
Key Factors and Parameter Ranges for Immune Control of Equine Infectious Anemia Virus Infection

Abstract
:1. Introduction
2. The Model
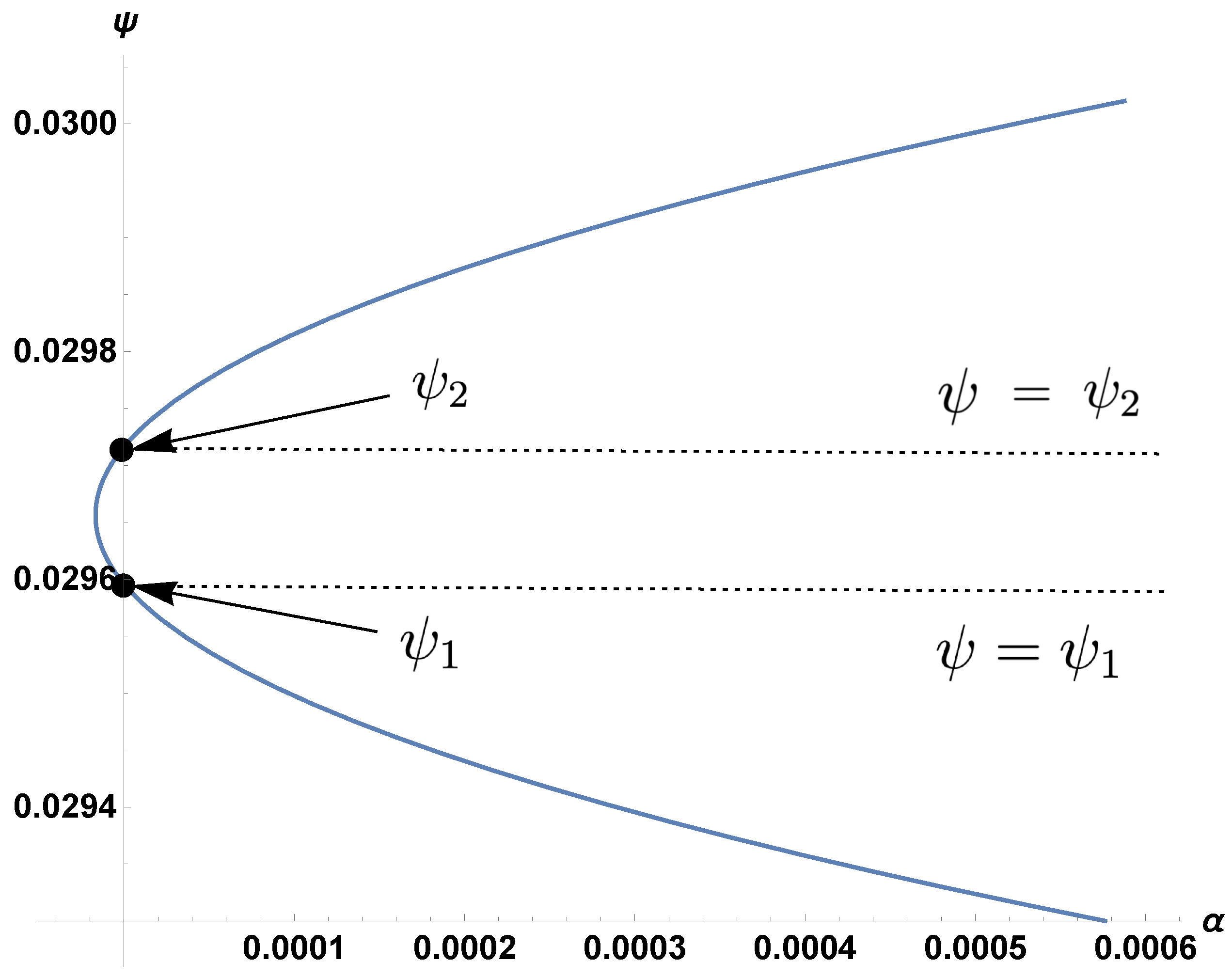
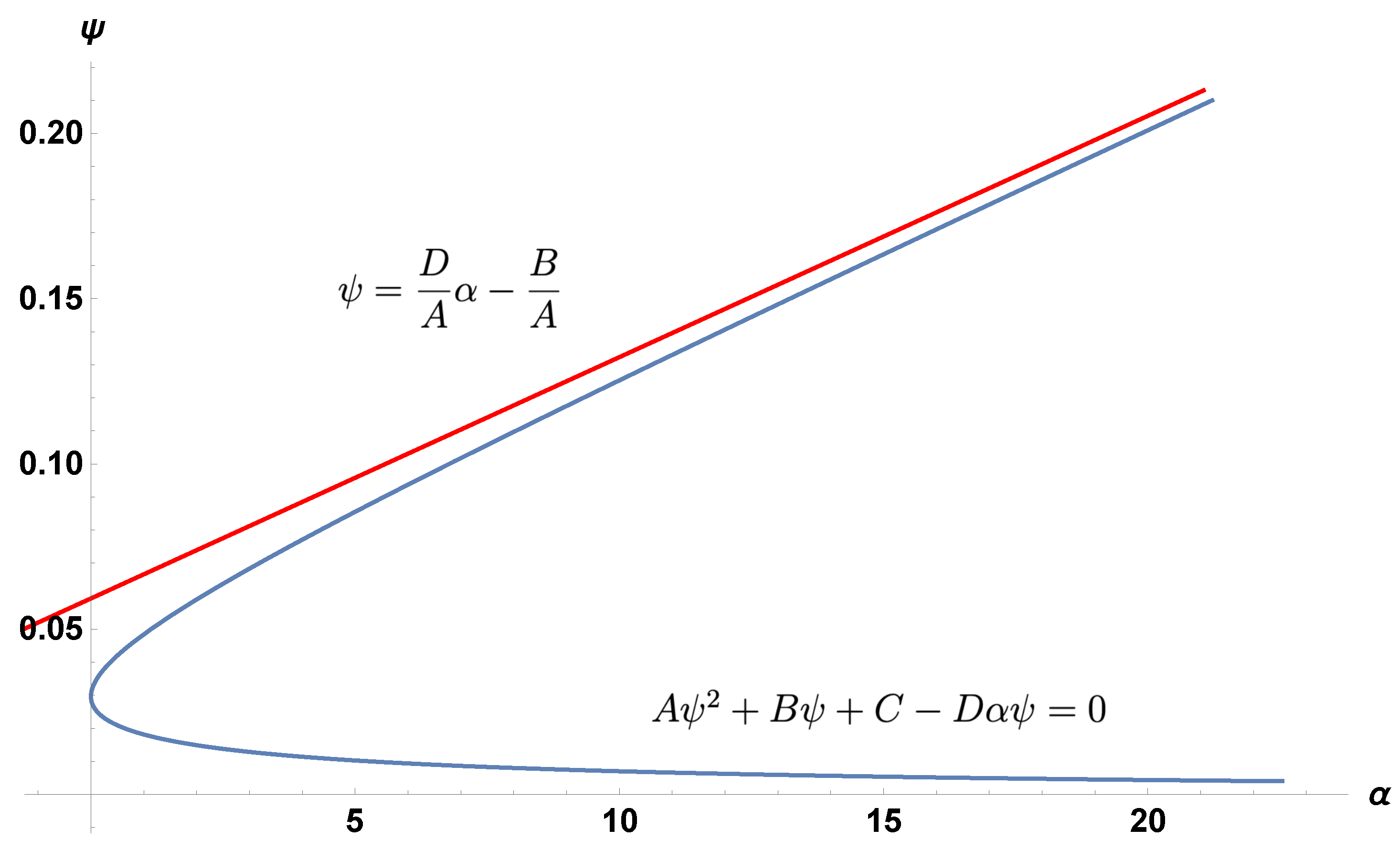
3. The Boundary and Coexistence Regions
4. Parameter Ranges and Sensitivity Analysis
4.1. Parameter Ranges
4.2. Sensitivity Analysis
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Derivation of Inequality (uid15)
References
- Albayrak, H.; Ozan, E. Serosurveillance for equine infectious anaemia in the Ardahan province of Turkey. Trop. Anim. Health Prod. 2010, 42, 1593–1595. [Google Scholar] [CrossRef]
- Issel, C.J.; Foil, L. Studies on equine infectious anemia virus transmission by insects. J. Am. Vet. Med. Assoc. 1984, 184, 293–297. [Google Scholar] [PubMed]
- Bolfa, P.; Barbuceanu, F.; Leau, S.E.; Leroux, C. Equine infectious anaemia in Europe: Time to re-examine the efficacy of monitoring and control protocols? Equine Vet. J. 2016, 48, 140–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroux, C.; Cadoré, J.L.; Montelaro, R.C. Equine Infectious Anemia Virus (EIAV): What has HIV’s country cousin got to tell us? Vet. Res. 2004, 35, 485–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craigo, J.K.; Ezzelarab, C.; Cook, S.J.; Chong, L.; Horohov, D.; Issel, C.J.; Montelaro, R.C. Envelope determinants of equine lentiviral vaccine protection. PLoS ONE 2013, 8, e66093. [Google Scholar] [CrossRef] [Green Version]
- Mealey, R.H.; Leib, S.R.; Littke, M.H.; Wagner, B.; Horohov, D.W.; McGuire, T.C. Viral load and clinical disease enhancement associated with a lentivirus cytotoxic T lymphocyte vaccine regimen. Vaccine 2009, 27, 2453–2468. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Blythe, D.C.; Loyd, H.; Mealey, R.H.; Tallmadge, R.L.; Dorman, K.S.; Carpenter, S. Decreased infectivity of a neutralization-resistant equine infectious anemia virus variant can be overcome by efficient cell-to-cell spread. J. Virol. 2011, 85, 10421–10424. [Google Scholar] [CrossRef] [Green Version]
- Issel, C.J.; Cook, R.F.; Mealey, R.H.; Horohov, D.W. Equine infectious anemia in 2014: Live with it or eradicate it? Vet. Clin. Equine Pract. 2014, 30, 561–577. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.N.; Rao, D.; Fu, X.Q.; Hu, M.M.; Dong, J.G. Equine infectious anemia virus in China. Oncotarget 2018, 9, 1356. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.D.; Leib, S.R.; Carpenter, S.; Mealey, R.H. Selection of a rare neutralization-resistant variant following passive transfer of convalescent immune plasma in equine infectious anemia virus-challenged SCID horses. J. Virol. 2010, 84, 6536–6548. [Google Scholar] [CrossRef] [Green Version]
- Taylor, S.D.; Leib, S.R.; Wu, W.; Nelson, R.; Carpenter, S.; Mealey, R.H. Protective effects of broadly neutralizing immunoglobulin against homologous and heterologous equine infectious anemia virus infection in horses with severe combined immunodeficiency. J. Virol. 2011, 85, 6814–6818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMichael, A.J.; Koff, W.C. Vaccines that stimulate T cell immunity to HIV-1: The next step. Nat. Immunol. 2014, 15, 319–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolard, S.N.; Kumaraguru, U. Viral vaccines and CTL response. J. Biomed. Biotechnol. 2010, 2010, 141657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammond, S.A.; Li, F.; McKeon, B.M., Sr.; Cook, S.J.; Issel, C.J.; Montelaro, R.C. Immune responses and viral replication in long-term inapparent carrier ponies inoculated with equine infectious anemia virus. J. Virol. 2000, 74, 5968–5981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Craigo, J.K.; Howe, L.; Steckbeck, J.D.; Cook, S.; Issel, C.; Montelaro, R.C. A live attenuated equine infectious anemia virus proviral vaccine with a modified S2 gene provides protection from detectable infection by intravenous virulent virus challenge of experimentally inoculated horses. J. Virol. 2003, 77, 7244–7253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craigo, J.K.; Li, F.; Steckbeck, J.D.; Durkin, S.; Howe, L.; Cook, S.J.; Issel, C.; Montelaro, R.C. Discerning an effective balance between equine infectious anemia virus attenuation and vaccine efficacy. J. Virol. 2005, 79, 2666–2677. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.; Leroux, C.; Issel, C. Equine infectious anemia and equine infectious anemia virus in 2013: A review. Vet. Microbiol. 2013, 167, 181–204. [Google Scholar] [CrossRef]
- Ciupe, S.M.; Schwartz, E.J. Understanding virus–host dynamics following EIAV infection in SCID horses. J. Theor. Biol. 2014, 343, 1–8. [Google Scholar] [CrossRef]
- Schwartz, E.J.; Smith?, R.J. Identifying the conditions under which antibodies protect against infection by equine infectious anemia virus. Vaccines 2014, 2, 397–421. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.J.; Nanda, S.; Mealey, R.H. Antibody escape kinetics of equine infectious anemia virus infection of horses. J. Virol. 2015, 89, 6945–6951. [Google Scholar] [CrossRef] [Green Version]
- Allen, L.J.; Schwartz, E.J. Free-virus and cell-to-cell transmission in models of equine infectious anemia virus infection. Math. Biosci. 2015, 270, 237–248. [Google Scholar] [CrossRef]
- Schwartz, E.J.; Vaidya, N.K.; Dorman, K.S.; Carpenter, S.; Mealey, R.H. Dynamics of lentiviral infection in vivo in the absence of adaptive immune responses. Virology 2018, 513, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Cangelosi, R.A.; Schwartz, E.J.; Wollkind, D.J. A quasi-steady-state approximation to the basic target-cell-limited viral dynamics model with a non-cytopathic effect. Front. Microbiol. 2018, 9, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geethamalini, S.; Balamuralitharan, S. Semianalytical solutions by homotopy analysis method for EIAV infection with stability analysis. Adv. Differ. Equ. 2018, 2018, 356. [Google Scholar] [CrossRef] [Green Version]
- Balamuralitharan, S.; Geethamalini, S. Parameter Estimation of Model for EIAV Infection using HPM. Int. J. Pure Appl. Math. 2017, 113, 196–204. [Google Scholar]
- Geethamalini, S.; Balamuralitharan, S. Dynamical analysis of EIAV infection with cytotoxic T-lymphocyte immune response delay. Results Appl. Math. 2019, 2, 100025. [Google Scholar] [CrossRef]
- Schwartz, E.J.; Costris-Vas, C.; Smith?, S.R. Modelling Mutation in Equine Infectious Anemia Virus Infection Suggests a Path to Viral Clearance with Repeated Vaccination. Viruses 2021, 13, 2450. [Google Scholar] [CrossRef]
- Schwartz, E.J.; Pawelek, K.A.; Harrington, K.; Cangelosi, R.; Madrid, S. Immune Control of Equine Infectious Anemia Virus Infection by Cell-Mediated and Humoral Responses. Appl. Math. 2013, 4, 171–177. [Google Scholar] [CrossRef]
- Meadows, T.; Schwartz, E.J. A model of virus infection with immune responses supports boosting CTL response to balance antibody response. In Computational and Mathematical Population Dynamics; Tuncer, N., Martcheva, M., Prosper, O., Childs, L., Eds.; World Scientific Publishing Europe Ltd.: London, UK, 2023; pp. 145–168. [Google Scholar]
- Riddle, D.F. Analytic Geometry; Cengage: Boston, MA, USA, 1995; p. 150. [Google Scholar]
- Kallrath, J. Complexity and Large Scale-Least Squares Problems. In Encyclopedia of Optimization, 2nd ed.; Floudas, C.A., Pardalos, P.M., Eds.; Springer: New York, NY, USA, 2008; pp. 435–447. [Google Scholar]
- Dong, H.; Song, B.; Dong, Z.; Wang, P. Multi-start Space Reduction (MSSR) surrogate-based global optimization method. Struct. Multidiscip. Optim. 2016, 54, 907–926. [Google Scholar] [CrossRef]
- Mealey, R.H.; Zhang, B.; Leib, S.R.; Littke, M.H.; McGuire, T.C. Epitope specificity is critical for high and moderate avidity cytotoxic T lymphocytes associated with control of viral load and clinical disease in horses with equine infectious anemia virus. Virology 2003, 313, 537–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mealey, R.H.; Sharif, A.; Ellis, S.A.; Littke, M.H.; Leib, S.R.; McGuire, T.C. Early detection of dominant Env-specific and subdominant Gag-specific CD8+ lymphocytes in equine infectious anemia virus-infected horses using major histocompatibility complex class I/peptide tetrameric complexes. Virology 2005, 339, 110–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craigo, J.K.; Montelaro, R.C. Lessons in AIDS vaccine development learned from studies of equine infectious, anemia virus infection and immunity. Viruses 2013, 5, 2963–2976. [Google Scholar] [CrossRef] [Green Version]
- Blower, S.M.; Dowlatabadi, H. Sensitivity and uncertainty analysis of complex models of disease transmission: An HIV model, as an example. Int. Stat. Rev. Int. Stat. 1994, 62, 229–243. [Google Scholar] [CrossRef]
- Marino, S.; Hogue, I.B.; Ray, C.J.; Kirschner, D.E. A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Theor. Biol. 2008, 254, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, E.J.; Blower, S.M. Predicting the Potential Individual- and Population- Level Effects of Imperfect Herpes Simplex Virus Type 2 Vaccines. J. Infect. Dis. 2005, 191, 1734–1746. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Mejia, G.; Hernandez-Vargas, E.A. Uncovering antibody cross-reaction dynamics in influenza A infections. Bioinformatics 2021, 37, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Sajadi, M.M.; Guan, Y.; DeVico, A.L.; Seaman, M.S.; Hossain, M.; Lewis, G.K.; Redfield, R.R. Correlation between circulating HIV-1 RNA and broad HIV-1 neutralizing antibody activity. J. Acquir. Immune Defic. Syndr. 2011, 57, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopman, G.; Mooij, P.; Dekking, L.; Mortier, D.; Nieuwenhuis, I.G.; van Heteren, M.; Kuipers, H.; Remarque, E.J.; Radošević, K.; Bogers, W.M. Correlation between virus replication and antibody responses in macaques following infection with pandemic influenza A virus. J. Virol. 2016, 90, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Tagmyer, T.L.; Craigo, J.K.; Cook, S.J.; Even, D.L.; Issel, C.J.; Montelaro, R.C. Envelope determinants of equine infectious anemia virus vaccine protection and the effects of sequence variation on immune recognition. J. Virol. 2008, 82, 4052–4063. [Google Scholar] [CrossRef] [Green Version]
- Funk, G.; Barbour, A.; Hengartner, H.; Kalinke, U. Mathematical model of a virus-neutralizing immunglobulin response. J. Theor. Biol. 1998, 195, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Tschetter, J.R.; Byrne, K.M.; Perryman, L.E.; MCGuire, T.C. Control of equine infectious anemia virus is not dependent on ADCC mediating antibodies. Virology 1997, 230, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Poropatich, K.; Sullivan, D.J., Jr. Human immunodeficiency virus type 1 long-term non-progressors: The viral, genetic and immunological basis for disease non-progression. J. Gen. Virol. 2011, 92, 247–268. [Google Scholar] [CrossRef]
- Collins, D.R.; Gaiha, G.D.; Walker, B.D. CD8+ T cells in HIV control, cure and prevention. Nat. Rev. Immunol. 2020, 20, 471–482. [Google Scholar] [CrossRef]
- Van Dorp, C.H.; van Boven, M.; de Boer, R.J. Immuno-epidemiological modeling of HIV-1 predicts high heritability of the set-point virus load, while selection for CTL escape dominates virulence evolution. PLoS Comput. Biol. 2014, 10, e1003899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cori, A.; Pickles, M.; van Sighem, A.; Gras, L.; Bezemer, D.; Reiss, P.; Fraser, C. CD4+ cell dynamics in untreated HIV-1 infection: Overall rates, and effects of age, viral load, sex and calendar time. AIDS 2015, 29, 2435–2446. [Google Scholar] [CrossRef] [PubMed]
- Glass, T.; Myer, L.; Lesosky, M. The role of HIV viral load in mathematical models of HIV transmission and treatment: A review. BMJ Glob. Health 2020, 5, e001800. [Google Scholar] [CrossRef]
- Hsu, D.C.; O’Connell, R.J. Progress in HIV vaccine development. Hum. Vaccines Immunother. 2017, 13, 1018–1030. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Definition | Units | Initial | Revised | |||
|---|---|---|---|---|---|---|
| Min | Max | Lower | Sample Value | Upper | ||
| Uninfected arrival () | 0 | 50 | 26.35 | 31 | 35.65 | |
| Uninfected death () | 0 | 0.05 | 0.036 | 0.042 | 0.048 | |
| Infectivity rate () | ||||||
| Infected cell death () | 0 | 0.05 | 0.015 | 0.018 | 0.021 | |
| Killing by CTLs (k) | 0.1 | 0.049 | 0.058 | 0.067 | ||
| Virus production (b) | 100 | 10000 | 2006 | 2360 | 2714 | |
| Virus clearance () | 0 | 20 | 7.5 | 8.82 | 10.14 | |
| Ab neutralization (f) | 0 | 30 | 13.03 | 15.33 | 17.63 | |
| CTL production () | 0 | 1 | 0 | 0.5 | 1 | |
| CTL death () | 0 | 200 | 44.19 | 51.98 | 59.78 | |
| Ab growth () | 0 | 150 | 0 | 75 | 150 | |
| Ab clearance () | 0 | 200 | 46.42 | 54.62 | 62.81 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hull-Nye, D.; Meadows, T.; Smith?, S.R.; Schwartz, E.J. Key Factors and Parameter Ranges for Immune Control of Equine Infectious Anemia Virus Infection. Viruses 2023, 15, 691. https://doi.org/10.3390/v15030691
Hull-Nye D, Meadows T, Smith? SR, Schwartz EJ. Key Factors and Parameter Ranges for Immune Control of Equine Infectious Anemia Virus Infection. Viruses. 2023; 15(3):691. https://doi.org/10.3390/v15030691
Chicago/Turabian StyleHull-Nye, Dylan, Tyler Meadows, Stacey R. Smith?, and Elissa J. Schwartz. 2023. "Key Factors and Parameter Ranges for Immune Control of Equine Infectious Anemia Virus Infection" Viruses 15, no. 3: 691. https://doi.org/10.3390/v15030691