/9 | 2/3 | 1/3 | 3/3 | 4/12 | 1/3 | 0/3 | 2/3 | 1/3 | ||
| Ocular discharge | 1/3 | 2/9 | 0/3 | 1/3 | 1/3 | 3/12 | 2/3 | 0/3 | 1/3 | 0/3 | ||
| Enlarged joints | 2/3 | 5/9 | 1/3 | 2/3 | 2/3 | 6/12 | 2/3 | 2/3 | 1/3 | 1/3 | ||
| Skin Purpura | 1/3 | 4/9 | 2/3 | 1/3 | 1/3 | 6/12 | 2/3 | 1/3 | 2/3 | 1/3 | ||
| Diarrhoea | 2/3 | 2/9 | 1/3 | 0/3 | 1/3 | 5/12 | 2/3 | 1/3 | 1/3 | 1/3 | ||
| Bloody diarrhoea | 0/3 | 2/9 | 1/3 | 0/3 | 1/3 | 4/12 | 1/3 | 0/3 | 0/3 | 1/3 | ||
| Gross lesions | Heart | Epicardial haemorrhage | 2/3 | 5/9 | 2/3 | 1/3 | 2/3 | 7/12 | 2/3 | 2/3 | 3/3 | 1/3 |
| Hydropericardium | 0/3 | 2/9 | 0/3 | 1/3 | 1/3 | 2/12 | 1/3 | 0/3 | 1/3 | 0/3 | ||
| Liver | Enlargement | 2/3 | 7/9 | 2/3 | 3/3 | 2/3 | 10/12 | 3/3 | 2/3 | 3/3 | 2/3 | |
| Friable | 3/3 | 8/9 | 3/3 | 3/3 | 2/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | ||
| Spleen | Thickened | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | |
| Dark | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | ||
| Enlarged | 3/3 | 7/9 | 2/3 | 3/3 | 2/3 | 9/12 | 3/3 | 2/3 | 2/3 | 2/3 | ||
| Friable | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | ||
| Lungs | Interstitial pneumonia | 2/3 | 7/9 | 2/3 | 2/3 | 3/3 | 9/12 | 2/3 | 2/3 | 2/3 | 3/3 | |
| Interlobular edema | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 11/12 | 3/3 | 3/3 | 2/3 | 3/3 | ||
| Kidney | Petechiae | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | |
| Thymus | Haemorrhage | 3/3 | 9/9 | 3/3 | 3/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | |
| Submaxillary lymph nodes | Enlargement | 2/3 | 8/9 | 3/3 | 2/3 | 3/3 | 12/12 | 3/3 | 3/3 | 3/3 | 3/3 | |
| Haemorrhage | 3/3 | 7/9 | 2/3 | 3/3 | 2/3 | 11/12 | 3/3 | 3/3 | 2/3 | 3/3 | ||
| Inguinal lymph nodes | Enlargement | 3/3 | 8/9 | 3/3 | 3/3 | 2/3 | 10/12 | 2/3 | 3/3 | 2/3 | 3/3 | |
| Haemorrhage | 1/3 | 4/9 | 1/3 | 2/3 | 1/3 | 6/12 | 1/3 | 2/3 | 2/3 | 1/3 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, X.; Fan, J.; Guo, X.; Chen, T.; Yang, J.; Zhang, Y.; Mi, L.; Zhang, F.; Miao, F.; Li, M.; et al. Comparison of Genotype II African Swine Fever Virus Strain SY18 Challenge Models. Viruses 2023, 15, 858. https://doi.org/10.3390/v15040858
Zhou X, Fan J, Guo X, Chen T, Yang J, Zhang Y, Mi L, Zhang F, Miao F, Li M, et al. Comparison of Genotype II African Swine Fever Virus Strain SY18 Challenge Models. Viruses. 2023; 15(4):858. https://doi.org/10.3390/v15040858
Chicago/Turabian StyleZhou, Xintao, Jiaqi Fan, Xiaopan Guo, Teng Chen, Jinjin Yang, Yanyan Zhang, Lijuan Mi, Fei Zhang, Faming Miao, Min Li, and et al. 2023. "Comparison of Genotype II African Swine Fever Virus Strain SY18 Challenge Models" Viruses 15, no. 4: 858. https://doi.org/10.3390/v15040858