
Pathology of African Swine Fever in Reproductive Organs of Mature Breeding Boars
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Cells and Virus
2.3. Animal Experiment
2.4. Sample Collection
2.5. Pathomorphological Analysis
2.5.1. Gross Pathology and Macroscopic Scoring
2.5.2. Histopathology and Immunohistochemistry
2.5.3. Semiquantitative Scoring
2.6. ASFV Genome Detection and Virus Isolation
2.7. Semen Processing and Investigation
3. Results
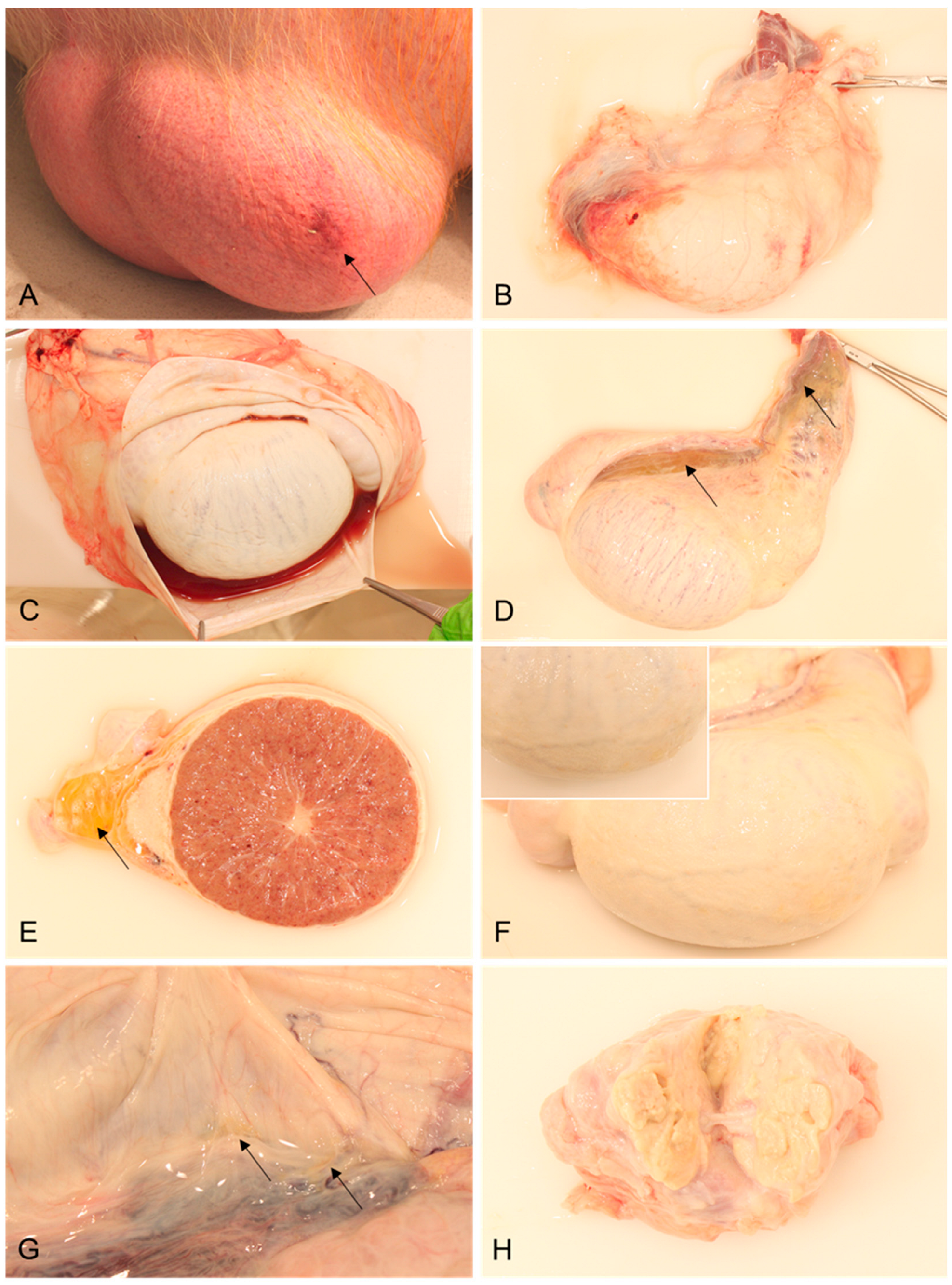
3.1. Gross Pathology
Pathology of Male Reproductive Organs
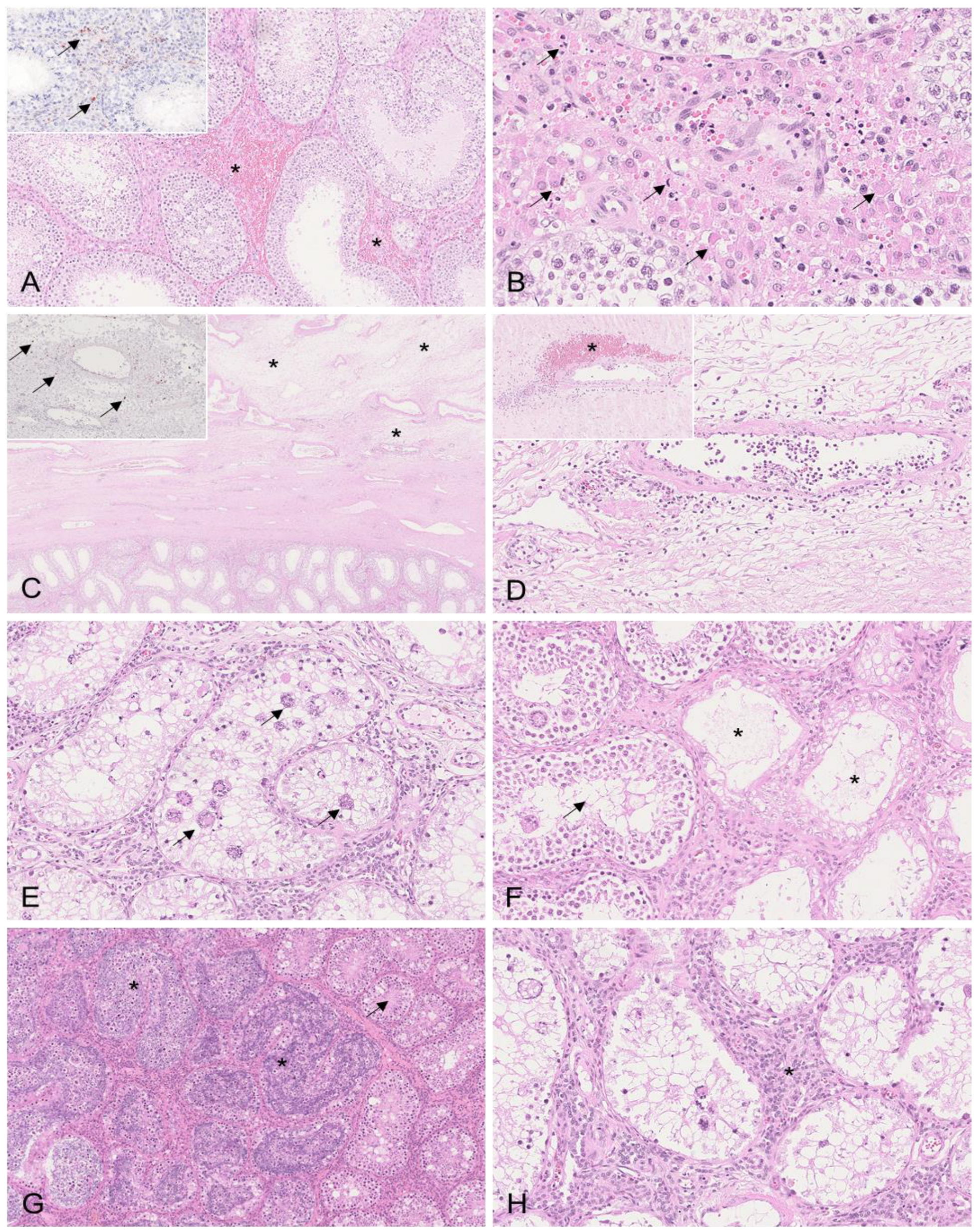
3.2. Histopathology and Immunohistochemistry
3.2.1. Testis
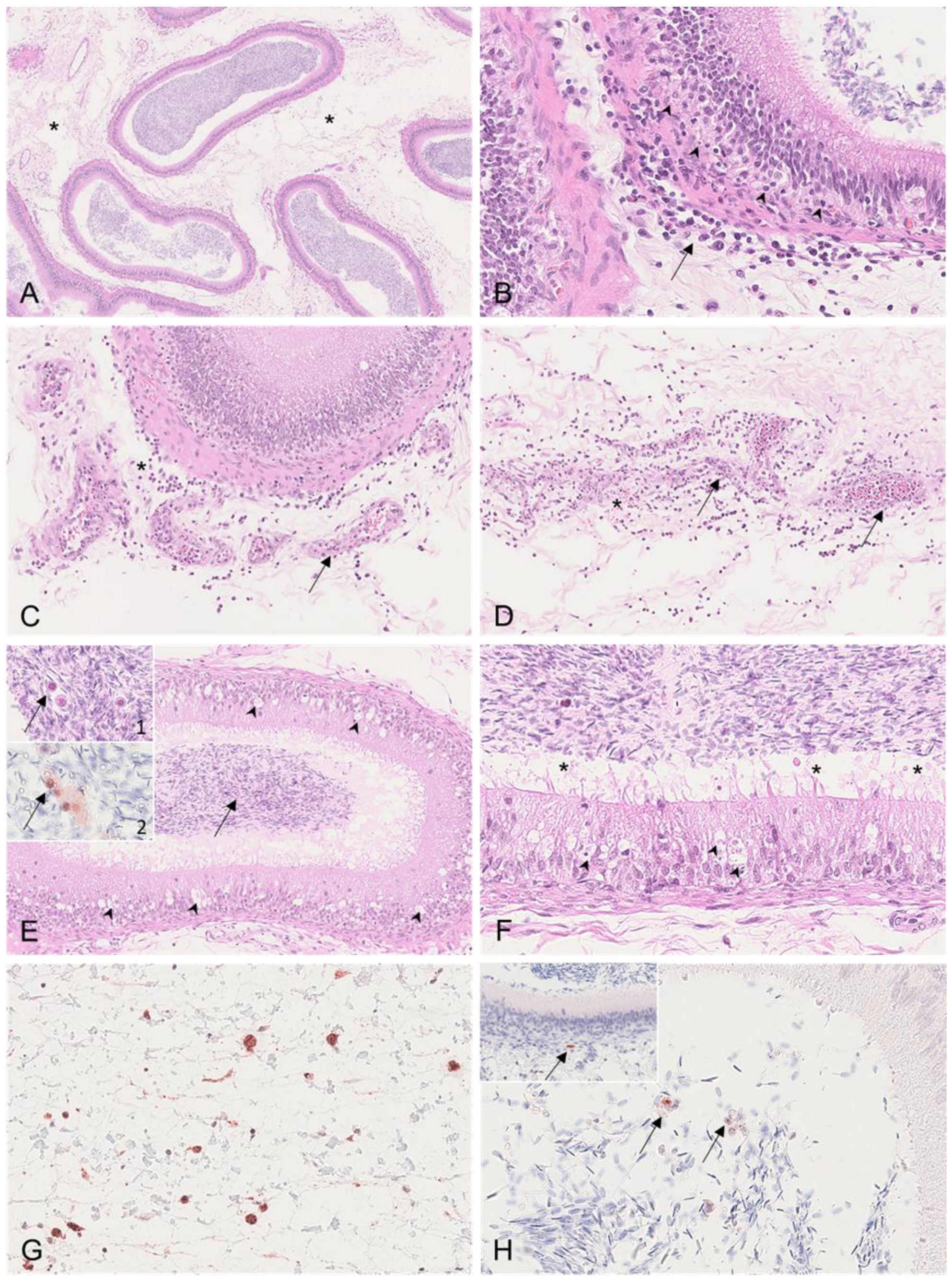
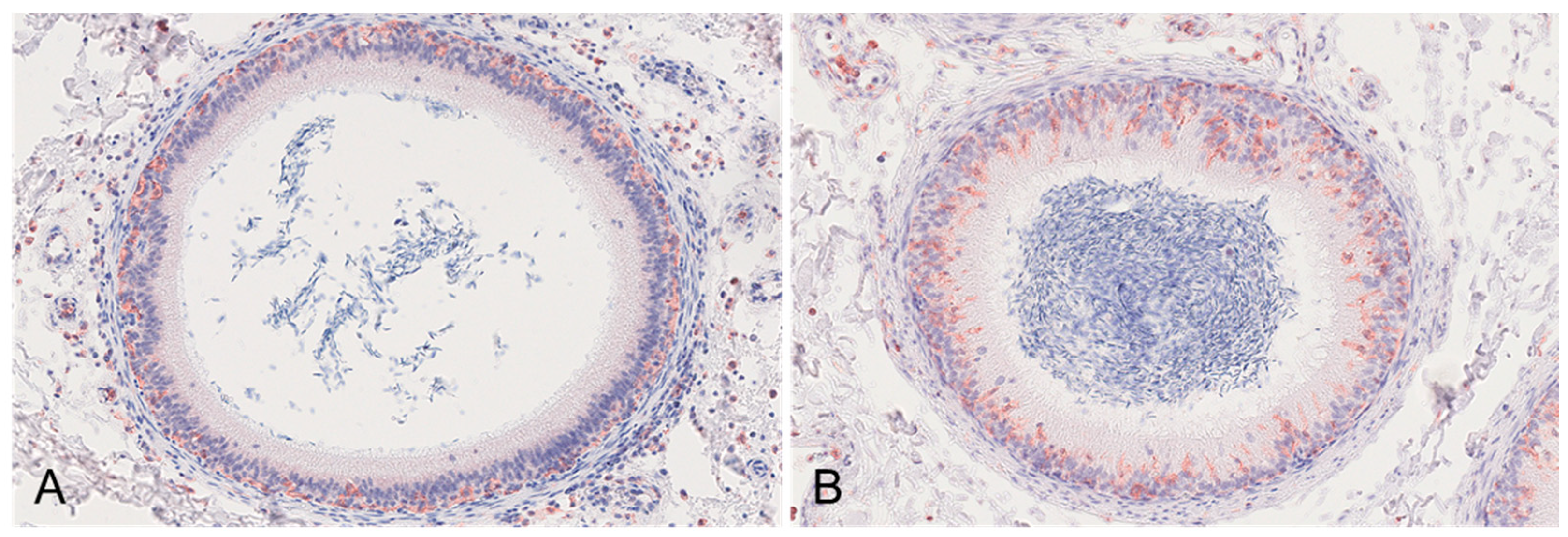
3.2.2. Epididymis
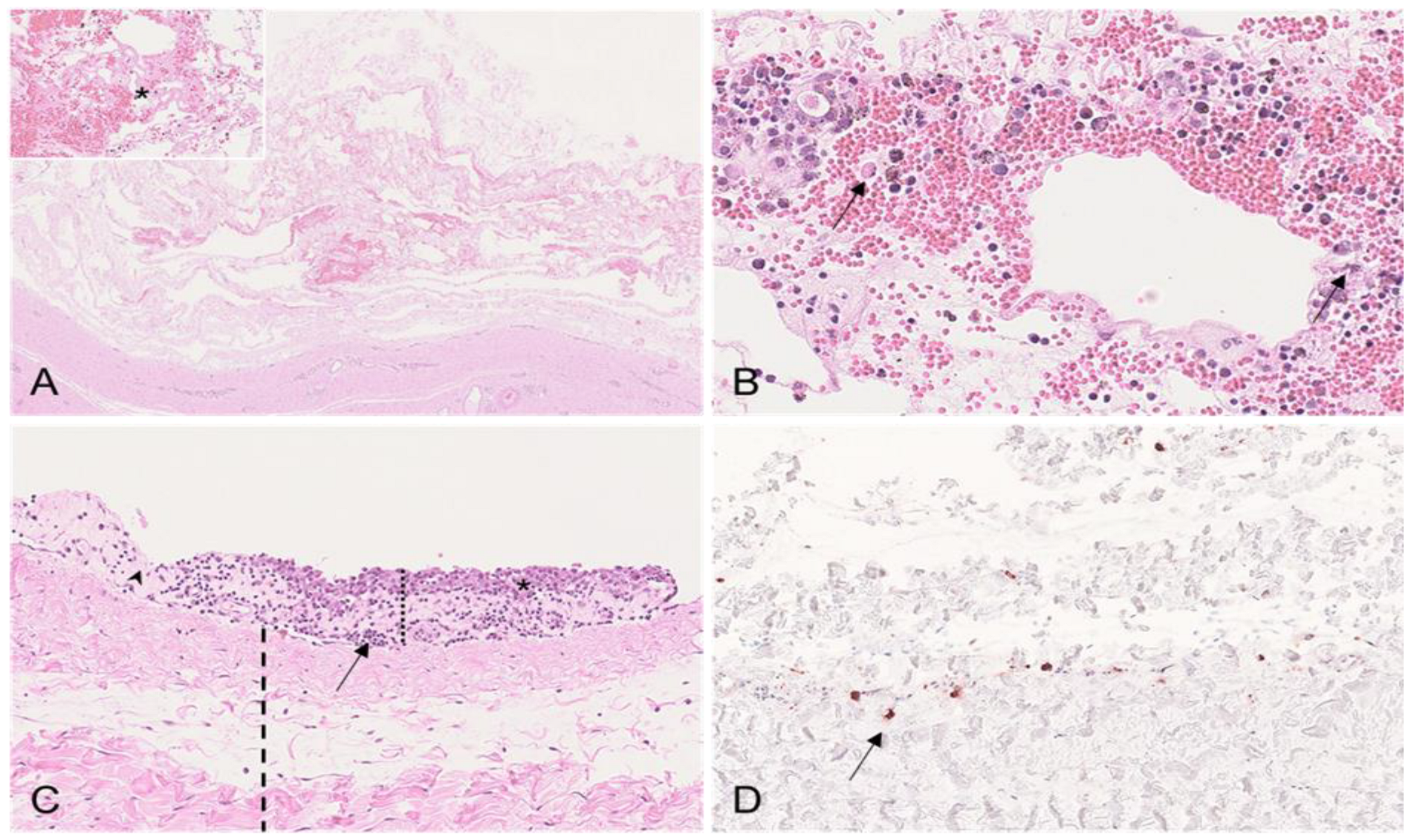
3.2.3. Tunicae Testis
3.2.4. Vesicular Gland
3.2.5. Prostate
3.2.6. Bulbourethral Gland
3.3. Detection of ASFV Genome and Infectious Virus
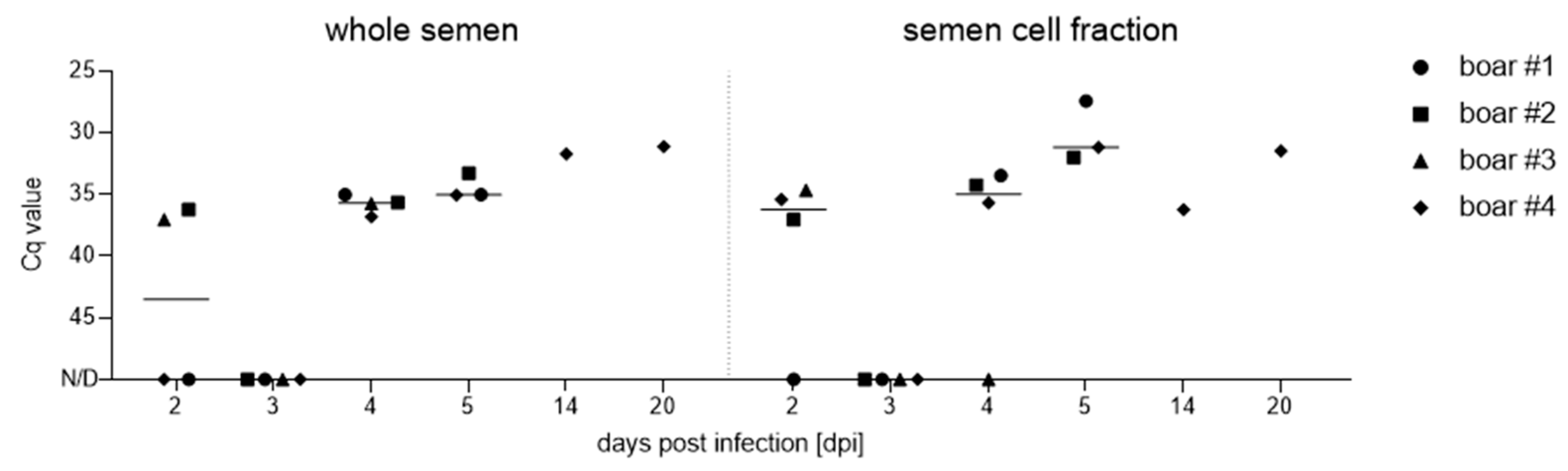
3.3.1. Semen and Semen Cell Fraction
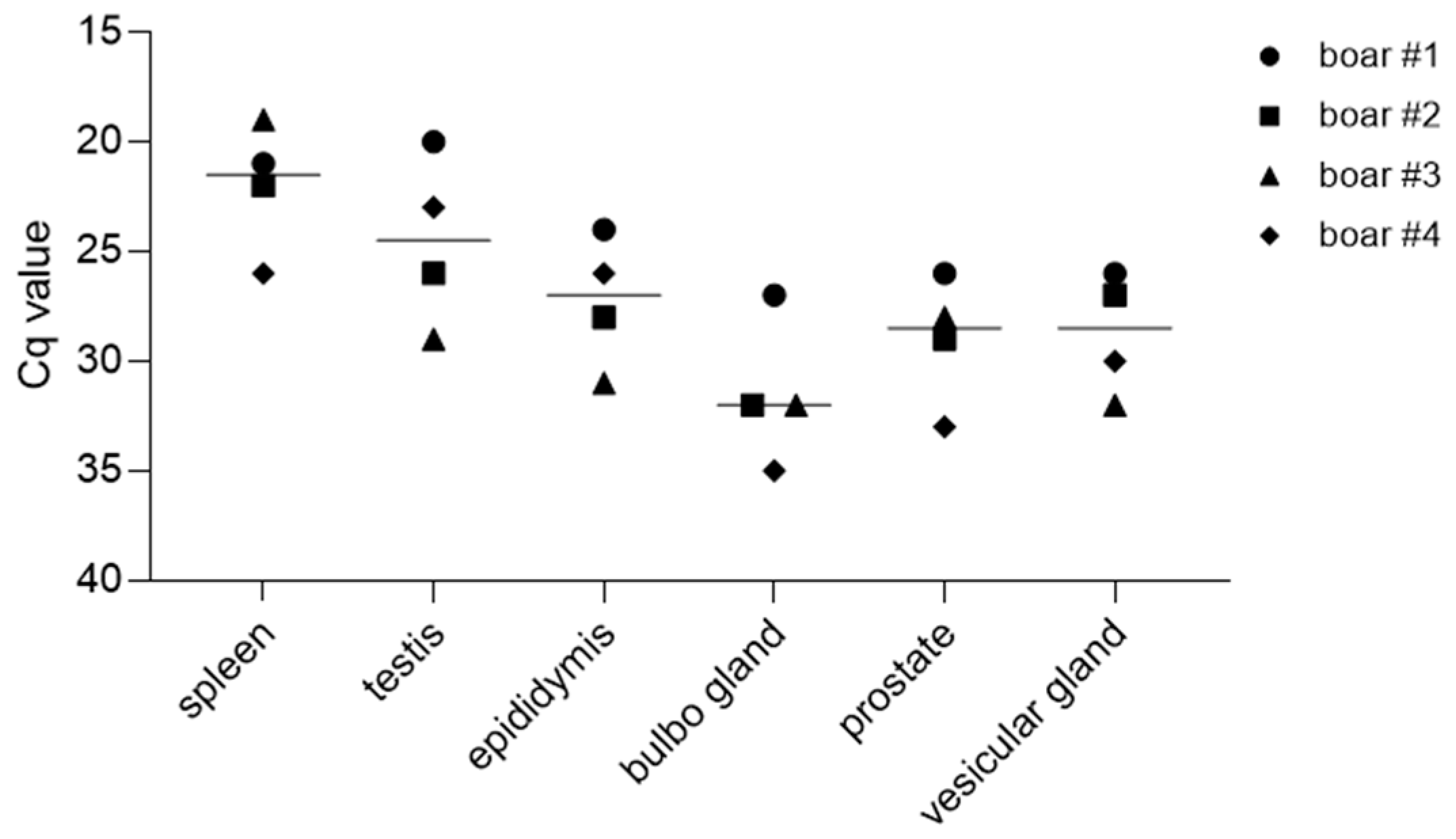
3.3.2. Reproductive Organs
3.4. Assessment of Sperm Motility and Viability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef]
- Brake, D.A. African Swine Fever Modified Live Vaccine Candidates: Transitioning from Discovery to Product Development through Harmonized Standards and Guidelines. Viruses 2022, 14, 2619. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Bui, V.N.; Dao, D.T.; Bui, N.A.; Le, T.D.; Kieu, M.A.; Nguyen, Q.H.; Tran, L.H.; Roh, J.-H.; So, K.-M.; et al. Pathogenicity of an African swine fever virus strain isolated in Vietnam and alternative diagnostic specimens for early detection of viral infection. Porc. Health Manag. 2021, 7, 36. [Google Scholar] [CrossRef]
- Walczak, M.; Żmudzki, J.; Mazur-Panasiuk, N.; Juszkiewicz, M.; Woźniakowski, G. Analysis of the Clinical Course of Experimental Infection with Highly Pathogenic African Swine Fever Strain, Isolated from an Outbreak in Poland. Aspects Related to the Disease Suspicion at the Farm Level. Pathogens 2020, 9, 237. [Google Scholar] [CrossRef]
- Guinat, C.; Reis, A.L.; Netherton, C.L.; Goatley, L.; Pfeiffer, D.U.; Dixon, L. Dynamics of African swine fever virus shedding and excretion in domestic pigs infected by intramuscular inoculation and contact transmission. Vet. Res. 2014, 45, 93. [Google Scholar] [CrossRef] [PubMed]
- Thacker, B.J.; Larsen, R.E.; Joo, H.S.; Leman, A.D. Swine diseases transmissible with artificial insemination. J. Am. Vet. Med. Assoc. 1984, 185, 511–516. [Google Scholar]
- Roszyk, H.; Franzke, K.; Breithaupt, A.; Deutschmann, P.; Pikalo, J.; Carrau, T.; Blome, S.; Sehl-Ewert, J. The Role of Male Reproductive Organs in the Transmission of African Swine Fever-Implications for Transmission. Viruses 2021, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Pikalo, J.; Schoder, M.E.; Sehl, J.; Breithaupt, A.; Tignon, M.; Cay, A.B.; Gager, A.M.; Fischer, M.; Beer, M.; Blome, S. The African swine fever virus isolate Belgium 2018/1 shows high virulence in European wild boar. Transbound. Emerg. Dis. 2020, 67, 1654–1659. [Google Scholar] [CrossRef]
- Friedrichs, V.; Reicks, D.; Hasenfuss, T.; Gerstenkorn, E.; Zimmerman, J.; Nelson, E.; Carrau, T.; Deutschmann, P.; Sehl-Ewert, J.; Roszyk, H.; et al. Artificial insemination as alternative transmission route for African Swine Fever Virus. Pathogens 2022, 11, 1539. [Google Scholar] [CrossRef]
- Pietschmann, J.; Guinat, C.; Beer, M.; Pronin, V.; Tauscher, K.; Petrov, A.; Keil, G.; Blome, S. Course and transmission characteristics of oral low-dose infection of domestic pigs and European wild boar with a Caucasian African swine fever virus isolate. Arch. Virol. 2015, 160, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Cardiel, I.; Ballester, M.; Solanes, D.; Nofrarias, M.; Lopez-Soria, S.; Argilaguet, J.M.; Lacasta, A.; Accensi, F.; Rodriguez, F.; Segales, J. Standardization of pathological investigations in the framework of experimental ASFV infections. Virus Res. 2013, 173, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Sehl, J.; Pikalo, J.; Schäfer, A.; Franzke, K.; Pannhorst, K.; Elnagar, A.; Blohm, U.; Blome, S.; Breithaupt, A. Comparative Pathology of Domestic Pigs and Wild Boar Infected with the Moderately Virulent African Swine Fever Virus Strain “Estonia 2014”. Pathogens 2020, 9, 662. [Google Scholar] [CrossRef] [PubMed]
- Sehl-Ewert, J.; Deutschmann, P.; Breithaupt, A.; Blome, S. Pathology of African Swine Fever in Wild Boar Carcasses Naturally Infected with German Virus Variants. Pathogens 2022, 11, 1386. [Google Scholar] [CrossRef]
- Gomez-Villamandos, J.C.; Hervas, J.; Mendez, A.; Carrasco, L.; Martin de las Mulas, J.; Villeda, C.J.; Wilkinson, P.J.; Sierra, M.A. Experimental African swine fever: Apoptosis of lymphocytes and virus replication in other cells. J. Gen. Virol. 1995, 76 Pt 9, 2399–2405. [Google Scholar] [CrossRef]
- Lee, S.; Illei, P.B.; Han, J.S.; Epstein, J.I. Florid mesothelial hyperplasia of the tunica vaginalis mimicking malignant mesothelioma: A clinicopathologic study of 12 cases. Am. J. Surg. Pathol. 2014, 38, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Dagur, G.; Gandhi, J.; Suh, Y.; Weissbart, S.; Sheynkin, Y.R.; Smith, N.L.; Joshi, G.; Khan, S.A. Classifying Hydroceles of the Pelvis and Groin: An Overview of Etiology, Secondary Complications, Evaluation, and Management. Curr. Urol. 2017, 10, 1–14. [Google Scholar] [CrossRef]
- Pedersen, K.; Blirup-Plum, S.A.; Kristensen, C.S.; Kvisgaard, L.K.; Skade, L.; Jensen, H.E.; Larsen, L.E. Virological and Histopathological Findings in Boars Naturally Infected With Porcine Reproductive and Respiratory Syndrome Virus Type 1. Front. Microbiol. 2022, 13, 874498. [Google Scholar] [CrossRef]
- Cavalieri, J.; van Camp, S.D. Bovine seminal vesiculitis. A review and update. Vet. Clin. North Am. Food Anim. Pract. 1997, 13, 233–241. [Google Scholar] [CrossRef]
- Ohsawa, K.; Imai, Y.; Sasaki, Y.; Kohsaka, S. Microglia/macrophage-specific protein Iba1 binds to fimbrin and enhances its actin-bundling activity. J. Neurochem. 2004, 88, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, S.; Meinhardt, A. The macrophages in testis function. J. Reprod. Immunol. 2017, 119, 107–112. [Google Scholar] [CrossRef]
- Chen, G.; Zheng, B. Effect of macrophages in semen on sperm quality. Reprod. Biol. Endocrinol. 2021, 19, 38. [Google Scholar] [CrossRef]
- Wang, M.; Yang, Y.; Cansever, D.; Wang, Y.; Kantores, C.; Messiaen, S.; Moison, D.; Livera, G.; Chakarov, S.; Weinberger, T.; et al. Two populations of self-maintaining monocyte-independent macrophages exist in adult epididymis and testis. Proc. Natl. Acad. Sci. USA 2021, 118, e2013686117. [Google Scholar] [CrossRef]
- DuPre, N.C.; Flavin, R.; Sfanos, K.S.; Unger, R.H.; To, S.; Gazeeva, E.; Fiorentino, M.; De Marzo, A.M.; Rider, J.R.; Mucci, L.A. Corpora amylacea in prostatectomy tissue and associations with molecular, histological, and lifestyle factors. Prostate 2018, 78, 1172–1180. [Google Scholar] [CrossRef]
- Hassler, O. Calcifications in the prostate gland and adjacent tissues. A combined biophysical and histological study. Pathol. Microbiol. 1968, 31, 97–107. [Google Scholar] [CrossRef]
- Magura, C.E.; Spector, M. Scanning electron microscopy of human prostatic corpora amylacea and corpora calculi, and prostatic calculi. Scanning Electron Microsc. 1979, 3, 713–720. [Google Scholar]
- Sánchez-Vizcaíno, J.M.; Mur, L.; Gomez-Villamandos, J.C.; Carrasco, L. An update on the epidemiology and pathology of African swine fever. J. Comp. Pathol. 2015, 152, 9–21. [Google Scholar] [CrossRef]
- Isobe, A.; Tasaki, M.; Inoue, T.; Terao, H.; Souba, J.; Nagano, K.; Hagiwara, H. Histopathological background data of the systemic organs of CLAWN miniature swine with coronary artery stent implantation. J. Toxicol. Pathol. 2017, 30, 25–38. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Gregory, M.; Cyr, D.G. The blood-epididymis barrier and inflammation. Spermatogenesis 2014, 4, e979619. [Google Scholar] [CrossRef] [PubMed]
- Byas, A.D.; Ebel, G.D. Comparative Pathology of West Nile Virus in Humans and Non-Human Animals. Pathogens 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Li, M.W.; Xia, W.; Mruk, D.D.; Wang, C.Q.; Yan, H.H.; Siu, M.K.; Lui, W.Y.; Lee, W.M.; Cheng, C.Y. Tumor necrosis factor {alpha} reversibly disrupts the blood-testis barrier and impairs Sertoli-germ cell adhesion in the seminiferous epithelium of adult rat testes. J. Endocrinol. 2006, 190, 313–329. [Google Scholar] [CrossRef]
- Korah, N.; Smith, C.E.; D’Azzo, A.; Mui, J.; Hermo, L. Characterization of cell- and region-specific abnormalities in the epididymis of cathepsin A deficient mice. Mol. Reprod. Dev. 2003, 66, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M. Diagnosis and therapy of acute prostatitis, epididymitis and orchitis. Andrologia 2008, 40, 76–80. [Google Scholar] [CrossRef]
- Sauter-Louis, C.; Conraths, F.J.; Probst, C.; Blohm, U.; Schulz, K.; Sehl, J.; Fischer, M.; Forth, J.H.; Zani, L.; Depner, K.; et al. African Swine Fever in Wild Boar in Europe-A Review. Viruses 2021, 13, 1717. [Google Scholar] [CrossRef] [PubMed]
- Zani, L.; Dietze, K.; Dimova, Z.; Forth, J.H.; Denev, D.; Depner, K.; Alexandrov, T. African Swine Fever in a Bulgarian Backyard Farm-A Case Report. Vet. Sci. 2019, 6, 94. [Google Scholar] [CrossRef]
- Forth, J.H.; Calvelage, S.; Fischer, M.; Hellert, J.; Sehl-Ewert, J.; Roszyk, H.; Deutschmann, P.; Reichold, A.; Lange, M.; Thulke, H.-H.; et al. African swine fever virus–variants on the rise. bioRxiv 2022. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
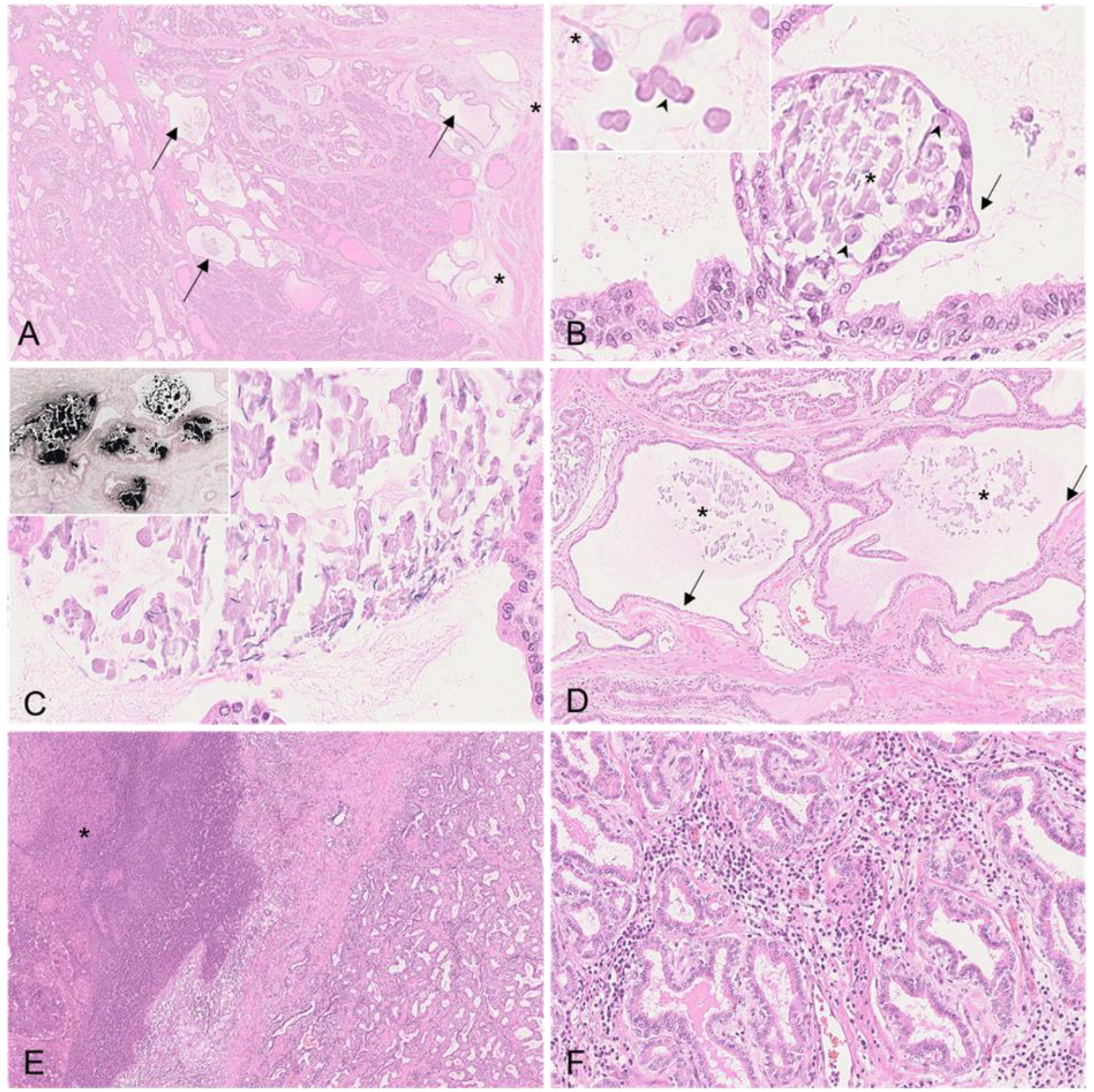{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Boar #1 | Boar #2 | Boar #3 | Boar #4 | |
|---|---|---|---|---|
| Day of humane endpoint | 10 | 11 | 17 | 25 |
| Scrotum | Scrotal skin with hemorrhages; serosanguinous hydrocele testis | - | Serous hydrocele testis | |
| Testicle | Parenchymal hemorrhages; edema of mesorchium | Mesothelial proliferations | ||
| Tunicae testis | Hemorrhage; edema | - | ||
| Prostate | - | Suppurative prostatitis | ||
| Boar #1 | Boar #2 | Boar #3 | Boar #4 | ||
|---|---|---|---|---|---|
| Day of humane end-point | 10 | 11 | 17 | 25 | |
| Testis | HE | Interstitial hemorrhages Single-cell apoptosis/necrosis of Leydig cells Edema of tunica albuginea Vasculitis/vasculopathy | Interstitial hemorrhages | Germ cell degeneration/atrophy with formation of multinucleated giant cells Germinal epithelial degeneration with vacuolation of tubular epithelium Tubular epithelial necrosis Tubular dilation Hyperplasia of Leydig cells Interstitial infiltration with neutrophils Interstitial hemorrhage Edema and congestion of tunica albuginea | Germ cell degeneration/atrophy with formation of multinucleated giant cells Germinal epithelial degeneration with vacuolation of tubular epithelium Tubular epithelial necrosis Edema and congestion of tunica albuginea |
| IHC | 3 | 1 | 0 | 0 | |
| Interstitial histiocytes Tunica albuginea | |||||
| Tunicae testis | HE | N/A | Hemorrhagic edema of fascia spermatica externa with fibrin Lymphohistioplasmacytic infiltrates in fascia spermatica externa Mesothelial proliferations | Hemorrhagic edema of fascia spermatica externa Lymphohistioplasmacytic infiltrates in fascia spermatica externa Mesothelial proliferations | Lymphohistioplasmacytic infiltrates in fascia spermatica externa |
| IHC | N/A | 2 | 0 | 0 | |
| N/A | Fascia spermatica externa, histiocytes | ||||
| Epididymis | HE | Head/body: Intertubular edema Interstitial hemorrhages Peritubular infiltration with histiocytes/lymphocytes Vasculitis/vasculopathy | Head/body: Intertubular edema Interstitial hemorrhages Perivascular infiltration of lymphocytes and histocytes | Head/body: Intertubular edema Perivascular infiltration with lymphocytes and histiocytes Vacuolation and degeneration of tubular epithelium Tail: Intertubular edema Peritubular infiltration with lymphocytes and histiocytes Tunica vaginalis: Perivascular infiltration of lymphocytes and histiocytes Fibrovascular projections | Head/body: Vacuolation and degeneration of tubular epithelium |
| IHC | 2 | 0 | 0 | 0 | |
| Interstitial histiocytes Myofibroblasts Luminal round cells | |||||
| Vesicular gland | HE | Luminal content with cellular debris and spermatozoa | Interstitial hemorrhages Luminal content with cellular debris and spermatozoa | Luminal content with cellular debris and spermatozoa | Single-cell apoptosis/necrosis of glandular epithelium Luminal content with cellular debris and spermatozoa |
| IHC | 0 | ||||
| Prostate | HE | - | - | Dilation of prostatic tubules Luminal content with cellular debris, spermatozoa, and corpora amylacea; mineralized Flattening of tubular epithelium with single-cell apoptosis/necrosis Edema of fibromuscular tissue | Suppurative prostatitis Dilation of prostatic tubules Luminal content with cellular debris Interstitial infiltration with plasma cells |
| IHC | 0 | ||||
| Bulbourethral gland | HE | - | - | - | - |
| IHC | 0 | ||||
| #1 | #2 | #3 | #4 | |
|---|---|---|---|---|
| Testis | +++ | ++ | ++ | − |
| Epididymis | ++ | ++ | ++ | − |
| Bulbourethral gland | ++ | + | + | − |
| Vesicular gland | ++ | ++ | ++ | ++ |
| Prostate | +++ | +++ | ++ | ++ |
| Spleen | +++ | +++ | − | ++ |
| dpi | Boar # | Normal | Abnormal | % Abnormalities |
|---|---|---|---|---|
| 2 | 1 | 108 | 13 | 10.7 |
| 2 | 167 | 13 | 7.2 | |
| 3 | 143 | 10 | 6.5 | |
| 4 | 100 | 7 | 6.5 | |
| 3 | 1 | 211 | 26 | 11 |
| 2 | 208 | 20 | 8.8 | |
| 3 | 129 | 15 | 10.4 | |
| 4 | 242 | 21 | 8 | |
| 4 | 1 | 113 | 16 | 12.4 |
| 2 | 247 | 28 | 10.2 | |
| 3 | 107 | 11 | 9.3 | |
| 4 | 148 | 17 | 10.3 | |
| 5 | 1 | 117 | 19 | 14 |
| 2 | 172 | 24 | 12.2 | |
| 3 | — | — | — | |
| 4 | 180 | 21 | 10.4 | |
| 14 | 4 | 139 | 29 | 17.3 |
| 20 | 4 | 247 | 51 | 17.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehl-Ewert, J.; Friedrichs, V.; Carrau, T.; Deutschmann, P.; Blome, S. Pathology of African Swine Fever in Reproductive Organs of Mature Breeding Boars. Viruses 2023, 15, 729. https://doi.org/10.3390/v15030729
Sehl-Ewert J, Friedrichs V, Carrau T, Deutschmann P, Blome S. Pathology of African Swine Fever in Reproductive Organs of Mature Breeding Boars. Viruses. 2023; 15(3):729. https://doi.org/10.3390/v15030729
Chicago/Turabian StyleSehl-Ewert, Julia, Virginia Friedrichs, Tessa Carrau, Paul Deutschmann, and Sandra Blome. 2023. "Pathology of African Swine Fever in Reproductive Organs of Mature Breeding Boars" Viruses 15, no. 3: 729. https://doi.org/10.3390/v15030729